Decrypting the Origin and Pathogenesis in Pregnant Ewes of a New Ovine Pestivirus Closely Related to Classical Swine Fever Virus

 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
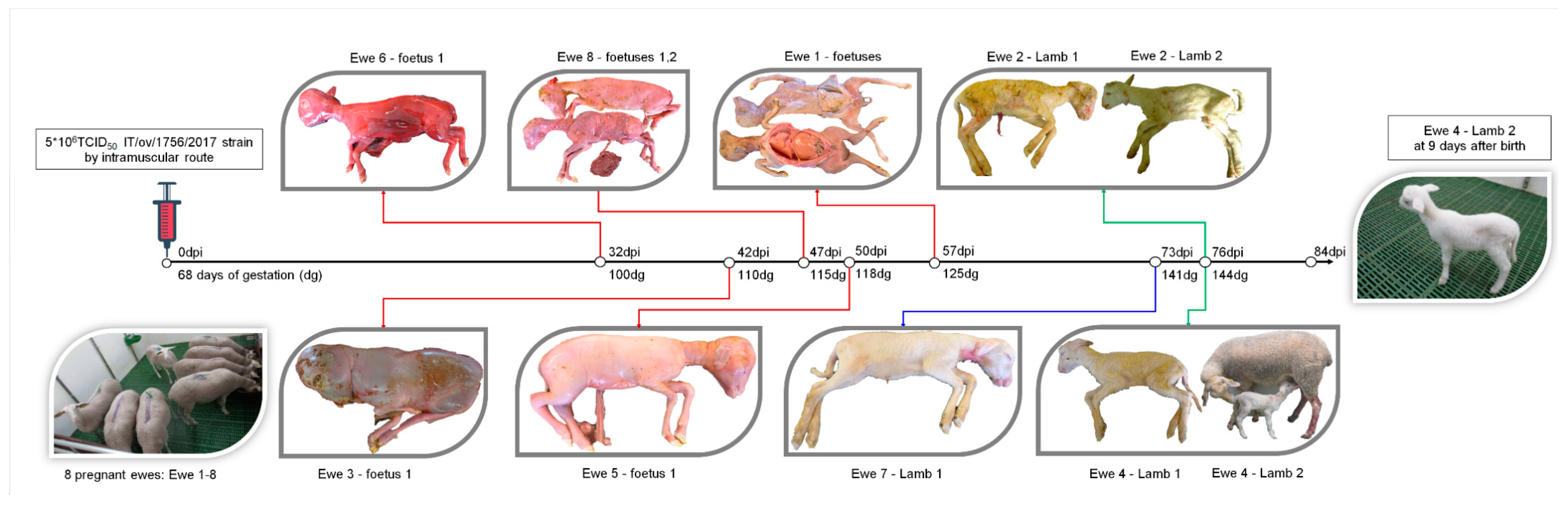
2.2. Experimental Infection in Pregnant Ewes
2.3. Pestivirus and CSFV-Specific RNA Detection
2.4. Design and Validation of a New RT-qPCR for the Specific Detection of OVPV RNA
2.5. Antibody Responses Characterization after OVPV Infection
2.6. Evolutionary Time Reconstruction and Cophylogenetic Analysis
3. Results
3.1. Specific and Sensitive Detection of OVPV RNA by the Newly-Designed Real Time TaqMan Assay
3.2. OVPV Causes a High Proportion of Abortions and Stillbirths in Pregnant Ewes after Infection
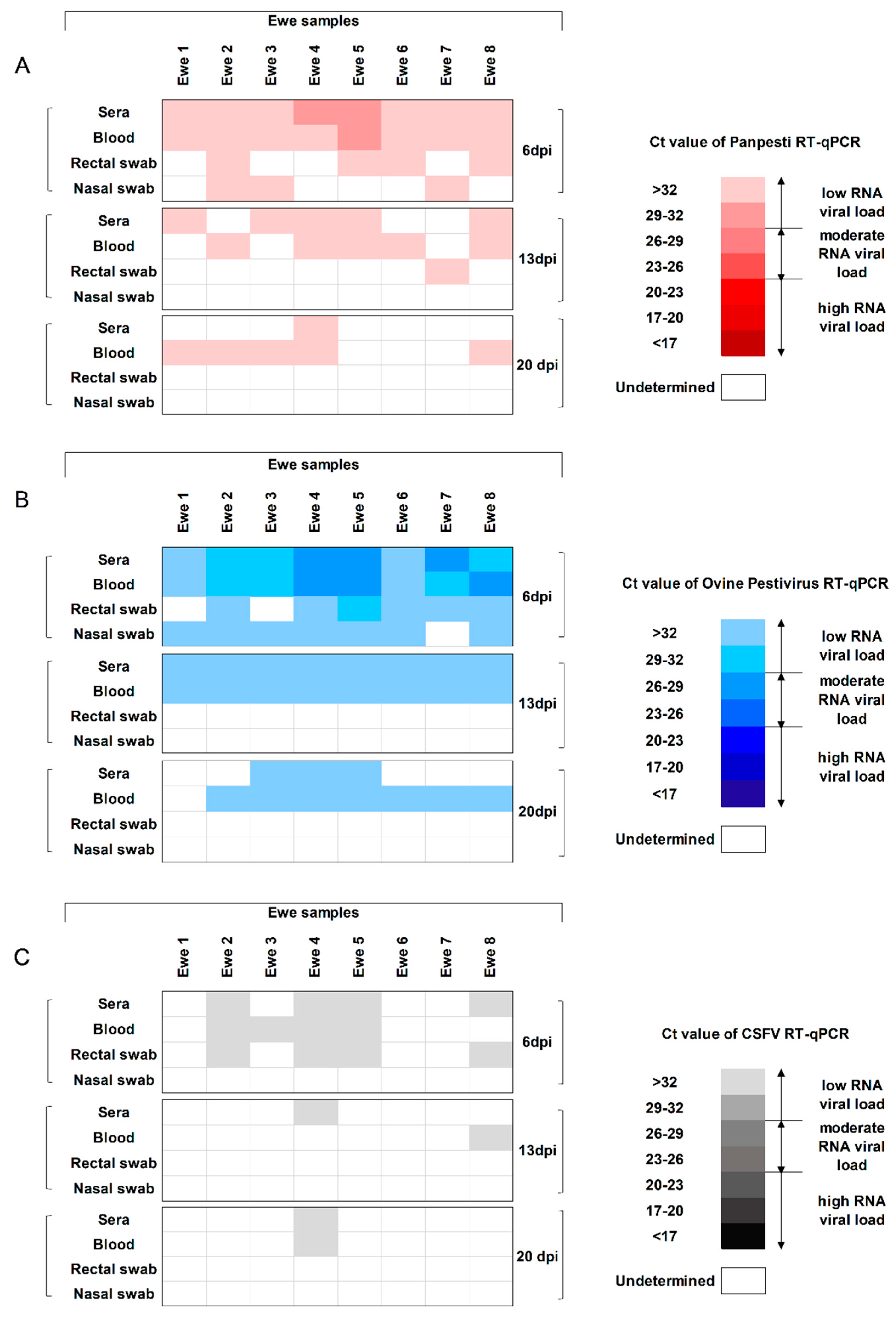
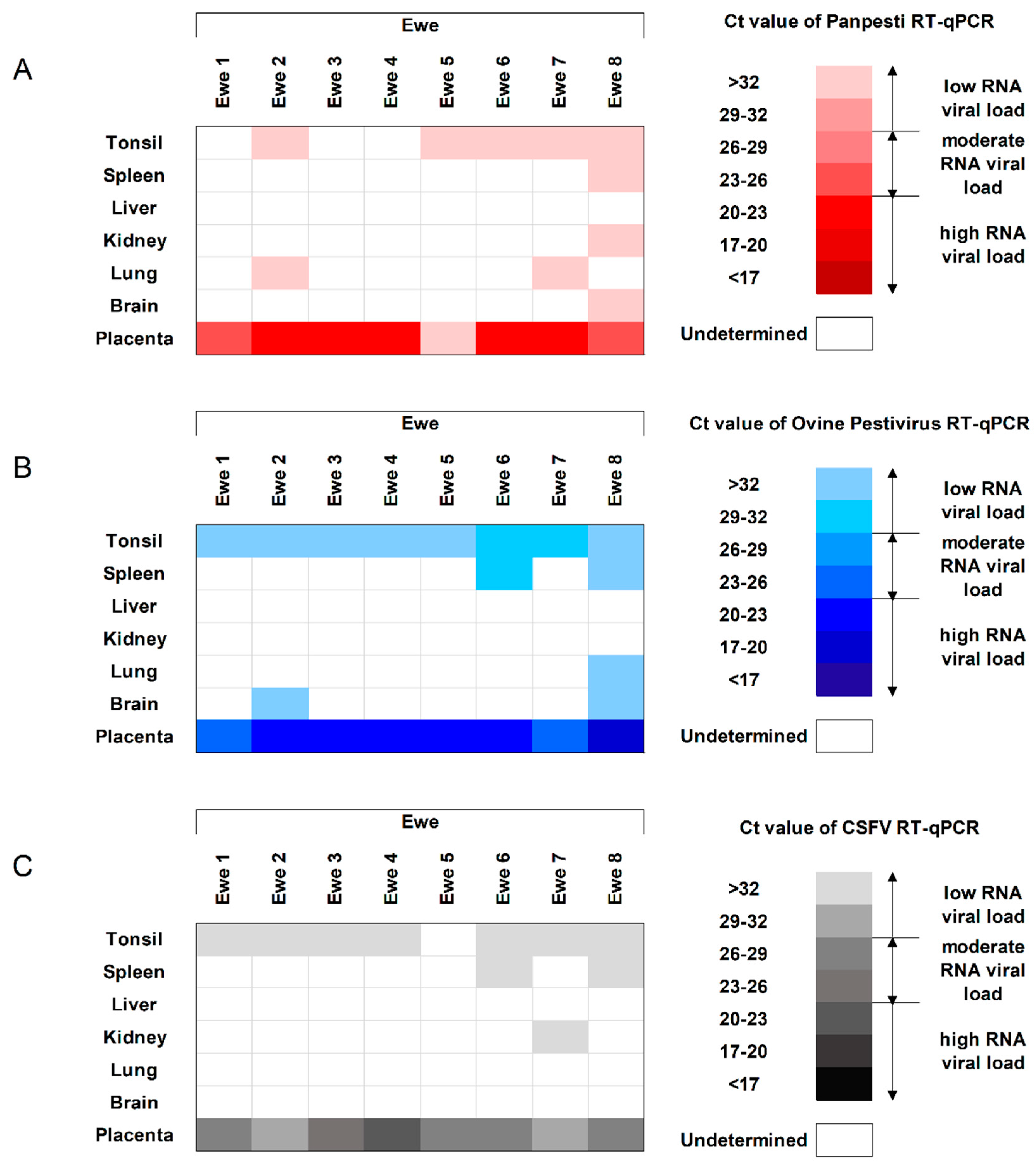
3.3. Viral Replication Capacity and Shedding of OVPV in the Infected Ewes
3.4. Efficient Vertical Transmission Capability and Generation of OVPV Congenitally-Persistently-Infected Lambs
3.5. The OVPV Viral Replication in Ewes, Fetuses, and Lambs Cross-Reacts with the Molecular Diagnosis of CSFV
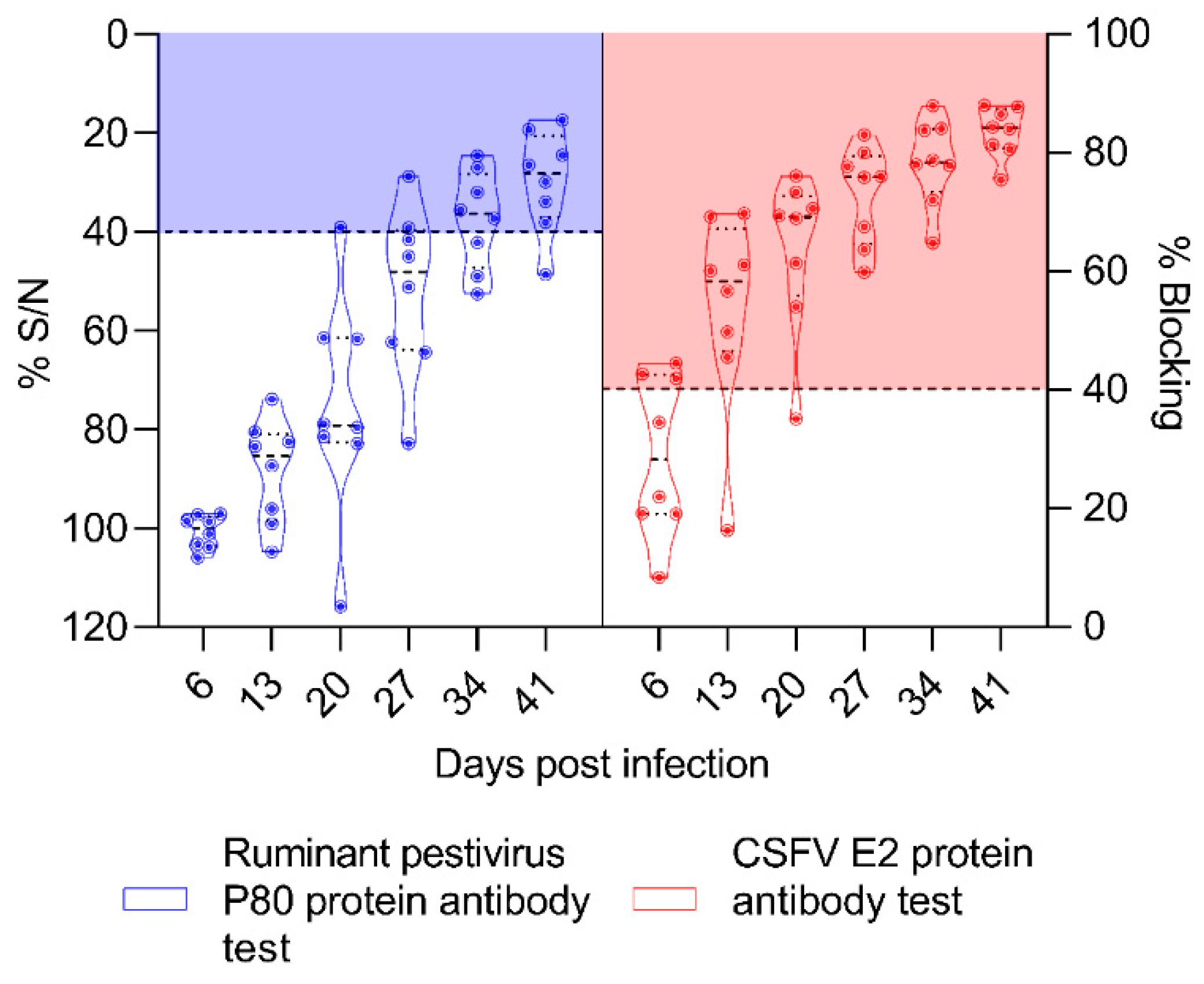
3.6. The OVPV Induced High and Early CSFV-Specific Antibody Response Detected by ELISA
3.7. OVPV Activates an Efficient Neutralizing Antibody Response that Cross-Reacts with CSFV and Other Pestiviruses
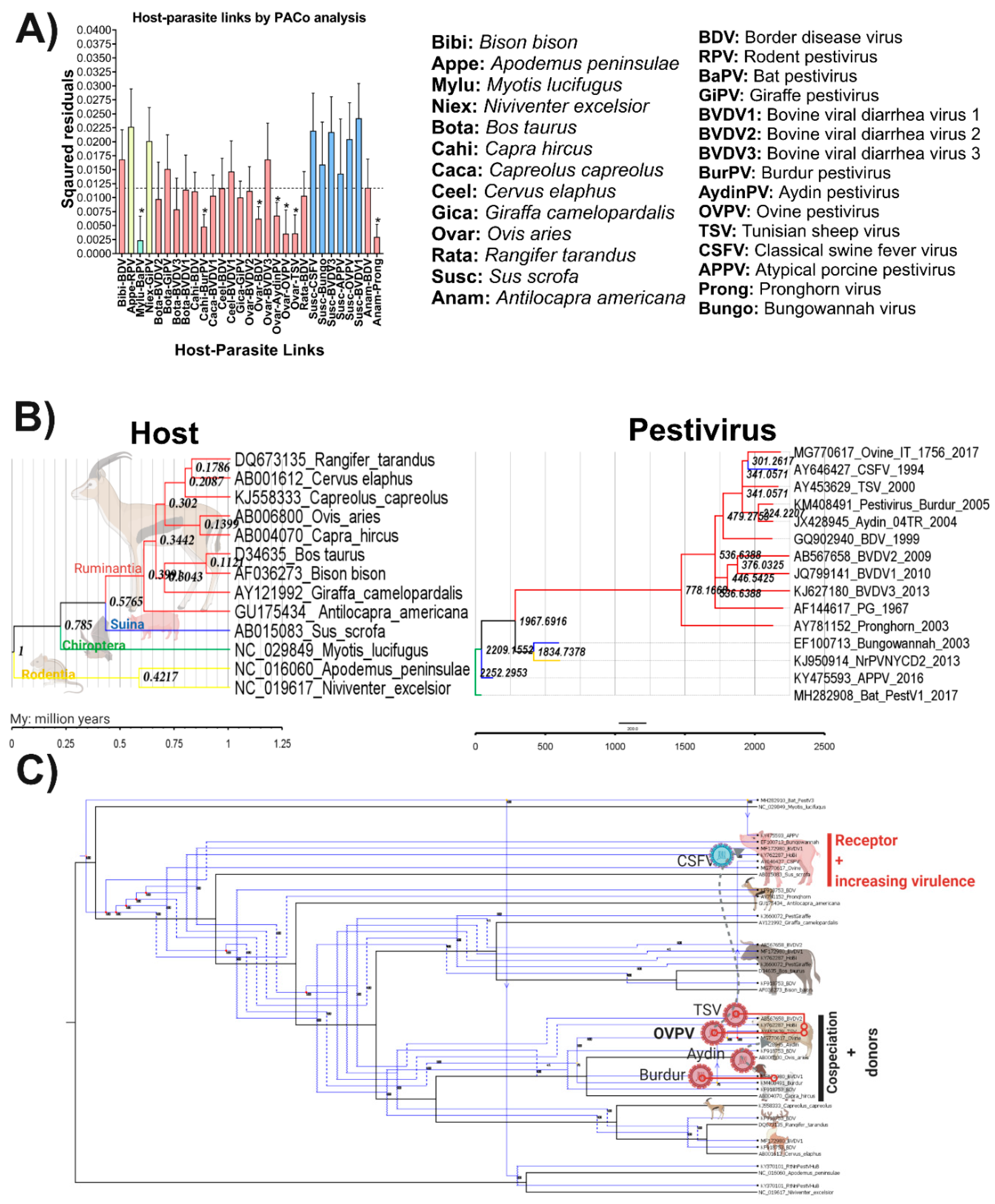
3.8. Evolutionary History and Cophylogenetic Reconstruction of OVPV and Their Vertebrate Hosts
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schweizer, M.; Peterhans, E. Pestiviruses. Annu. Rev. Anim. Biosci. 2014, 2, 141–163. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.B.; Meyers, G.; Bukh, J.; Gould, E.A.; Monath, T.; Muerhoff, A.S.; Pletnev, A.; Rico-Hesse, R.; Stapleton, J.T.; Simmonds, P.; et al. Proposed revision to the taxonomy of the genus Pestivirus, family Flaviviridae. J. Gen. Virol. 2017, 98, 2106–2112. [Google Scholar] [CrossRef] [PubMed]
