Phylogenetic Characterization of Arboviruses in Patients Suffering from Acute Fever in Rondônia, Brazil

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
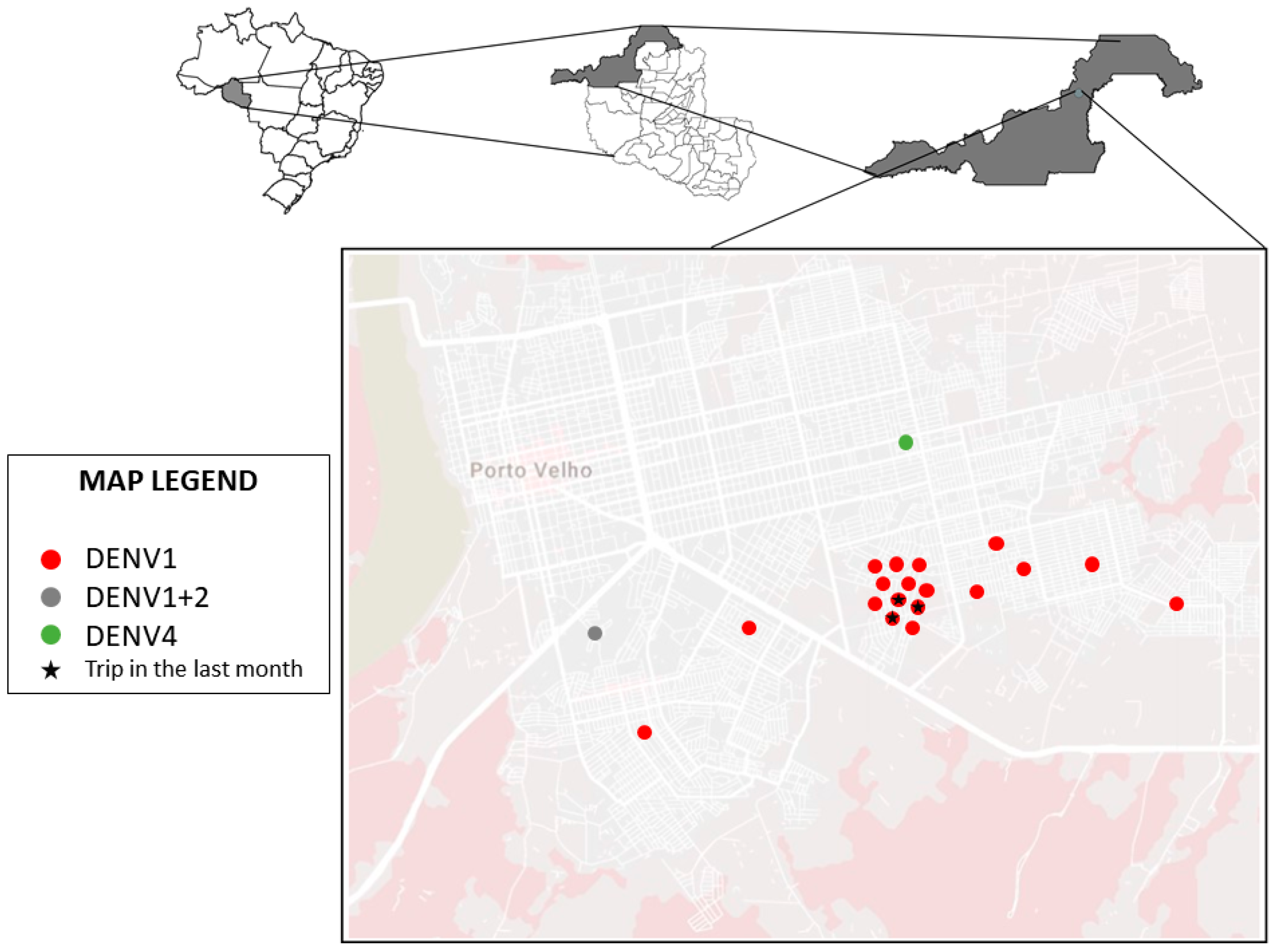
2.1. Location of Study
2.2. Samples
2.3. Ethics Declaration
2.4. Analyses
2.4.1. RT-qPCR for Zika, Dengue and Chikungunya
2.4.2. RT-qPCR for the Mayaro (MAYV) and Oropouche (OROV) Viruses
2.4.3. Reverse Transcription
2.4.4. PCR for DENV Sequencing
2.4.5. Evolutionary Analysis
2.4.6. Statistical Analysis
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Davis, L.E.; Beckham, J.D.; Tyler, K.L. North American encephalitic arboviruses. Neurol. Clin. 2008, 26, 727–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caputo, A.T.; Alonzi, D.S.; Kiappes, J.L.; Struwe, W.B.; Cross, A.; Basu, S.; Darlot, B.; Roversi, P. Dengue and Zika: Control and Antiviral Treatment Strategies; Springer: Singapore, 2018; Volume 1062, pp. 265–276. [Google Scholar] [CrossRef]
- Kuno, G.; Mackenzie, J.S.; Junglen, S.; Hubálek, Z.; Plyusnin, A.; Gubler, D.J. Vertebrate reservoirs of arboviruses: Myth, synonym of amplifier, or reality? Viruses 2017, 9, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weaver, S.C.; Reisen, W.K. Present and future arboviral threats. Antivir. Res. 2010, 85, 328–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima-Camara, T.N. Emerging arboviruses and public health challenges in Brazil. Rev. Saude Publica 2016, 50, S1518–S8787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Younger, D.S. Epidemiology of Zika Virus. Neurol. Clin. 2016, 34, 1049–1056. [Google Scholar] [CrossRef] [PubMed]
- Emmanuel, A.N.; Oliver, N.O.; Angela, U.N. Zika Virus infection: Damaging consequences in humans. Am. J. Life Sci. Res. 2016. [Google Scholar] [CrossRef]
- Kraemer, M.U.G.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.N.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. Albopictus. eLife 2015, 4, 1–18. [Google Scholar] [CrossRef]
- Long, K.C.; Ziegler, S.A.; Thangamani, S.; Hausser, N.L.; Kochel, T.J.; Higgs, S.; Tesh, R.B. Experimental transmission of Mayaro virus by Aedes aegypti. Am. J. Trop. Med. Hyg. 2011, 85, 750–757. [Google Scholar] [CrossRef] [Green Version]
