Indoleamine 2,3-Dioxygenase Is Involved in Interferon Gamma’s Anti-BKPyV Activity in Renal Cells

 , , ,
, , ,  and
and 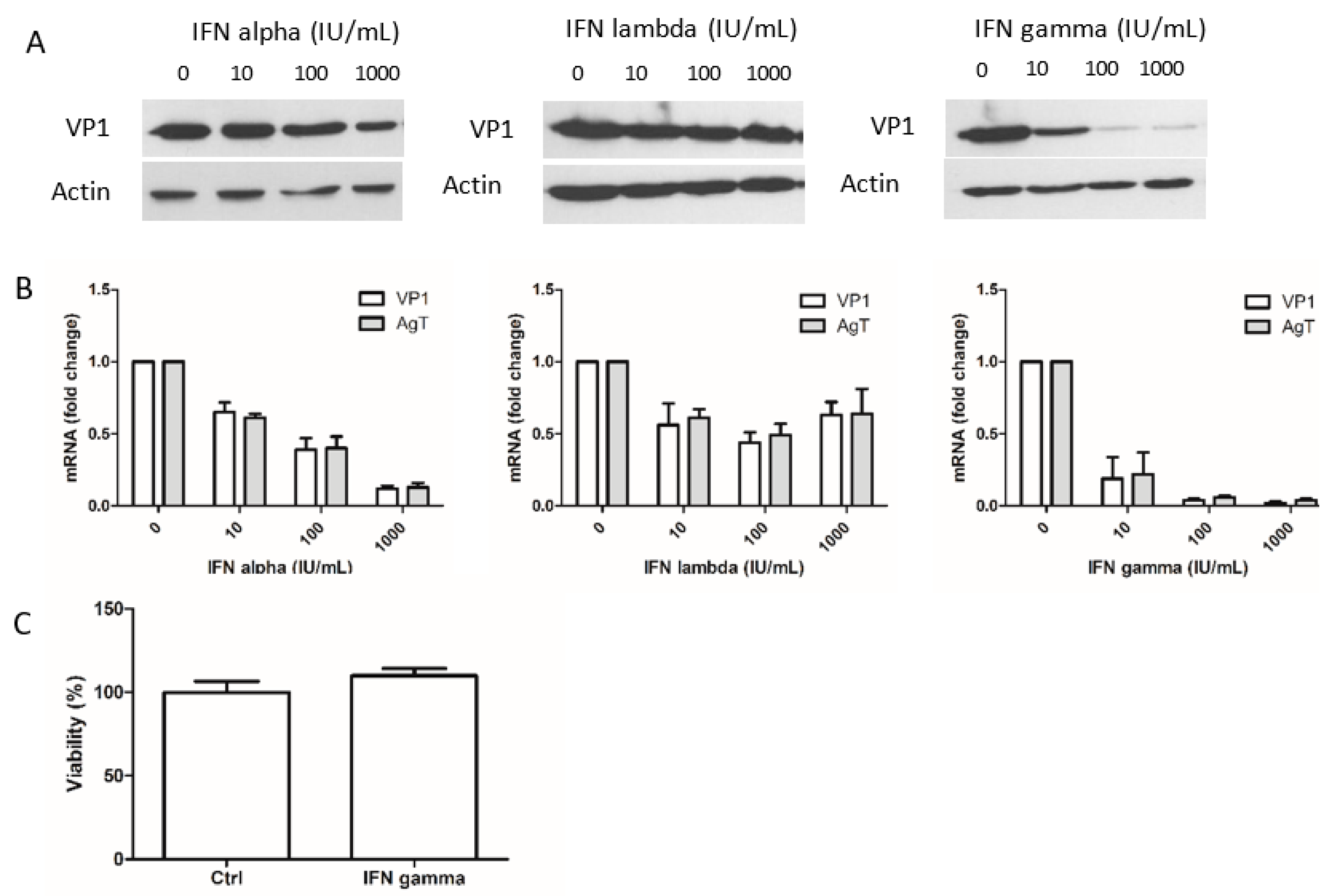{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures
2.2. Antibodies and Reagents
2.3. BKPyV Production
2.4. Cell Viability Assay
2.5. Immunofluorescence Staining
2.6. Western-Blot
2.7. Real-Time PCR
2.8. Kynurenine Assay
2.9. Statistical Analysis
3. Results
3.1. Interferon-Gamma Inhibits BKPyV Multiplication more Potently than IFN-Alpha and IFN-Lambda 1
3.2. The Jak-Stat Pathway Is Involved in the Antiviral Effect of IFNs on BKPyV Infection
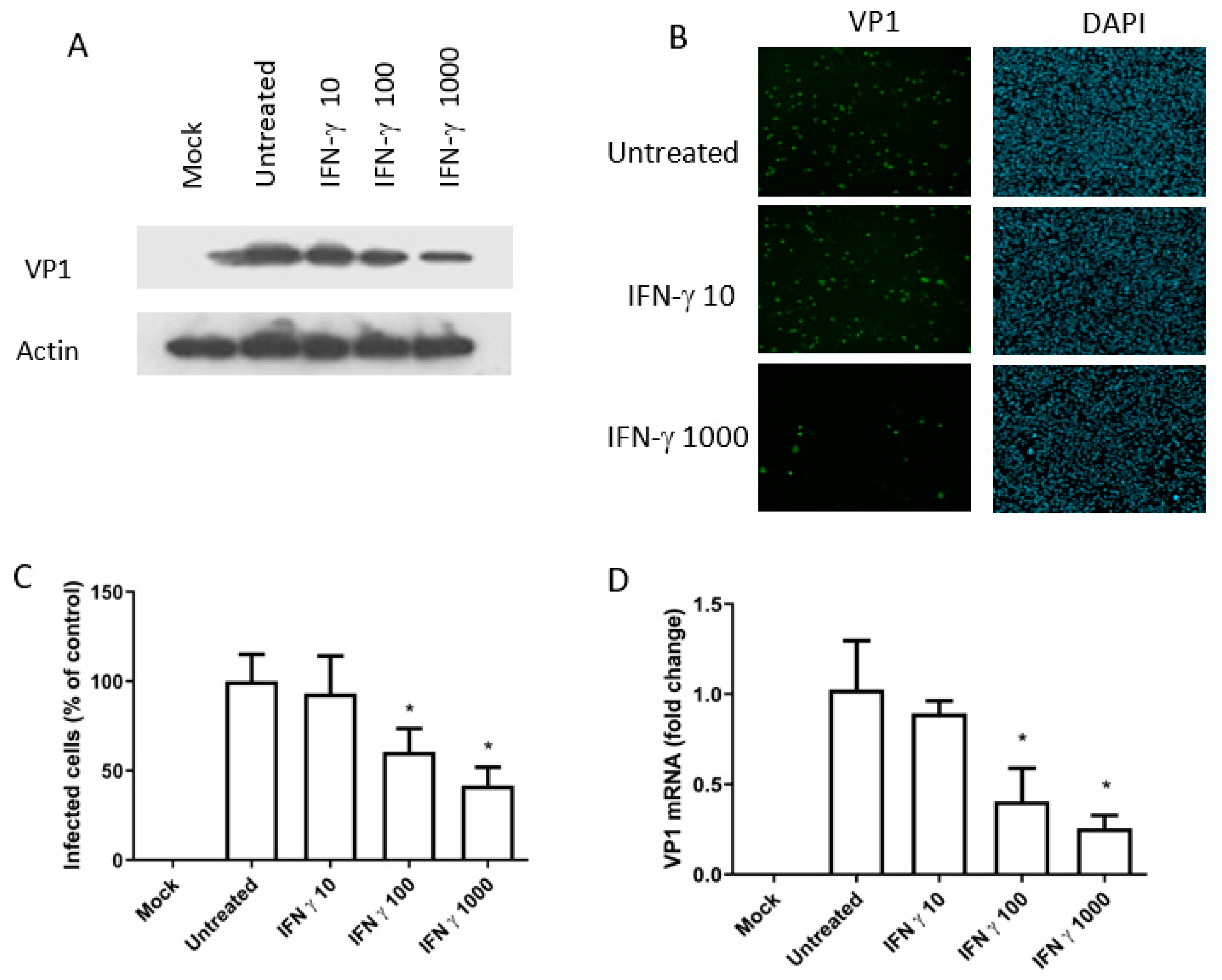
3.3. The Antiviral Activity of IFN-Gamma (BKPyV Infection of Caki-1 Cells)
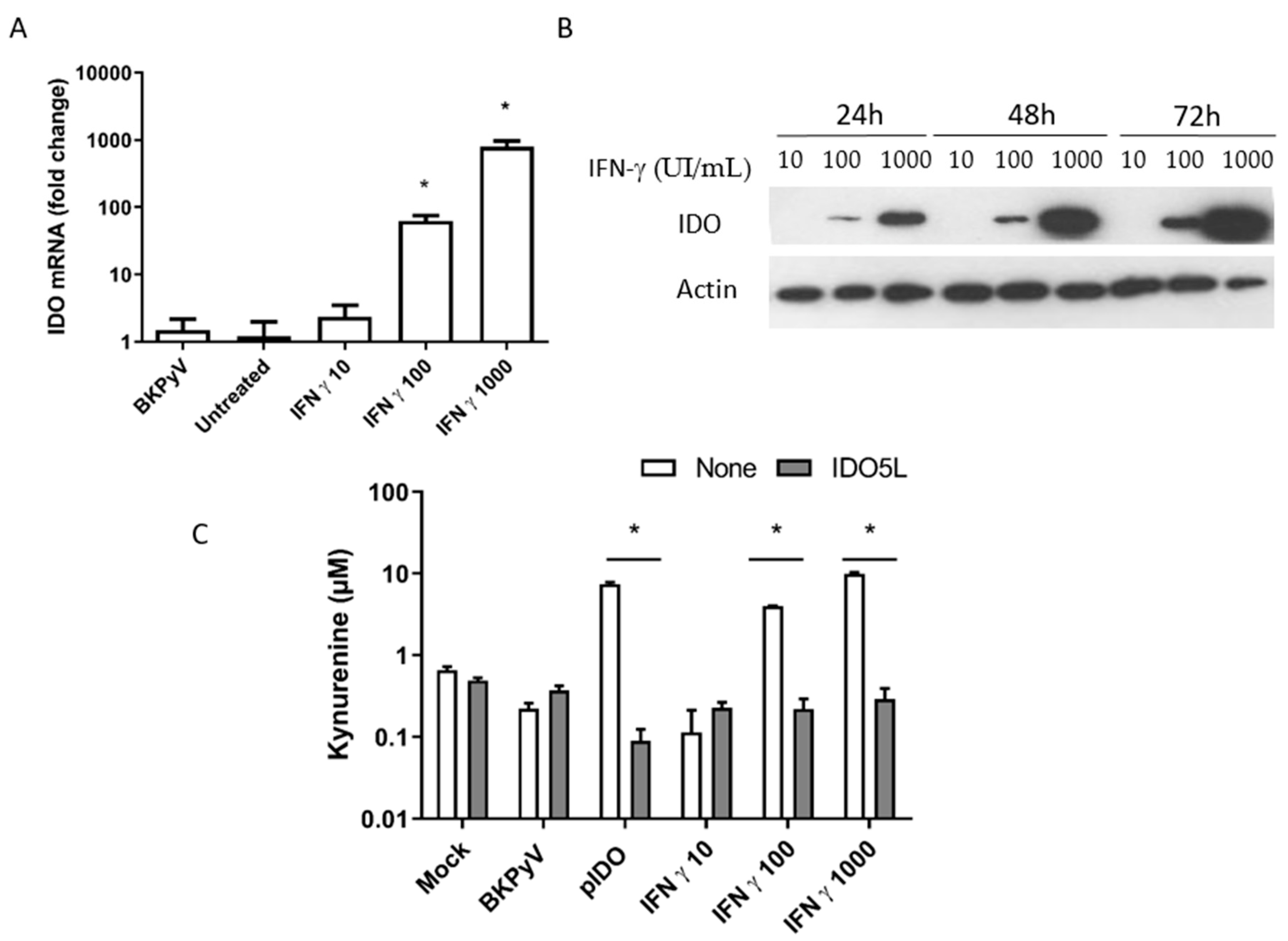
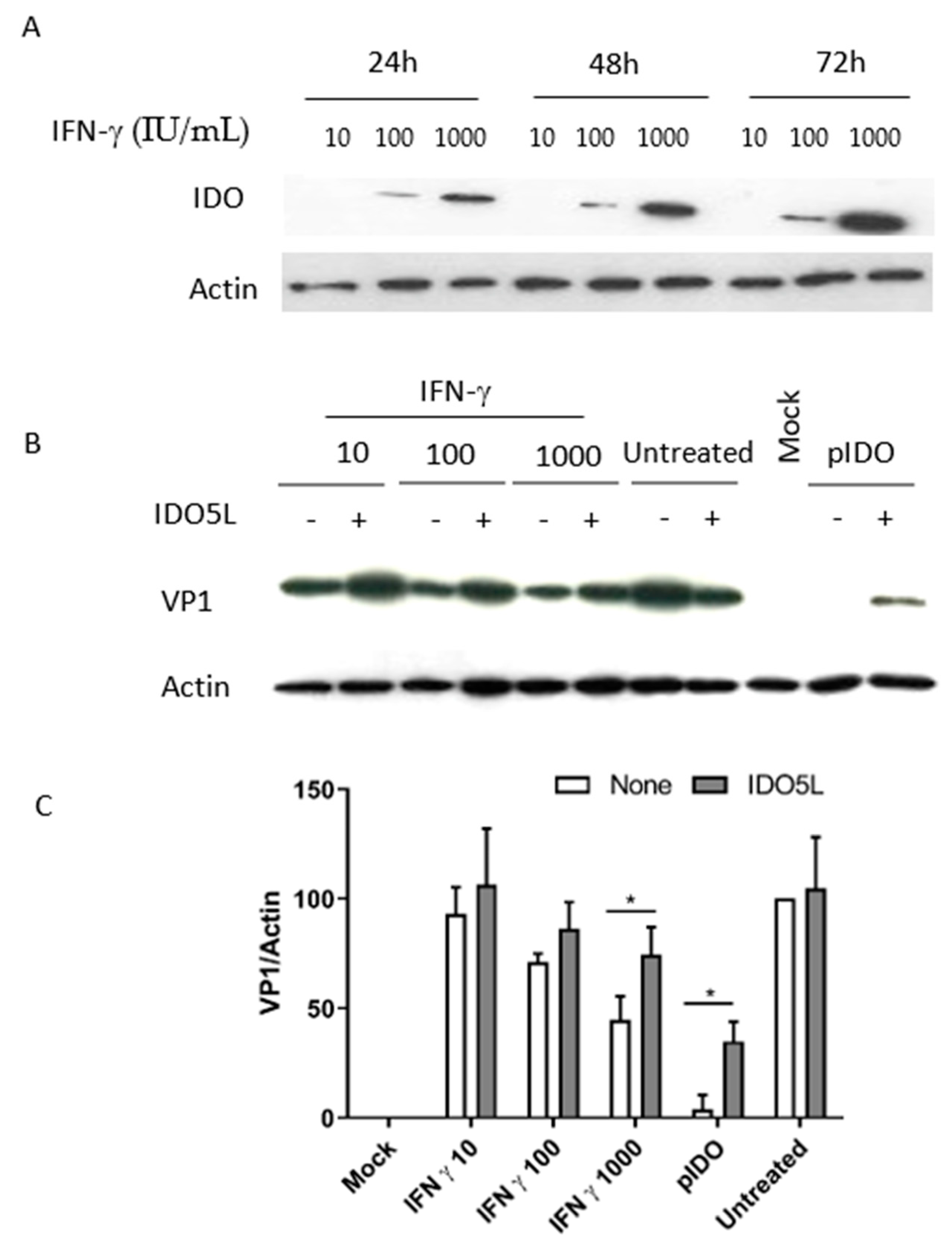
3.4. Expression of IDO in Caki-1 Cells
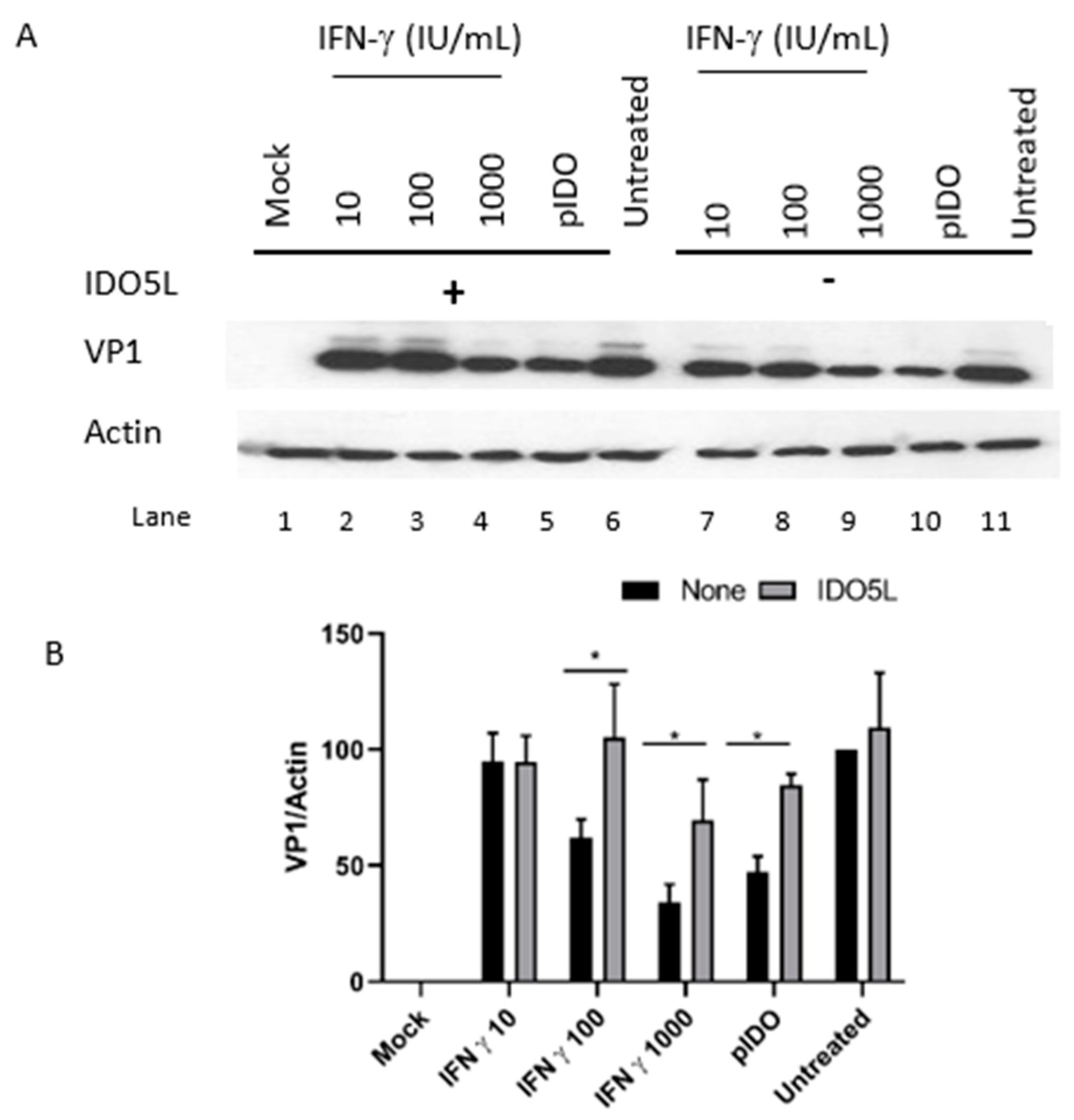