- Hause, B.M.; Collin, E.A.; Peddireddi, L.; Yuan, F.; Chen, Z.; Hesse, R.A.; Gauger, P.C.; Clement, T.; Fang, Y.; Anderson, G. Discovery of a novel putative atypical porcine pestivirus in pigs in the USA. J. Gen. Virol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Arruda, B.L.; Arruda, P.H.; Magstadt, D.R.; Schwartz, K.J.; Dohlman, T.; Schleining, J.A.; Patterson, A.R.; Visek, C.A.; Victoria, J.G. Identification of a divergent lineage porcine pestivirus in nursing piglets with congenital tremors and reproduction of disease following experimental inoculation. PLoS ONE 2016, 11, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Groof, A.; Deijs, M.; Guelen, L.; Van Grinsven, L.; Van Os-Galdos, L.; Vogels, W.; Derks, C.; Cruijsen, T.; Geurts, V.; Vrijenhoek, M.; et al. Atypical porcine pestivirus: A possible cause of congenital tremor type A-II in newborn piglets. Viruses 2016, 8, 271. [Google Scholar] [CrossRef]
- Lamp, B.; Schwarz, L.; Högler, S.; Riedel, C.; Sinn, L.; Rebel-Bauder, B.; Weissenböck, H.; Ladinig, A.; Rümenapf, T. Novel pestiviruses in pigs. Emerg. Infect. Dis. 2017, 23, 2015–2018. [Google Scholar] [CrossRef]
- Tautz, N.; Tews, B.A.; Meyers, G. The Molecular Biology of Pestiviruses. In Advances in Virus Research; Academic Press Inc.: Cambridge, MA, USA, 2015; Volume 93, pp. 47–160. ISBN 9780128021798. [Google Scholar]
- Paton, D.J. Pestivirus diversity. J. Comp. Pathol. 1995, 112, 215–236. [Google Scholar] [CrossRef]
- Passler, T.; Walz, P.H. Bovine viral diarrhea virus infections in heterologous species. Anim. Health Res. Rev. 2010, 11, 191–205. [Google Scholar] [CrossRef]
- Braun, U.; Hilbe, M.; Peterhans, E.; Schweizer, M. Border disease in cattle. Vet. J. 2019, 246, 12–20. [Google Scholar] [CrossRef]
- Blome, S.; Staubach, C.; Henke, J.; Carlson, J.; Beer, M. Classical swine fever—an updated review. Viruses 2017, 9, 86. [Google Scholar] [CrossRef] [Green Version]
- Sozzi, E.; Lavazza, A.; Gaffuri, A.; Bencetti, F.C.; Prosperi, A.; Lelli, D.; Chiapponi, C.; Moreno, A. Isolation and full-length sequence analysis of a pestivirus from aborted lamb fetuses in Italy. Viruses 2019, 11, 744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Şevik, M. Genomic characterization of pestiviruses isolated from bovine, ovine and caprine foetuses in Turkey: A potentially new genotype of Pestivirus I species. Transbound. Emerg. Dis. 2020. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Wang, L.; Qiao, S.; Deng, R.; Zhang, G. Genetic characterization and recombination analysis of atypical porcine pestivirus. Infect. Genet. Evol. 2020, 81, 104259. [Google Scholar] [CrossRef] [PubMed]