- Kantor, A.M.; Lin, J.; Wang, A.; Thompson, D.C.; Franz, A.W.E. Infection Pattern of Mayaro virus in Aedes aegypti (Diptera: Culicidae) and transmission potential of the virus in mixed infections with Chikungunya virus. J. Med. Entomol. 2019, 56, 832–843. [Google Scholar] [CrossRef]
- Bonica, M.B.; Goenaga, S.; Martin, M.L.; Feroci, M.; Luppo, V.; Muttis, E.; Fabbri, C.; Morales, M.A.; Enria, D.; Micieli, M.V.; et al. Vector competence of Aedes aegypti for different strains of zika virus in Argentina. PLoS Negl. Trop. Dis. 2019, 13, 1–12. [Google Scholar] [CrossRef]
- Honório, N.A.; Wiggins, K.; Eastmond, B.; Câmara, D.C.P.; Alto, B.W. Experimental vertical transmission of chikungunya virus by brazilian and florida aedes albopictus populations. Viruses 2019, 11, 353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shroyer, D.A.; Rosen, L.; Lien, J.C.; Freier, J.E.; Tesh, R.B. Transovarial transmission of dengue viruses by mosquitoes: Aedes albopictus and Aedes aegypti. Am. J. Trop. Med. Hyg. 1983, 32, 1108–1119. [Google Scholar] [CrossRef]
- Figueiredo, L.T.M. The Brazilian flaviviruses. Microbes Infect. 2000, 2, 1643–1649. [Google Scholar] [CrossRef]
- Travassos Da Rosa, A.P.A.; Travassos Da Rosa, J.F.S.; Vasconcelos, P.F.C.; Pinheiro, F.P.; Rodrigues, S.G.; Travassos Da Rosa, E.S.; Dias, L.B.; Cruz, A.C.R. Arboviruses isolated in the Evandro Chagas Institute, including some described for the first time in the Brazilian Amazon region, their known hosts, and their pathology for man. In An Overview Arbovirology Brazil Neighbouring Ctries; Belém Instituto Evandro Chagas: Belém, Brazil, 1998; pp. 19–31. [Google Scholar]
- Ciccozzi, M.; Lai, A.; Zehender, G.; Borsetti, A.; Cella, E.; Ciotti, M.; Sagnelli, E.; Sagnelli, C.; Angeletti, S. The phylogenetic approach for viral infectious disease evolution and epidemiology: An updating review. J. Med. Virol. 2019, 91, 1707–1724. [Google Scholar] [CrossRef] [PubMed]
- Lemey, P.; Rambaut, A.; Drummond, A.J.; Suchard, M.A. Bayesian phylogeography finds its roots. PLoS Comput. Biol. 2009, 5, e1000520. [Google Scholar] [CrossRef] [Green Version]