3.5. The Antiviral Effect of IDO (BKPyV Infection of Caki-1 Cells)
3.6. Antiviral Effect of IFN-Gamma and IDO in RPTE/TERT1 Cells
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Purighalla, R.; Shapiro, R.; McCauley, J.; Randhawa, P. BK virus infection in a kidney allograft diagnosed by needle biopsy. Am. J. Kidney Dis. 1995, 26, 671–673. [Google Scholar] [CrossRef]
- Binet, I.; Nickeleit, V.; Hirsch, H.H.; Prince, O.; Dalquen, P.; Gudat, F.; Mihatsch, M.J.; Thiel, G. Polyomavirus disease under new immunosuppressive drugs: A cause of renal graft dysfunction and graft loss. Transplantation 1999, 67, 918–922. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, H.H.; Randhawa, P.S. AST infectious diseases community of practice BK polyomavirus in solid organ transplantation—Guidelines from the American society of transplantation infectious diseases community of practice. Clin. Transplant. 2019, 33, e13528. [Google Scholar] [CrossRef] [PubMed]
- Randhawa, P.S.; Finkelstein, S.; Scantlebury, V.; Shapiro, R.; Vivas, C.; Jordan, M.; Picken, M.M.; Demetris, A.J. Human Polyoma virus-associated interstitial nephritis in the allograft kidney. Transplantation 1999, 67, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Isaacs, A.; Lindenmann, J. Virus interference. I. The interferon. J. Interferon Res. 1987, 7, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Abend, J.R.; Low, J.A.; Imperiale, M.J. Inhibitory effect of gamma interferon on BK virus gene expression and replication. J. Virol. 2007, 81, 272–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mellor, A.L.; Munn, D.H. Ido expression by dendritic cells: Tolerance and tryptophan catabolism. Nat. Rev. Immunol. 2004, 4, 762–774. [Google Scholar] [CrossRef]