- Wensvoort, G.; Terpstra, C.; Boonstra, J.; Bloemraad, M.; Van Zaane, D. Production of monoclonal antibodies against swine fever virus and their use in laboratory diagnosis. Vet. Microbiol. 1986, 12, 101–108. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938. [Google Scholar] [CrossRef]
- Rosell, R.; Cabezón, O.; Pujols, J.; Domingo, M.; Muñoz, I.; Núñez, J.I.; Ganges, L. Identification of a porcine pestivirus as a border disease virus from naturally infected pigs in Spain. Vet. Rec. 2014, 174, 18. [Google Scholar] [CrossRef]
- Tarradas, J.; de la Torre, M.E.; Rosell, R.; Perez, L.J.; Pujols, J.; Muñoz, M.; Muñoz, I.; Muñoz, S.; Abad, X.; Domingo, M.; et al. The impact of CSFV on the immune response to control infection. Virus Res. 2014, 185, 82–91. [Google Scholar] [CrossRef] [Green Version]
- García-Pérez, A.L.; Minguijón, E.; Estévez, L.; Barandika, J.F.; Aduriz, G.; Juste, R.A.; Hurtado, A. Clinical and laboratorial findings in pregnant ewes and their progeny infected with Border disease virus (BDV-4 genotype). Res. Vet. Sci. 2009, 86, 345–352. [Google Scholar] [CrossRef]
- Decaro, N.; Losurdo, M.; Larocca, V.; Lucente, M.S.; Mari, V.; Varello, K.; Patruno, G.; Camero, M.; Sciarra, M.; Occhiogrosso, L.; et al. HoBi-like pestivirus experimental infection in pregnant ewes: Reproductive disorders and generation of persistently infected lambs. Vet. Microbiol. 2015, 178, 173–180. [Google Scholar] [CrossRef]
- Hoffmann, B.; Depner, K.; Schirrmeier, H.; Beer, M. A universal heterologous internal control system for duplex real-time RT-PCR assays used in a detection system for pestiviruses. J. Virol. Methods 2006, 136, 200–209. [Google Scholar] [CrossRef]
- Hoffmann, B.; Beer, M.; Schelp, C.; Schirrmeier, H.; Depner, K. Validation of a real-time RT-PCR assay for sensitive and specific detection of classical swine fever. J. Virol. Methods 2005, 130, 36–44. [Google Scholar] [CrossRef]