- Giron, S.; Franke, F.; Decoppet, A.; Cadiou, B.; Travaglini, T.; Thirion, L.; Durand, G.; Jeannin, C.; L’Ambert, G.; Grard, G.; et al. Vector-borne transmission of Zika virus in Europe, southern France, August 2019. Eur. Surveill 2019, 24, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Durand, G.A.; Piorkowski, G.; Thirion, L.; Ninove, L.; Giron, S.; Zandotti, C.; Denis, J.; Badaut, C.; Failloux, A.B.; Grard, G.; et al. Vector-borne transmission of the Zika virus Asian genotype in Europe. Viruses 2020, 12, 296. [Google Scholar] [CrossRef] [Green Version]
- de Tanabe, E.L.L.; Tanabe, I.S.B.; dos Santos, E.C.; da Marques, J.P.S.; Borges, A.A.; de Lima, M.C.; Anderson, L.; Bassi, Ê.J. Report of east-central South African Chikungunya virus genotype during the 2016 outbreak in the Alagoas State, Brazil. Rev. Inst. Med. Trop. Sao Paulo 2018, 60. [Google Scholar] [CrossRef]
- Naveca, F.G.; Claro, I.; Giovanetti, M.; de Jesus, J.G.; Xavier, J.; de Iani, F.C.M.; do Nascimento, V.A.; de Souza, V.C.; Silveira, P.P.; Lourenço, J.; et al. Genomic, epidemiological and digital surveillance of Chikungunya virus in the Brazilian Amazon. PLoS Negl. Trop. Dis. 2019, 13, e0007065. [Google Scholar] [CrossRef] [Green Version]
- Tun, M.M.N.; Muthugala, R.; Kyaw, A.K.; Shimada, S.; Morita, K.; Hayasaka, D. Pathogenetic potential relating to metabolic activity in a mouse model of infection with the chikungunya virus East/Central/South African Genotype. Viruses 2020, 12, 169. [Google Scholar] [CrossRef] [Green Version]
- Goes De Jesus, J.; Da Luz Wallau, G.; Lima Maia, M.; Xavier, J.; Oliveira Lima, M.A.; Fonseca, V.; De Abreu, A.S.; De Oliveira Tosta, S.F.; Do Amaral, H.R.; Lima, I.A.B.; et al. Persistence of chikungunya ECSA genotype and local outbreak in an upper medium class neighborhood in Northeast Brazil. PLoS ONE 2020, 15, e0226098. [Google Scholar] [CrossRef]
- Ahmad, F.U.; Paul, S.K.; Aung, M.S.; Mazid, R.; Alam, M.; Ahmed, S.; Haque, N.; Hossain, M.A.; Paul, S.; Sharmin, R.; et al. Co-circulation of dengue virus type 3-genotype I and type 2-Cosmopolitan genotype in 2018 outbreak in Dhaka, Bangladesh. New Microbes New Infect. 2020, 33, 100629. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.C.V. Vírus Emergentes: A Infecção Pelo Vírus Do Dengue; Universidade de Lisboa: Lisboa, Portugal, 2014. [Google Scholar]
