Enteric Viral Co-Infections: Pathogenesis and Perspective


Division of Infectious Diseases, Department of Medicine, Edison Family Center for Genome Sciences & Systems Biology, Washington University School of Medicine, St. Louis, MO 63110, USA
*
Author to whom correspondence should be addressed.
Viruses 2020, 12(8), 904; https://0-doi-org.brum.beds.ac.uk/10.3390/v12080904
Submission received: 13 July 2020
/
Revised: 10 August 2020
/
Accepted: 14 August 2020
/
Published: 18 August 2020
(This article belongs to the Special Issue Viral Coinfection)
Abstract
:Enteric viral co-infections, infections involving more than one virus, have been reported for a diverse group of etiological agents, including rotavirus, norovirus, astrovirus, adenovirus, and enteroviruses. These pathogens are causative agents for acute gastroenteritis and diarrheal disease in immunocompetent and immunocompromised individuals of all ages globally. Despite virus–virus co-infection events in the intestine being increasingly detected, little is known about their impact on disease outcomes or human health. Here, we review what is currently known about the clinical prevalence of virus–virus co-infections and how co-infections may influence vaccine responses. While experimental investigations into enteric virus co-infections have been limited, we highlight in vivo and in vitro models with exciting potential to investigate viral co-infections. Many features of virus–virus co-infection mechanisms in the intestine remain unclear, and further research will be critical.
1. Introduction
Every year, millions of children are infected with viruses that target the gastrointestinal tract to cause acute gastroenteritis (AGE), which is the inflammation of the stomach or small or large intestine [1,2]. Indeed, approximately one billion episodes of diarrhea occur in children under five annually [3]. Symptoms accompanying AGE include malaise, abdominal pain and cramping, nausea, vomiting, and diarrhea, usually lasting 1 to 5 days, but occasionally up to 14 days [4]. RNA viruses, including rotavirus (RV), norovirus (NoV), sapovirus (SaV), astrovirus (AstV), and enteroviruses that spread via fecal–oral transmission, as well as DNA viruses, including enteric adenoviruses (AdV), are associated with AGE in immunocompetent and immunocompromised individuals of all age groups [5] (Table 1).
In the past, AGE was generally attributed to a single causative agent known to be associated with clinical symptoms. However, due to their high prevalence, exposure to multiple viruses at similar times can potentially occur in the same host, and these viruses may either infect simultaneously or within a short window of time. The outcomes of either infection may thus be influenced by the combined contribution of multiple agents. With increasing capacity to efficiently analyze samples for the presence of multiple putative pathogens using quantitative real-time PCR approaches, our appreciation that more than one virus may be present in an infected individual has grown. Enteric virus co-infections are commonly identified for all viruses linked to AGE [14] (Table 1). Despite our increased awareness of the prevalence of enteric viral co-infections, however, the mechanisms underlying viral co-infection have not yet been carefully explored. There are significant gaps in our knowledge regarding the dynamics between co-infecting agents and how these influence disease severity, immune responses, or vaccine efficacy.
Here, we will review what is known about the clinical aspects of enteric virus co-infection and discuss potential models to investigate viral co-infection in vitro and in vivo. We will first summarize what is known about enteric viral co-infection prevalence and its impact on health and disease, as well as how vaccine responses may be influenced by co-infection events. Second, we will present several of the possible cellular and molecular consequences of enteric viral co-infections. Finally, we will conclude by exploring past and current model systems used to study viral infections in the gut that could be applied to co-infections.
2. Virus–Virus Co-Infection Prevalence
By quantity and diversity, prokaryotic and eukaryotic viruses in the human intestinal microbiota likely outnumber their bacterial counterparts, with the size of the virome still remaining largely unknown [15,16,17]. In this review, we will predominantly focus on intestinal eukaryotic viruses as these are the known causative agents of AGE, but will briefly mention that co-infection relationships between bacteriophages play a major role in shaping complex microbial ecosystems [18,19]. Superinfection exclusion serves to limit subsequent phage infection of bacteria already having a prophage present, but in other contexts virus–virus interactions may serve to facilitate infections especially when the co-infecting phage are highly distinct (for example, ssDNA and dsDNA phage) [20]. While phage co-infection interactions have predominantly been explored in vitro or in environmental samples, new evidence suggests phage co-infection is also occurring in the human gastrointestinal tract, and thus the interrogation of these interactions is likely to be an important aspect of future microbiota research [21].