- Tarradas, J.; Monsó, M.; Fraile, L.; de la Torre, B.G.; Muñoz, M.; Rosell, R.; Riquelme, C.; Pérez, L.J.; Nofrarías, M.; Domingo, M.; et al. A T-cell epitope on NS3 non-structural protein enhances the B and T cell responses elicited by dendrimeric constructions against CSFV in domestic pigs. Vet. Immunol. Immunopathol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-González, S.; Perez-Simó, M.; Muñoz, M.; Bohorquez, J.A.; Rosell, R.; Summerfield, A.; Domingo, M.; Ruggli, N.; Ganges, L. Efficacy of a live attenuated vaccine in classical swine fever virus postnatally persistently infected pigs. Vet. Res. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez, L.J.; Díaz de Arce, H.; Perera, C.L.; Rosell, R.; Frías, M.T.; Percedo, M.I.; Tarradas, J.; Dominguez, P.; Núñez, J.I.; Ganges, L. Positive selection pressure on the B/C domains of the E2-gene of classical swine fever virus in endemic areas under C-strain vaccination. Infect. Genet. Evol. 2012, 12, 1405–1412. [Google Scholar] [CrossRef] [PubMed]
- Terpstra, C.; Bloemraad, M.; Gielkens, A.L.J. The neutralizing peroxidase-linked assay for detection of antibody against swine fever virus. Vet. Microbiol. 1984. [Google Scholar] [CrossRef]
- Rios, L.; Coronado, L.; Naranjo-Feliciano, D.; Martínez-Pérez, O.; Perera, C.L.; Hernandez-Alvarez, L.; Díaz De Arce, H.; Núñez, J.I.; Ganges, L.; Pérez, L.J. Deciphering the emergence, genetic diversity and evolution of classical swine fever virus. Sci. Rep. 2017, 7, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree v112.2008. Available online: http://treebioedacuk/software/figtree (accessed on 17 July 2020).
- Balbuena, J.A.; Míguez-Lozano, R.; Blasco-Costa, I. PACo: A Novel Procrustes Application to Cophylogenetic Analysis. PLoS ONE 2013. [Google Scholar] [CrossRef] [Green Version]
- Conow, C.; Fielder, D.; Ovadia, Y.; Libeskind-Hadas, R. Jane: A new tool for the cophylogeny reconstruction problem. Algorithm. Mol. Biol. 2010. [Google Scholar] [CrossRef] [Green Version]
- Thornton, P.K. Livestock production: Recent trends, future prospects. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 2853–2867. [Google Scholar] [CrossRef] [Green Version]
- VanderWaal, K.; Deen, J. Global trends in infectious diseases of swine. Proc. Natl. Acad. Sci. USA 2018, 115, 11495–11500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sweasey, D.; Patterson, D.S.; Richardson, C.; Harkness, J.W.; Shaw, I.G.; Williams, W.W. Border disease: A sequential study of surviving lambs and an assessment of its effect on profitability. Vet. Rec. 1979. [Google Scholar] [CrossRef] [PubMed]