- de Bastos, M.S.; de Figueiredo, R.M.P.; Ramasawmy, R.; Itapirema, E.; Gimaque, J.B.L.; Santos, L.O.; Figueiredo, L.T.M.; Mourão, M.P.G. Simultaneous circulation of all four dengue serotypes in Manaus, State of Amazonas, Brazil in 2011. Rev. Soc. Bras. Med. Trop. 2012, 45, 393–394. [Google Scholar] [CrossRef]
- Rodrigues, N.; Maria, R.; Nogueira, R.; Maria, A.; de Filippis, B.; Nogueira, F.D.B.; Queiroz, R.; Barreto, F. Twenty years of DENV-2 activity in Brazil: Molecular characterization and phylogeny of strains isolated from 1990 to 2010. PLoS Negl. Trop. Dis. 2013, 7, e2095. [Google Scholar] [CrossRef] [Green Version]
- Yung, C.F.; Lee, K.S.; Thein, T.L.; Tan, L.K.; Gan, V.C.; Wong, J.G.X.; Lye, D.C.; Ng, L.C.; Leo, Y.S. Dengue serotype-specific differences in clinical manifestation, laboratory parameters and risk of severe disease in adults, Singapore. Am. J. Trop. Med. Hyg. 2015, 92, 999–1005. [Google Scholar] [CrossRef] [PubMed]
- Amâncio, F.F.; Ferraz, M.L.; de Almeida, M.C.M.; Pessanha, J.E.M.; Iani, F.C.M.; Fraga, G.L.; Lambertucci, J.R.; Carneiro, M. Dengue virus serotype 4 in a highly susceptible population in Southeast Brazil. J. Infect. Public Health 2014, 7, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Bertolacci-Rocha, L.G.; da Cunha, R.V.; de Lichs, G.G.C.; Fabbro, M.M.F.J.D.; Motta-Castro, A.R.C. Introduction of the dengue virus type 4 in the State of Mato Grosso do Sul, Brazil. Cad. Saude Publica 2014, 30, 1789–1792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira, D.S.; Bifano, G.D.S.; Honda, E.R.; Tada, M.S.; Batista, W.C. Isolation and identification of Dengue virus serotype 3 in Rondonia, Brazil. VIRUS Rev. Res. 2008, 13, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Batista, W.C.; Bifano, S.; Daniele, P.; Travagini, C.; Mendes, M.M.D.O.M.; Tada, S.; Stabeli, R.; Hildebrando, L.; Pesquisas, D.; Tropicais, P.; et al. Análise Epidemiológica Molecular do Vírus Dengue no Estado de Rondônia. Available online: https://rededengue.fiocruz.br/images/rededengue/Analise_epidemiologica_molecular_Rondonia.pdf (accessed on 1 June 2020).