- Bodaghi, B.; Goureau, O.; Zipeto, D.; Laurent, L.; Virelizier, J.L.; Michelson, S. Role of IFN-gamma-induced indoleamine 2,3 dioxygenase and inducible nitric oxide synthase in the replication of human cytomegalovirus in retinal pigment epithelial cells. J. Immunol. Baltim. Md 1950 1999, 162, 957–964. [Google Scholar]
- Adams, O.; Besken, K.; Oberdörfer, C.; MacKenzie, C.R.; Takikawa, O.; Däubener, W. Role of indoleamine-2,3-dioxygenase in alpha/beta and gamma interferon-mediated antiviral effects against herpes simplex virus infections. J. Virol. 2004, 78, 2632–2636. [Google Scholar] [CrossRef] [Green Version]
- Obojes, K.; Andres, O.; Kim, K.S.; Däubener, W.; Schneider-Schaulies, J. Indoleamine 2,3-dioxygenase mediates cell type-specific anti-measles virus activity of gamma interferon. J. Virol. 2005, 79, 7768–7776. [Google Scholar] [CrossRef] [Green Version]
- Terajima, M.; Leporati, A.M. Role of indoleamine 2,3-dioxygenase in antiviral activity of interferon-gamma against vaccinia virus. Viral Immunol. 2005, 18, 722–729. [Google Scholar] [CrossRef] [PubMed]