Recently, the metagenomic sequencing analysis of stool samples of several cohorts of healthy neonates revealed a stepwise viral colonization model wherein prophages may be the viral pioneers in the gut at one month after birth while, by four months, eukaryotic viruses became prominent [22,23,24]. The first several years of life are then a particularly dramatic period of co-infection, and the simultaneous detection of multiple enteropathogens is common, both associated with AGE and even in the absence of symptoms. As an example of prevalence in early life, a cohort of Dutch infants had at least one enteropathogen, including eukaryotic viruses and bacterial pathogens, detected in ~73% of samples tested, and of these, nearly half had two distinct pathogens detected [25]. Similarly, a longitudinal study conducted in two healthy British infants during their first year of life showed that a variety of human viruses, including AdV, AstV, RV, and multiple enteroviruses, were present during this period [26]. Indeed, 92% of the samples tested in this study were positive for one to five different viruses, and approximately half of the samples contained two or more viruses. Gut pathogens, including enteric viruses, are thus extremely common in early life, and virus–virus interactions, apart from being synergistically pathogenic, may play important roles in the development of the host–virome homeostatic relationship, as well as immune system education [15].
Risk factors for acquiring enteric co-infections include young age, daycare attendance, and households with more than three children [25], as well as contaminated drinking water and poor sanitation [27]. Children from high income regions often exhibit lower rates of co-infection than those from lower income regions. For example, using the same assay, one French study found a 7% co-infection rate, compared to a 77% co-infection rate observed in a cohort of Ghanaian children [28,29,30]. In one of the largest multi-site studies to look at pathogens associated with community diarrhea in developing countries, one enteropathogen was detected in 77% of diarrheal stools and 65% of non-diarrheal samples, and two or more pathogens in 41 and 29%, respectively, in the first two years of life [31]. Thus, an improved understanding of the consequences of these extremely common co-infection events is critical. Below, we will briefly describe the major enteric viral pathogens, and then will detail what has been discovered about viral co-infection frequency worldwide.
Reoviridae: Despite the successful implementation of vaccine programs against rotavirus (RV) starting in 2006, it continues to be the leading cause of viral diarrheal mortality in children under the age of 5 globally, causing approximately 200,000 deaths per year [32]. To date, there are two internationally licensed oral RV vaccines [33,34]: the monovalent Rotarix and the pentavalent RotaTeq. Additional newer-generation RV vaccines are licensed in individual countries [33]. RV group A is the most common cause of AGE in countries without effective RV vaccination programs, causing significant childhood morbidity and mortality. RV is generally understood to target the mature enterocytes of the small intestine, likely mediating diarrheal illness via both direct effects on the epithelium and indirect effects from host responses [35].
Caliciviridae: Norovirus (NoV) has emerged as a leading cause of pediatric gastroenteritis [36], in addition to its well-established role in epidemic diarrheal disease in adults in healthcare settings and cruise ships [37]. NoV GII strains are the most common viral cause of epidemic AGE worldwide, with GII.4 strains most frequently associated with person-to-person transmission, due to their extremely contagious nature and their rapid rate of genetic and antigenic evolution [38]. Indeed, NoV causes ~18% of all cases of diarrhea in outpatient and inpatient settings among all age groups [39]. NoV has been shown to infect both immune cells and enteroendocrine cells of the small intestinal epithelium in immunocompromised patients [40,41]. Though less widely appreciated as a cause of AGE, sapovirus (SaV) is another member of the Caliciviridae family that causes diarrheal illness similar to NoV, especially in young children and the elderly, and frequently closely follows NoV in prevalence [42,43]. Although there are several NoV vaccines in development and two in clinical trials (bivalent GI.1/GII.4 vaccine and monovalent GI.1 oral vaccine) [44,45], there are no licensed vaccines against NoV or SaV.