- Vieira, D.S.; Honda, E.R.; Pereira, S.S.; Bifano, G.D.S.; Tada, M.S.; Batista, W.C. Characterization of dengue virus serotype 1 in epidemics in Porto Velho, Rondônia, in 2001–2003. Rev. Soc. Bras. Med. Trop. 2007, 40, 268–271. [Google Scholar] [CrossRef] [Green Version]
- Batista, W.C.; Bifano, S.; Vieira, D.S.; Honda, E.R. Case report/relato de caso notification of the first isolation of Cacipacore virus in a human in the State of Rondônia, Brazil Notificação do primeiro isolamento do vírus Cacipacoré em um ser humano, no Estado de. J. Med. Virol. 2011, 44, 528–530. [Google Scholar]
- de Lucena, L.T.; Aguiar, L.O.; Bogoevich, A.C.A.; de Azevedo, F.S.; dos Santos, A.C.P.; do Vale, D.B.A.P.; Pereira, D.B.; Villalobos-Salcedo, J.M. Dengue na Amazônia: Aspectos epidemiológicos no Estado de Rondônia, Brasil, de 1999 a 2010. Rev. Pan-Amazônica Saúde 2011, 2, 19–25. [Google Scholar] [CrossRef]
- Rios, F.G.F.; Nascimento, V.A.; Dal’acqua, D.S.V.; Naveca, F.G.; Julião, G.R. Vertical/venereal transmission of Dengue and Zika virus on field mosquitoes in the state of Rondônia, Brazilian Amazon. Mem. Inst. Oswaldo Cruz 2020. (Submitted). [Google Scholar]
- Naveca, F.G.; do Nascimento, V.A.; de Souza, V.C.; Nunes, B.T.D.; Rodrigues, D.S.G.; da Costa Vasconcelos, P.F. Multiplexed reverse transcription real-time polymerase chain reaction for simultaneous detection of Mayaro, Oropouche, and oropouche-like viruses. Mem. Inst. Oswaldo Cruz 2017, 112, 510–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanciotti, R.S.; Calisher, C.H.; Gubler, D.J.; Chang, G.J.; Vorndam, A.V. Rapid detection and typing of dengue viruses from clinical samples by using reverse transcriptase-polymerase chain reaction. J. Clin. Microbiol. 1992, 30, 545–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Prosdocimi, F. Racionalizando a Utilização do Algoritmo PHRED Para a Análise de Seqüências de DNA; Universidade Federal de Minas Gerais: Belo Horizonte, Brazil, 2006. [Google Scholar]
- Shrivastava, S.; Tiraki, D.; Diwan, A.; Lalwani, S.K.; Modak, M.; Mishra, A.C.; Arankalle, V.A. Co-circulation of all the four dengue virus serotypes and detection of a novel clade of DENV-4 (genotype I) virus in Pune, India during 2016 season. PLoS ONE 2018, 13, e0192672. [Google Scholar] [CrossRef] [Green Version]
- Touret, F.; Baronti, C.; Goethals, O.; Van Loock, M.; de Lamballerie, X.; Querat, G. Phylogenetically based establishment of a dengue virus panel, representing all available genotypes, as a tool in dengue drug discovery. Antivir. Res. 2019, 168, 109–113. [Google Scholar] [CrossRef]
- Suchard, M.A.; Lemey, P.; Baele, G.; Ayres, D.L.; Drummond, A.J.; Rambaut, A. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. 2018, 4, 16. [Google Scholar] [CrossRef] [Green Version]
- ABES. Associação Brasileira de Engenharia Sanitária e Ambiental ABES: Ranking ABES da Universalização do Saneamento. Available online: http://abes-dn.org.br/pdf/Ranking_2019.pdf%3E (accessed on 1 June 2020).