- Scherer, C.F.C.; Flores, E.F.; Weiblen, R.; Caron, L.; Irigoyen, L.F.; Neves, J.P.; Maciel, M.N. Experimental infection of pregnant ewes with bovine viral diarrhea virus type-2 (BVDV-2): Effects on the pregnancy and fetus. Vet. Microbiol. 2001, 79, 285–299. [Google Scholar] [CrossRef]
- Kuca, T.; Passler, T.; Newcomer, B.W.; Neill, J.D.; Galik, P.K.; Riddell, K.P.; Zhang, Y.; Walz, P.H. Identification of conserved amino acid substitutions during serial infection of pregnant cattle and sheep with Bovine Viral Diarrhea Virus. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Bohórquez, J.A.; Muñoz-González, S.; Pérez-Simó, M.; Muñoz, I.; Rosell, R.; Coronado, L.; Domingo, M.; Ganges, L. Foetal immune response activation and high replication rate during generation of classical swine fever congenital infection. Pathogens 2020, 9, 285. [Google Scholar] [CrossRef] [Green Version]
- Ridpath, J.F. Emerging pestiviruses infecting domestic and wildlife hosts. Anim. Health Res. 2015, 16, 55–59. [Google Scholar] [CrossRef]
- Bohórquez, J.A.; Wang, M.; Pérez-Simó, M.; Vidal, E.; Rosell, R.; Ganges, L. Low CD4/CD8 ratio in classical swine fever postnatal persistent infection generated at 3 weeks after birth. Transbound. Emerg. Dis. 2019, 66, 752–762. [Google Scholar] [CrossRef]
- Van Oirschot, J.T. Vaccinology of classical swine fever: From lab to field. Vet. Microbiol. 2003, 96, 367–384. [Google Scholar] [CrossRef]
- Ganges, L.; Núñez, J.I.; Sobrino, F.; Borrego, B.; Fernández-Borges, N.; Frías-Lepoureau, M.T.; Rodríguez, F. Recent advances in the development of recombinant vaccines against classical swine fever virus: Cellular responses also play a role in protection. Vet. J. 2008, 177, 169–177. [Google Scholar] [CrossRef]
- Kaden, V.; Lange, E.; Riebe, R.; Lange, B. Classical swine fever virus strain “C”. How long is it detectable after oral vaccination? J. Vet. Med. B Infect. Dis. Vet. Public Health 2004. [Google Scholar] [CrossRef]