Astroviridae: Astroviruses (AstV) are causative agents of diarrheal disease particularly in children and immunocompromised patients. Few studies report AstV-mediated AGE in normal healthy adults, but most children are infected with AstV and develop antibodies to the virus early in life [46,47]. Classical AstV has eight serotypes (AstV1–8) that usually cause self-limiting infections but can spread systemically in immunocompromised individuals [48]. Some novel strains of AstV have also recently been associated with central nervous system infections, mostly in immunocompromised patients, but their association with AGE is not well-established [49,50,51,52]. The replication of human AstV in enteroids in vitro indicates a multi-cellular tropism, including both intestinal progenitor cells and mature enterocytes [53].
Picornaviridae: Enterovirus species A and B (enteroviruses and coxsackieviruses), and C (polioviruses (PV)) are prevalent enteric pathogens causing a range of diseases, including AGE. PV is an enteric virus transmitted through the fecal–oral route, which during the pre-vaccine era could spread from the gut to the central nervous system to cause poliomyelitis, a disease leading to temporary or permanent paralysis [54,55]. Following the launch of the Global Polio Eradication Initiative in 1988, global vaccination programs led to the eradication of the indigenous wild PV type 2 in 2015 and type 3 in 2019 [56]. PV vaccines include an inactivated PV (developed by Jonas Salk) administered via injection [57] and a live attenuated PV (developed by Albert Sabin) administered orally—the oral polio vaccine (OPV) [58]. Both vaccines show high efficacy, but OPV is considerably easier to deploy in low-resource settings around the world and confers longer-lasting community immunity [59]. However, in rare cases, OPV can revert to its pathogenic variant and provoke poliomyelitis [60], and recent vaccine-derived PV outbreaks have been documented in several countries, highlighting the importance of continued PV research [61]. Other human enteroviruses, including enterovirus-A71, coxsackievirus-A6 and coxsackievirus-A16, cause hand, foot, and mouth disease (HFMD), predominantly in children aged 1 to 3 years old worldwide [62]. Enterovirus-A71 is closely associated and co-circulates with coxsackievirus-A16, causing seasonal outbreaks of HFMD in Central China, with reported co-infection in ~2% of cases [63]. In general, co-infection between enterovirus-A71 and other enteroviruses during HFMD outbreaks are associated with more severe symptoms [63,64,65]. While human enteroviruses have not classically been associated with AGE, there is increasing appreciation that many of these viruses can be associated with pediatric diarrhea [65,66,67,68].
Adenoviridae: Adenoviruses (AdV) are non-enveloped DNA viruses known to cause conjunctivitis, upper and lower respiratory disease, and AGE. AdV can be transmitted via respiratory droplets, fomites, and fecal–oral transmission, and predominantly infects the respiratory and gastrointestinal epithelia [69]. Epidemiological data indicate that the majority of AdV infections occur in children less than 5 years of age, but AdV epidemics are also common in adults. AdV genotypes 40 and 41 are commonly associated with AGE in pediatric populations worldwide [70,71,72,73,74]. The intestinal tract also appears to be a common site of AdV reactivation under conditions of immunosuppression [75]. In patients with congenital immunodeficiencies, AdV can cause disseminated and other lethal disease [76,77,78], whereas in acquired immunodeficiency associated with HIV infection, AdV infections are more commonly associated with AGE [79]. Although AdV infection in the respiratory tract is well characterized, gastrointestinal infection is less well-understood in terms of cell tropism, entry mechanism and intestinal immune responses [75]. AdV is prevalent in intestinal biopsies from healthy individuals, with co-infection by multiple strains commonly detected. AdV DNA has been found in lamina propria lymphocytes, but the full cellular tropism of human AdV remains unclear [80].