- Dugan, A.S.; Gasparovic, M.L.; Tsomaia, N.; Mierke, D.F.; O’Hara, B.A.; Manley, K.; Atwood, W.J. Identification of amino acid residues in BK virus VP1 that are critical for viability and growth. J. Virol. 2007, 81, 11798–11808. [Google Scholar] [CrossRef] [Green Version]
- Handala, L.; Blanchard, E.; Raynal, P.-I.; Roingeard, P.; Morel, V.; Descamps, V.; Castelain, S.; Francois, C.; Duverlie, G.; Brochot, E.; et al. BK polyomavirus hijacks extracellular vesicles for En Bloc transmission. J. Virol. 2020, 94, e01834-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Descamps, V.; Helle, F.; Louandre, C.; Martin, E.; Brochot, E.; Izquierdo, L.; Fournier, C.; Hoffmann, T.W.; Castelain, S.; Duverlie, G.; et al. The kinase-inhibitor sorafenib inhibits multiple steps of the hepatitis C virus infectious cycle in vitro. Antivir. Res. 2015, 118, 93–102. [Google Scholar] [CrossRef]
- Handala, L.; Fiore, T.; Rouill, Y. QuantIF: An Imagej macro to automatically determine the percentage of infected cells after immunofluorescence. Viruses 2019, 11, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubois, A.; François, C.; Descamps, V.; Fournier, C.; Wychowski, C.; Dubuisson, J.; Castelain, S.; Duverlie, G. Enhanced anti-HCV activity of interferon alpha 17 subtype. Virol. J. 2009, 6, 70. [Google Scholar] [CrossRef] [Green Version]
- Lepiller, Q.; Soulier, E.; Li, Q.; Lambotin, M.; Barths, J.; Fuchs, D.; Stoll-Keller, F.; Liang, T.J.; Barth, H. Antiviral and immunoregulatory effects of indoleamine-2,3-dioxygenase in hepatitis C virus infection. J. Innate Immun. 2015, 7, 530–544. [Google Scholar] [CrossRef]
- Yeung, A.W.S.; Wu, W.; Freewan, M.; Stocker, R.; King, N.J.C.; Thomas, S.R. Flavivirus infection induces indoleamine 2,3-dioxygenase in human monocyte-derived macrophages via tumor necrosis factor and NF-κB. J. Leukoc. Biol. 2012, 91, 657–666. [Google Scholar] [CrossRef]
- Mao, R.; Zhang, J.; Jiang, D.; Cai, D.; Levy, J.M.; Cuconati, A.; Block, T.M.; Guo, J.-T.; Guo, H. Indoleamine 2,3-dioxygenase mediates the antiviral effect of gamma interferon against hepatitis B virus in human hepatocyte-derived cells. J. Virol. 2011, 85, 1048–1057. [Google Scholar] [CrossRef] [Green Version]
- Adams, O.; Besken, K.; Oberdörfer, C.; MacKenzie, C.R.; Rüssing, D.; Däubener, W. Inhibition of human herpes simplex virus type 2 by interferon gamma and tumor necrosis factor alpha is mediated by indoleamine 2,3-dioxygenase. Microbes Infect. 2004, 6, 806–812. [Google Scholar] [CrossRef]
- Interferon Gamma Prevents Infectious Entry of Human Papillomavirus 16 via an L2-Dependent Mechanism. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pmc/articles/PMC5411602/ (accessed on 12 May 2020).