- Walz, P.H.; Riddell, K.P.; Newcomer, B.W.; Neill, J.D.; Falkenberg, S.M.; Cortese, V.S.; Scruggs, D.W.; Short, T.H. Comparison of reproductive protection against bovine viral diarrhea virus provided by multivalent viral vaccines containing inactivated fractions of bovine viral diarrhea virus 1 and 2. Vaccine 2018, 36, 3853–3860. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-González, S.; Sordo, Y.; Pérez-Simó, M.; Suárez, M.; Canturri, A.; Rodriguez, M.P.; Frías-Lepoureau, M.T.; Domingo, M.; Estrada, M.P.; Ganges, L. Efficacy of E2 glycoprotein fused to porcine CD154 as a novel chimeric subunit vaccine to prevent classical swine fever virus vertical transmission in pregnant sows. Vet. Microbiol. 2017, 205, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Casciari, C.; Sozzi, E.; Bazzucchi, M.; Moreno Martin, A.M.; Gaffuri, A.; Giammarioli, M.; Lavazza, A.; De Mia, G.M. Serological relationship between a novel ovine pestivirus and classical swine fever virus. Transbound. Emerg. Dis. 2020, 1–5. [Google Scholar] [CrossRef] [PubMed]
- van Rijn, P.A.; van Gennip, R.G.P.; de Meijer, E.J.; Moormann, R.J.M. A preliminary map of epitopes on envelope glycoprotein E1 of HCV strain Brescia. Vet. Microbiol. 1992, 33, 221–230. [Google Scholar] [CrossRef]
- Geoghegan, J.L.; Duchêne, S.; Holmes, E.C. Comparative analysis estimates the relative frequencies of co-divergence and cross-species transmission within viral families. PLoS Pathog. 2017, 13, 1–17. [Google Scholar] [CrossRef]
- Geoghegan, J.L.; Senior, A.M.; Holmes, E.C. Pathogen population bottlenecks and adaptive landscapes: Overcoming the barriers to disease emergence. Proc. R. Soc. B 2016, 283. [Google Scholar] [CrossRef] [Green Version]
- Peters, R. Tunis, broad-tailed, mountain-sheep. In Memoris of the Philadelpia Society for Promoting Agriculture; Philadelphia Society for Promoting Agriculture: Philadephia, PA, USA, 1810; pp. 211–240. [Google Scholar]
- Brier, C.E. Tending our vines: From the correspondence and writings of Richard Peters and John Jay. Pa. Hist. 2013, 80, 85–111. [Google Scholar] [CrossRef]
- Mearns, R. Abortion in sheep. 1. Investigation and principal causes. In Practice 2007, 29, 40–46. [Google Scholar] [CrossRef]