Coronaviridae: Coronaviruses have been previously considered as possible rare causes of AGE in infants [11]. However, the end of 2019 witnessed the beginning of an ongoing global pandemic from novel severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), with nearly 19 million cases and >706,000 deaths to date (as of 6 August 2020), dramatically increasing the awareness of these viral pathogens. SARS-CoV-2 uses receptor protein ACE2 for entry, which is expressed on lung epithelium but also on enterocytes in the gastrointestinal tract [12,13]. Infection is often associated with gastrointestinal symptoms including diarrhea, and SARS-CoV-2 viral RNA can be detected in fecal samples and viral antigens identified in intestinal tissues of COVID-19 patients [81,82,83]. In addition, there is mounting evidence for the SARS-CoV-2 antigen in the intestines of animal models, such as macaques and ferrets [84,85]. Although SARS-CoV-2 infection exhibits milder clinical symptoms in children as compared to adults, prolonged fecal shedding has been observed in pediatric cases [83,86]. It has already been suggested that SARS-CoV-2 infection may prevent co-infection by other respiratory viruses [87]. While it is still unclear if SARS-CoV-2 exhibits fecal–oral transmission [88,89], the epidemic has raised important questions about this novel pathogen in the intestine and whether it may be an important future consideration in enteric viral co-infections.
2.1. Co-Infection in Acute Gastroenteritis (AGE)
Though co-infections are more commonly thought of as viruses from different viral families mediating simultaneous infection, they can, of course, also include concurrent infections by multiple viral strains or species of the same viral genus. An investigation by the Rota-net Italy Study Group demonstrated that mixed infections with two or more RV strains were present in 7.6% of their samples, with G1 + G9P(8) strains representing the most common mixed infections in children with AGE [90]. In a study investigating the extensive RV genomic variation during chronic infection in immunocompromised children, patients unable to clear their initial RV infection were subsequently co-infected with other circulating RV strains leading to prolonged life-threatening diarrhea [91]. Mixed co-infections with different NoV genogroups have also been reported, as young children may experience multiple and often repeated NoV infections with remarkable genetic diversity [92,93,94]. Mixed enterovirus co-infections have not been frequently detected in AGE [68,95], though they have in HFMD [96], but as enteroviruses are now increasingly appreciated as causative agents for AGE, increased testing may also increase co-infection detection rates. Continuously improving techniques for carefully genotyping enteric viruses in clinical samples will likely continue to reveal frequent co-infections by closely-related viruses.
During epidemic AGE outbreaks, such as those that occur from contaminated water, co-infection with multiple unrelated viruses can be common. For example, in a pediatric AGE outbreak in Finland, in which drinking water was contaminated by sewage, RV was detected in 66% of cases, calicivirus including either NoV or SaV in 62%, and both in 40% [97]. Similarly, 23.2% of samples were positive for multiple enteric viruses in two outbreaks of AGE in Mumbai [98]. Even in community AGE, however, co-infections make up a moderate proportion of cases and are likely under-reported as some etiologic agents may not be tested for across all studies.
In analyses pre-dating the RV vaccine, one study conducted in Spanish children found that ~5% of the cohort showed mixed infections associated with acute diarrhea. Virus–virus co-infections were more frequent, with RV–AstV and RV–AdV being most common, than virus–bacteria co-infections [99]. Similarly, a study interrogating the prevalence of viral infections among children with AGE in France found co-infections in 50/299 positive samples (16.7%), wherein 94% of the cases were combinations of RV with NoV, AstV, or AdV in children under the age of 15 months [100].
The introduction of the RV vaccine altered the prevalence of RV infection, but it continues to be frequently detected during AGE. A study of enteric RNA virus co-infections in diarrheic children and adults in Southwestern Canada found that 2.8% of total RNA virus-positive samples showed combinations of NoV GII-SaV, NoV GII-RV, NoV GII-AstV, SaV-AstV, and SaV-RV at varying percentages [101]. A similar study focused on pediatric AGE in Spain identified more than half of children as being infected with at least one virus, including AdV, RV, SaV, AstV, and NoV, with co-infections detected in 21% of cases [102]. In Turkish children under the age of 5, RV was present in about one-third of AGE samples, and co-infections were found in 10.4% of samples with RV-NoV and NoV-AdV as the most frequently observed [8]. In China, RV and NoV-GII co-infections were reported in 14% of children suffering from severe diarrhea [39]. A study conducted to determine the agents associated with AGE in children in Nepal reported that nearly all children tested had at least one enteric viral agent detected, and approximately half were co-infected with RV and enterovirus [27]. Overall, these studies support that in healthy children and adults experiencing AGE episodes, between ~3–50% may have more than one viral pathogen present. Some reports indicate that RV co-infections, at least in healthy children, are not associated with greater symptom severity than single RV infection [25,27], while another reported that co-infections with NoV may lead to more severe AGE with enhanced comorbidities, such as vomiting and fever, compared to NoV alone [94]. Further clinical studies exploring the effects of co-infection on symptom severity are needed to better clarify the relative contribution of co-infection to disease outcomes.