- Teixeira, T.R.D.A.; Medronho, R.D.A. Indicadores sócio-demográficos e a epidemia de dengue em 2002 no Estado do Rio de Janeiro, Brasil. Cad. Saude Publica 2008, 24, 2160–2170. [Google Scholar] [CrossRef] [Green Version]
- Vargas, W.P.; Kawa, H.; Sabroza, P.C.; Soares, V.B.; Honório, N.A.; De Almeida, A.S. Association among house infestation index, dengue incidence, and sociodemographic indicators: Surveillance using geographic information system. BMC Public Health 2015, 15, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kautner, I.; Robinson, M.J.; Kuhnle, U. Dengue virus infection: Epidemiology, pathogenesis, clinical presentation, diagnosis, and prevention. J. Pediatr. 1997, 131, 516–524. [Google Scholar] [CrossRef]
- da Brasil, M.S. Dengue diagnóstico e manejo clínico criança. Secr. Vigilância em Saúde 2016, 5, 1–56. [Google Scholar]
- World Health Organization. Dengue Guidelines for Diagnosis, Treatment, Prevention and Control: New Edition 2009; World Health Organization: Geneva, Switzerland, 2009. [Google Scholar]
- Halstead, S.B. Etiologies of the experimental dengues of siler and simmons. Am. J. Trop. Med. Hyg. 1974, 23, 974–982. [Google Scholar] [CrossRef] [PubMed]
- da Brasil, M.S. Boletim Epidemiológico, Monitoramento dos Casos de Arboviroses Urbanas Transmitidas pelo Aedes (Dengue, Chikungunya e Zika), Semanas Epidemiológicas 1 a 34, 2019; Ministério da Saúde: Brasília, Brazil, 2019; Volume 50. [Google Scholar]
- da Brasil, M.S. Boletim Epidemiológico, Monitoramento dos Casos de Dengue, Febre de Chikungunya e Febre Pelo Vírus Zika Até a Semana Epidemiológica 4, 2016; Ministério da Saúde: Brasília, Brazil, 2016; Volume 47, pp. 1–7. [Google Scholar]
- Ministério da Saúde Monitoramento dos casos de dengue, febre de chikungunya e febre pelo vírus Zika até a Semana Epidemiológica 5 de 2018. Bol. Epidemiol. 2018, 46, 43–2015. [CrossRef]
- Fried, J.R.; Gibbons, R.V.; Kalayanarooj, S.; Thomas, S.J.; Srikiatkhachorn, A.; Yoon, I.K.; Jarman, R.G.; Green, S.; Rothman, A.L.; Cummings, D.A.T. Serotype-specific differences in the risk of dengue hemorrhagic fever: An analysis of data collected in Bangkok, Thailand from 1994 to 2006. PLoS Negl. Trop. Dis. 2010, 4, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Franco, L.; Palacios, G.; Martinez, J.A.; Vázquez, A.; Savji, N.; de Ory, F.; Sanchez-Seco, M.P.; Martín, D.; Lipkin, W.I.; Tenorio, A. First report of sylvatic DENV-2-associated dengue hemorrhagic fever in West Africa. PLoS Negl. Trop. Dis. 2011, 5, e1251. [Google Scholar] [CrossRef]
- Cardosa, J.; Ooi, M.H.; Tio, P.H.; Perera, D.; Holmes, E.C.; Bibi, K.; Manap, Z.A. Dengue virus serotype 2 from a sylvatic lineage isolated from a patient with dengue hemorrhagic fever. PLoS Negl. Trop. Dis. 2009, 3, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Dhanoa, A.; Hassan, S.S.; Ngim, C.F.; Lau, C.F.; Chan, T.S.; Adnan, N.A.A.; Eng, W.W.H.; Gan, H.M.; Rajasekaram, G. Impact of dengue virus (DENV) co-infection on clinical manifestations, disease severity and laboratory parameters. BMC Infect. Dis. 2016, 16, 406. [Google Scholar] [CrossRef] [Green Version]