- Kirkbride, C.A. Kirkbride’s Diagnosis of Abortion and Neonatal Loss in Animals, 4th ed.; Njaa, B.L., Ed.; John Wiley & Sons, Ltd.: Chichester, UK, 2012; ISBN 9780470958520. [Google Scholar]
- Hurtado, A.; García-Pérez, A.L.; Aduriz, G.; Juste, R.A. Genetic diversity of ruminant pestiviruses from Spain. Virus Res. 2003, 92, 67–73. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pestivirus Species and Subgenotype | Reference Strain /Isolate | Source |
|---|---|---|
| BDV | BDV 137/4 | Central Veterinary Laboratory (CVL), Weybridge, UK |
| BDV FRITJNERS | Institute of Animal Science and Health (ID-DLO), Lelystad Netherlands | |
| BDV MOREDUM | EU Reference Laboratory for CSF, Germany | |
| BVDV | BVDV NADL | EU Reference Laboratory for CSF, Germany |
| CSFV 1.1 | Alfort 187 | EU Reference Laboratory for CSF, Germany |
| HCLV vaccine (C-Strain) [24] | CReSA, Spain | |
| Thiverval | CReSA, Spain | |
| Koslov | EU Reference Laboratory for CSF, Germany | |
| Glentorf | EU Reference Laboratory for CSF, Germany | |
| CSFV 1.2 | Brescia | Institute of Animal Science and health (ID-DLO) Lelystad Netherlands |
| Baker | Central Veterinary Laboratory (CVL), Weybridge, UK | |
| CSFV 1.4 | Margarita | CReSA, Spain |
| Pinar del Rio [25] | CReSA, Spain | |
| CSFV 2.1 | Paderborn (CSF277 reference strain) | CReSA, Spain |
| Spain 97 | Laboratori Sanitat Ramadera (Barcelona) Spain | |
| CSFV 2.2 | CSF573 reference strain (Italy Parna’98) | CReSA, Spain |
| CSFV 2.3 | Diepholz (CSF104 reference strain) | EU Reference Laboratory for CSF, Germany |
| Catalonia 01 [18] | CReSA, Spain | |
| Spreda (CSF123 reference strain) | CReSA, Spain | |
| Rostock (CSF184 reference strain) | EU Reference Laboratory for CSF, Germany | |
| Uelzen (CSF639 reference strain) | CReSA, Spain | |
| APPV | Nasal swab from field-infected pig | CReSA, Spain |
| Thymus from field-infected pig | CReSA, Spain | |
| Novel ovine pestivirus (OVPV) | Inoculum Italian ovine pestivirus [12] | Istituto Zooprofilattico Sperimentale della Lombardia e Dell’Emilia Romagna-IZSLER |
| Animal Number | 6 dpi | 13 dpi | 20 dpi | 27 dpi | 34 dpi | 41 dpi | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Homologous Virus (Italy OVPV) | CSFV Alfort/ 187 | CSFV Margarita | CSFV Diepholz1 | CSFV Catalonia01 | BVDV NADL | BDV 137/4 | BDV-pig-SP-2007 | ||||||