- Assetta, B.; De Cecco, M.; O’Hara, B.; Atwood, W.J. JC polyomavirus infection of primary human renal epithelial cells is controlled by a type I IFN-induced response. mBio 2016, 7, e00903-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaitin, D.A.; Roisman, L.C.; Jaks, E.; Gavutis, M.; Piehler, J. Inquiring into the differential action of interferons (IFNs): An IFN-α2 mutant with enhanced affinity to IFNAR1 is functionally similar to IFN-β. Mol. Cell. Biol. 2006, 26, 1888–1897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schachtner, T.; Müller, K.; Stein, M.; Diezemann, C.; Sefrin, A.; Babel, N.; Reinke, P. BK virus-specific immunity kinetics: A predictor of recovery from polyomavirus BK-associated nephropathy. Am. J. Transplant. 2011, 11, 2443–2452. [Google Scholar] [CrossRef] [PubMed]
- Schachtner, T.; Stein, M.; Babel, N.; Reinke, P. The loss of BKV-specific immunity from pretransplantation to posttransplantation identifies kidney transplant recipients at increased risk of BKV replication. Am. J. Transplant. 2015, 15, 2159–2169. [Google Scholar] [CrossRef] [PubMed]
- Bae, H.; Na, D.H.; Chang, J.-Y.; Park, K.H.; Min, J.W.; Ko, E.J.; Lee, H.; Yang, C.W.; Chung, B.H.; Oh, E.-J. Usefulness of BK virus-specific interferon-γ enzyme-linked immunospot assay for predicting the outcome of BK virus infection in kidney transplant recipients. Korean J. Intern. Med. 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, F.T.; Cao, J.S.; Zhao, J.; Yu, Y.; Qi, F.; Dai, X.C. IDO expressing dendritic cells suppress allograft rejection of small bowel transplantation in mice by expansion of Foxp3+ regulatory T cells. Transpl. Immunol. 2015, 33, 69–77. [Google Scholar] [CrossRef]
- Benavente, F.M.; Soto, J.A.; Pizarro-Ortega, M.S.; Bohmwald, K.; González, P.A.; Bueno, S.M.; Kalergis, A.M. Contribution of IDO to human respiratory syncytial virus infection. J. Leukoc. Biol. 2019, 106, 933–942. [Google Scholar] [CrossRef]
- Divanovic, S.; Sawtell, N.M.; Trompette, A.; Warning, J.I.; Dias, A.; Cooper, A.M.; Yap, G.S.; Arditi, M.; Shimada, K.; Duhadaway, J.B.; et al. Opposing biological functions of tryptophan catabolizing enzymes during intracellular infection. J. Infect. Dis. 2012, 205, 152–161. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fiore, T.; Martin, E.; Descamps, V.; Brochot, E.; Morel, V.; Handala, L.; Dakroub, F.; Castelain, S.; Duverlie, G.; Helle, F.; et al. Indoleamine 2,3-Dioxygenase Is Involved in Interferon Gamma’s Anti-BKPyV Activity in Renal Cells. Viruses 2020, 12, 865. https://0-doi-org.brum.beds.ac.uk/10.3390/v12080865
Fiore T, Martin E, Descamps V, Brochot E, Morel V, Handala L, Dakroub F, Castelain S, Duverlie G, Helle F, et al. Indoleamine 2,3-Dioxygenase Is Involved in Interferon Gamma’s Anti-BKPyV Activity in Renal Cells. Viruses. 2020; 12(8):865. https://0-doi-org.brum.beds.ac.uk/10.3390/v12080865
Chicago/Turabian StyleFiore, Tony, Elodie Martin, Véronique Descamps, Etienne Brochot, Virginie Morel, Lynda Handala, Fatima Dakroub, Sandrine Castelain, Gilles Duverlie, François Helle, and et al. 2020. "Indoleamine 2,3-Dioxygenase Is Involved in Interferon Gamma’s Anti-BKPyV Activity in Renal Cells" Viruses 12, no. 8: 865. https://0-doi-org.brum.beds.ac.uk/10.3390/v12080865