- Osanai, C.H. A Epidemia de Dengue em Boa Vista, Território Federal de Roraima, 1981−1982; Escola Nacional de Saúde Pública: Rio de Janeiro, Brazil, 1984. [Google Scholar]
- De Figueiredo, R.M.P.; Naveca, F.G.; Bastos, M.D.S.; Melo, M.D.N.; Viana, S.D.S.; Mourão, M.P.G.; Costa, C.A.; Farias, I.P. Dengue virus type 4, Manaus, Brazil. Emerg. Infect. Dis. 2008, 14, 667–669. [Google Scholar] [CrossRef]
- Walimbe, A.M.; Lotankar, M.; Cecilia, D.; Cherian, S.S. Global phylogeography of Dengue type 1 and 2 viruses reveals the role of India. Infect. Genet. Evol. 2014, 22, 30–39. [Google Scholar] [CrossRef]
- de Bruycker-Nogueira, F.; Mir, D.; dos Santos, F.B.; Bello, G. Evolutionary history and spatiotemporal dynamics of DENV-1 genotype V in the Americas. Infect. Genet. Evol. 2016, 45, 454–460. [Google Scholar] [CrossRef]
- Orchid, M.A.; Philippe, L.; Andrew, J.T.; Oliver, G.P.; Shannon, N.B.; Brandi, A.M.; Marc, A.S.; Jerome, E.F.; Andrew, R.; Christine, V.F.C. Phylogeography and population dynamics of dengue viruses in the Americas. Molecular Biol. Evol. 2012, 29, 1533–1543. [Google Scholar]
- Villabona-Arenas, C.J.; de Zanotto, P.M.A. Worldwide spread of dengue virus type 1. PLoS ONE 2013, 8, e62649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villabona-Arenas, C.J.; de Andrade Zanotto, P.M. Evolutionary history of Dengue virus type 4: Insights into genotype phylodynamics. Infect. Genet. Evol. 2011, 11, 878–885. [Google Scholar] [CrossRef]
- Weaver, S.C.; Vasilakis, N. Molecular evolution of dengue viruses: Contributions of phylogenetics to understanding the history and epidemiology of the preeminent arboviral disease. Infect. Genet. Evol. 2009, 9, 523–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.; Vasilakis, N. Dengue-Quo tu et quo vadis? Viruses 2011, 3, 1562–1608. [Google Scholar] [CrossRef]
- Dutra, K.R.; Drumond, B.P.; de Rezende, I.M.; Nogueira, M.L.; de Oliveira Lopes, D.; Calzavara Silva, C.E.; Siqueira Ferreira, J.M.; dos Santos, L.L. Molecular surveillance of dengue in Minas Gerais provides insights on dengue virus 1 and 4 circulation in Brazil. J. Med. Virol. 2017, 89, 966–973. [Google Scholar] [CrossRef]
- Nunes, M.R.T.; Faria, N.R.; Vasconcelos, H.B.; de Medeiros, D.B.A.; de Lima, C.P.S.; Carvalho, V.L.; da Silva, E.V.P.; Cardoso, J.F.; Sousa, E.C.; Nunes, K.N.B.; et al. Phylogeography of dengue virus serotype 4, Brazil, 2010–2011. Emerg. Infect. Dis. 2012, 18, 1858–1864. [Google Scholar] [CrossRef] [Green Version]
- Andrade, E.H.P.; Figueiredo, L.B.; Vilela, A.P.P.; Rosa, J.C.C.; Oliveira, J.G.; Zibaoui, H.M.; Araújo, V.E.M.; Miranda, D.P.J.; Ferreira, P.C.P.; Abrahão, J.S.; et al. Spatial-temporal co-circulation of dengue virus 1, 2, 3, and 4 associated with coinfection cases in a hyperendemic area of Brazil: A 4-week survey. Am. J. Trop. Med. Hyg. 2016, 94, 1080–1084. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5’–3’) | Position | DNA Size (pb) |