| Ewe 1 | Neg. a | 1/160 | 1/640 | 1/320 | 1/5120 | 1/10240 | 1/80 | 1/40 | 1/1280 | 1/1280 | 1/40 | 1/80 | 1/320 |
| Ewe 2 | Neg. | 1/320 | 1/1280 | 1/1280 | 1/1280 | 1/5120 | 1/80 | 1/60 | 1/2560 | 1/1920 | 1/160 | 1/320 | 1/160 |
| Ewe 3 | Neg. | 1/80 | 1/160 | 1/1280 | 1/1280 | 1/5120 | 1/80 | 1/30 | 1/920 | 1/3840 | 1/80 | 1/20 | 1/320 |
| Ewe 4 | Neg. | 1/640 | 1/1280 | 1/1280 | 1/640 | 1/2560 | 1/20 | 1/20 | 1/15360 | 1/2560 | 1/20 | 1/40 | 1/160 |
| Ewe 5 | Neg. | 1/320 | 1/1280 | 1/1280 | 1/1280 | 1/1280 | 1/10 | 1/15 | 1/640 | 1/320 | 1/10 | 1/40 | 1/40 |
| Ewe 6 | Neg. | 1/320 | 1/2560 | 1/640 | 1/1280 | 1/5120 | 1/20 | 1/30 | 1/3840 | 1/960 | 1/40 | 1/160 | 1/160 |
| Ewe 7 | Neg. | 1/160 | 1/640 | 1/2560 | 1/5120 | 1/5120 | 1/40 | 1/160 | 1/30720 | 1/3840 | 1/80 | 1/640 | 1/160 |
| Ewe 8 | Neg. | 1/320 | 1/1280 | 1/2560 | 1/2560 | 1/1280 | 1/20 | 1/60 | 1/5120 | 1/960 | 1/80 | 1/40 | 1/160 |
| Mean value | / | 1/290 | 1/1140 | 1/1400 | 1/2320 | 1/4480 | 1/44 | 1/51 | 1/7555 | 1/1960 | 1/64 | 1/168 | 1/185 |
| SD | / | 171 | 713 | 801 | 1808 | 2903 | 31 | 46 | 10511 | 1340 | 48 | 215 | 93 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Sozzi, E.; Bohórquez, J.A.; Alberch, M.; Pujols, J.; Cantero, G.; Gaffuri, A.; Lelli, D.; Rosell, R.; Bensaid, A.; et al. Decrypting the Origin and Pathogenesis in Pregnant Ewes of a New Ovine Pestivirus Closely Related to Classical Swine Fever Virus. Viruses 2020, 12, 775. https://0-doi-org.brum.beds.ac.uk/10.3390/v12070775
Wang M, Sozzi E, Bohórquez JA, Alberch M, Pujols J, Cantero G, Gaffuri A, Lelli D, Rosell R, Bensaid A, et al. Decrypting the Origin and Pathogenesis in Pregnant Ewes of a New Ovine Pestivirus Closely Related to Classical Swine Fever Virus. Viruses. 2020; 12(7):775. https://0-doi-org.brum.beds.ac.uk/10.3390/v12070775
Chicago/Turabian StyleWang, Miaomiao, Enrica Sozzi, José Alejandro Bohórquez, Mònica Alberch, Joan Pujols, Guillermo Cantero, Alessandra Gaffuri, Davide Lelli, Rosa Rosell, Albert Bensaid, and et al. 2020. "Decrypting the Origin and Pathogenesis in Pregnant Ewes of a New Ovine Pestivirus Closely Related to Classical Swine Fever Virus" Viruses 12, no. 7: 775. https://0-doi-org.brum.beds.ac.uk/10.3390/v12070775