Even less clear, however, are the impacts of co-infection in immunocompromised patients. Immunocompromised patients have been reported to have only slightly higher prevalence of individual enteric viral pathogens as the general population [103,104], but may experience both more symptomatic and prolonged infections [103,105]. A potential consequence of prolonged infection in immunocompromised patients and the elderly may be the intra-host evolution of genetically distinct viral populations and viral spread in nursing homes [106,107,108]. A study comparing HIV-positive and HIV-negative children reported a higher prevalence of AstV and enterovirus co-infection in HIV-infected children, with multiple strains of AstV detected in HIV-infected children with or without AGE [109]. An investigation in HIV-seropositive and -seronegative children with diarrhea in Brazil reported a significantly higher rate of co-infections with >1 enteric virus including NoV, AdV and human bocavirus in HIV-1 seropositive children [110]. There have also been a number of case reports of severe co-infections in immunocompromised individuals, such as those of AdV and coxsackievirus detected in a child with a primary immunodeficiency with persistent severe diarrhea [111], or AdV and NoV co-infection reported in a patient diagnosed with chronic myeloid leukemia undergoing stem cell transplantation [112]. Certainly, immunocompromised patients with AGE are at increased risk of complications and prolonged hospitalizations. The additional study of co-infections in both immunocompromised and elderly cohorts may help to provide insights into mechanisms related to the immune regulation of enteric viruses.
2.2. Co-Infection in Asymptomatic Individuals
Both human gut virome studies and investigations specifically testing for viral pathogens have shown that individuals with detectable enteric virus loads in stool may be free of clinical symptoms or disease pathology [15,16,26,113]. The long-term shedding of AstV, RV, AdV, and enteroviruses was detected in healthy infants in the UK from 1 up to 12 months without any clinical symptoms for AGE [26]. Asymptomatic infection and prolonged shedding of NoV is also common [36,92,114].
While there are fewer studies that have examined viral co-infection in asymptomatic individuals, those that have support frequent enteric viral co-infection in the absence of AGE. Up to 17 different enteric viruses were detected in fecal samples from children without diarrhea in Bangladesh, and among the positive samples, 53% demonstrated co-infection with multiple viruses [115]. Similarly, in a study in Cameroon, 53.7% of fecal specimens sampled from asymptomatic healthy children contained at least one identifiable enteric virus, with co-infections detected in 35.2%, almost all including NoV [116].
While co-infections are thus prevalent even in the absence of symptoms, the literature does suggest that, in matched cohorts, co-infection rates tend to be higher in cases of AGE than in asymptomatic controls. The analysis of samples from a Japanese daycare revealed that 1.6% of stools from asymptomatic children exhibited viral co-infections, whereas viral co-infections accounted for 5.7% of AGE cases [117]. Similarly, in southwest China, RV-NoV GII co-infection was found in 1.1% of asymptomatic children tested, as compared to 4.4% of samples from AGE cases [39]. Continued sampling of asymptomatic pediatric cohorts in diverse geographic locations will be important to help more fully define the worldwide prevalence of viral co-infections in the absence of symptoms.
2.3. Co-Infection Effects on Vaccines and Treatments
As discussed above, vaccines are currently available for PV and RV, but not for other enteric viruses associated with AGE. These vaccines are administered in early childhood, which is also the most common period for viral co-infections to occur. The inhibitory effect of human enteroviruses on OPV efficacy has been recognized for many decades, and indeed has contributed to attempts to vaccinate children in months when these viruses were circulating at lower levels [118,119]. A systematic review supports that concurrent enterovirus infection is consistently associated with decreased seroconversion for some types of PV, and in general that concurrent diarrhea which could be attributed to different enteropathogens is associated with decreased per-dose seroconversion overall [120]. Recently acquired enterovirus infections seem to have more dramatic effects in preventing immune response development than persistent infections [121]. In addition to enterovirus infections inhibiting effective immune responses to OPV, there have been more recent studies suggesting these may also impair effective RV vaccine responses. Diminished RV IgA and failure to seroconvert after RV vaccination were associated with concurrent enterovirus infections in a cohort of infants in urban Bangladesh [122]. Importantly, the co-administration of OPV and RV vaccines themselves have also been suggested to inhibit effective vaccine responses [123,124].