|---|---|---|---|
| D1 | TCAATATGCTGAAACGCGCGAGAAACCG | 134–161 | 511 pb |
| D2 | TTGCACCAACAGTCAATGTCTTCAGGTTC | 616–644 |
| Signs and Symptoms | Negative DENV | Positive DENV | |||||
|---|---|---|---|---|---|---|---|
| N = 288 | % | N = 20 | % | Chi-Square | p Value | Test t ANOVA | |
| CEFALEIA | 260 | 90.3% | 17 | 85.0% | 1.495 | 0.474 | 0.290 |
| FEVER | 258 | 89.6% | 20 | 100.0% | 1.619 | 0.203 | 0.205 |
| CHILLS | 243 | 84.4% | 19 | 95.0% | 1.139 | 0.286 | 0.288 |
| SWEATING | 175 | 60.8% | 12 | 60.0% | 0.069 | 0.792 | 0.793 |
| NAUSEA | 157 | 54.5% | 15 | 75.0% | 2.244 | 0.134 | 0.135 |
| VOMIT | 88 | 30.6% | 6 | 30.0% | 0.140 | 0.708 | 0.710 |
| MUSCLE PAIN | 255 | 88.5% | 18 | 90.0% | 0.000 | 0.988 | 0.988 |
| JOINT PAIN | 234 | 81.3% | 15 | 75.0% | 0.894 | 0.344 | 0.346 |
| DIARRHEA | 78 | 27.1% | 3 | 15.0% | 2.006 | 0.157 | 0.158 |
| DYSURIA | 47 | 16.3% | 4 | 20.0% | 0.027 | 0.869 | 0.869 |
| OLIGURIA | 64 | 22.2% | 2 | 10.0% | 2.248 | 0.134 | 0.135 |
| LACK OF APPETITE | 169 | 58.7% | 11 | 55.0% | 15.563 | 0.001 | 0.729 |
| RASH | 18 | 6.3% | 2 | 10.0% | 0.201 | 0.654 | 0.655 |
| STAINS | 24 | 8.3% | 2 | 10.0% | 0.003 | 0.953 | 0.954 |
| PRURITUS | 20 | 6.9% | 1 | 5.0% | 0.245 | 0.621 | 0.622 |
| DIZZINESS | 140 | 48.6% | 11 | 55.0% | 0.074 | 0.786 | 0.787 |
| VERTIGO | 86 | 29.9% | 8 | 40.0% | 0.384 | 0.536 | 0.537 |
| MENTAL CONFUSION | 29 | 10.1% | 1 | 5.0% | 0.843 | 0.359 | 0.360 |
| DYSPNEA | 62 | 21.5% | 5 | 25.0% | 0.027 | 0.870 | 0.871 |
| BLEEDING | 8 | 2.8% | 0 | 0.0% | 0.676 | 0.411 | 0.413 |
| ABDOMINAL PAIN | 133 | 46.2% | 6 | 30.0% | 2.767 | 0.096 | 0.097 |
| RETRO-ORBITARY PAIN | 174 | 60.4% | 14 | 70.0% | 0.722 | 0.697 | 0.428 |
| WEIGHT LOSS | 129 | 44.8% | 1 | 5.0% | 0.310 | 1.400 | 0.238 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Queiroz, J.A.d.S.; Botelho-Souza, L.F.; Nogueira-Lima, F.S.; Rampazzo, R.d.C.P.; Krieger, M.A.; Zambenedetti, M.R.; Marchini, F.K.; Borghetti, I.A.; Pereira, D.B.; Salcedo, J.M.V.; et al. Phylogenetic Characterization of Arboviruses in Patients Suffering from Acute Fever in Rondônia, Brazil. Viruses 2020, 12, 889. https://0-doi-org.brum.beds.ac.uk/10.3390/v12080889
Queiroz JAdS, Botelho-Souza LF, Nogueira-Lima FS, Rampazzo RdCP, Krieger MA, Zambenedetti MR, Marchini FK, Borghetti IA, Pereira DB, Salcedo JMV, et al. Phylogenetic Characterization of Arboviruses in Patients Suffering from Acute Fever in Rondônia, Brazil. Viruses. 2020; 12(8):889. https://0-doi-org.brum.beds.ac.uk/10.3390/v12080889
Chicago/Turabian StyleQueiroz, Jackson Alves da Silva, Luan Felipo Botelho-Souza, Felipe Souza Nogueira-Lima, Rita de Cássia Pontello Rampazzo, Marco Aurélio Krieger, Miriam Ribas Zambenedetti, Fabricio Klerinton Marchini, Ivo Alberto Borghetti, Dhelio Batista Pereira, Juan Miguel Vilalobos Salcedo, and et al. 2020. "Phylogenetic Characterization of Arboviruses in Patients Suffering from Acute Fever in Rondônia, Brazil" Viruses 12, no. 8: 889. https://0-doi-org.brum.beds.ac.uk/10.3390/v12080889