While some recent studies have suggested that co-infections with bacterial or viral enteropathogens do not alter RV vaccine efficacy in a statistically significant manner, across these studies there is a trend for an 8–11% increase in vaccine efficacy against severe RV diarrhea in children who were not co-infected [30,125,126]. Thus, it remains possible that viral co-infections are an important contributor to decreased vaccine efficacy in settings with high enteropathogen burdens. Further work will be needed to fully clarify the impact of enteric viral infections on the induction of effective protective responses with vaccination, and will continue to be an important consideration as vaccines are developed for other enteric viruses, such as NoV.
While most enteric viruses causing AGE do not have specific treatments available, an important note is that frequently diarrheal episodes are treated with antibiotics which have no efficacy against viral pathogens. In the MAL-ED multi-site study, almost half of all diarrheal episodes were treated with antibiotics. Analysis later showed that 91.7% of these antibiotic courses were inappropriate [3]. Thus, improved recognition and detection of the enteric viruses that mediate AGE, as well as an appreciation that multiple viruses may be present to drive diarrheal disease, is increasingly important to limit unnecessary administration of antibiotics.
3. Cellular and Molecular Consequences of Co-Infection
Despite frequent reports of viral co-infections, their consequences and underlying mechanisms are poorly understood. Here, we discuss potential consequences of co-infections, suggestions which will need to be bolstered by careful future studies (Figure 1).
3.1. Viral Recombination or Reassortment
Co-infections with related strains facilitate viral recombination, which can occur via homologous or non-homologous recombination, or reassortment. Improvements in viral genomic sequencing methods have facilitated the detection of recombinants, leading to accelerated detection of these events over the past decade. Enteroviruses have been frequently observed to recombine to permit the emergence of new viral strains [127,128], and indeed OPV and circulating enteroviruses also have the capacity to generate recombinants with enhanced pathogenicity and fitness [129]. Recombination events are also frequent in human and animal AstV [130] and NoV [131]. Because RV is a segmented RNA virus, a concerning outcome for co-infection is reassortment of the viral genome that can facilitate the emergence of novel epidemic strains [132]. While there has been less evidence for AdV recombination during intestinal infection, it is certainly a possible outcome considering the frequently persistent presence of these viruses in the gastrointestinal tract [75]. Cross-family viral recombination events are much less likely, but certainly not impossible [133].
3.2. Virus Aggregates
Recent studies have revealed that multiple enteric eukaryotic viruses, including RV, NoV, and PV may be transmitted in aggregates or viral clusters [134,135]. Groups of assorted virions, often containing mutagenized genomes, are replicated and transmitted cell-to-cell in vesicle-like structures [135,136,137]. For aggregation to occur, virions must be able to accumulate and be transmitted from infected cells to neighboring cells with minimal interference. Aggregation may offer viruses multiple advantages, including an increased cellular multiplicity of infection [138], and greater opportunity for complementation or recombination to restore fitness [135]. Gut bacteria have been shown to facilitate the aggregation of PV, and this could be a mechanism of the bacterially-mediated enhancement of infection by other enteric viruses [139]. Fundamentally, aggregates serve as an important and newly-appreciated mechanism for the co-infection of multiple related virions. Whether aggregates have the potential to include multiple unrelated viruses, and their role in the cellular consequences of co-infections, remains to be seen.
3.3. Viral Interference
Viral interference is a phenomenon first recognized in the 1950’s, wherein a cell infected by a virus is resistant to a second super-infecting virus, mediated by interferons (IFNs) [140]. This protection is also conferred to neighboring cells. Viral interference has also been proposed as an important mechanism by which enterovirus co-infections limit OPV and RV vaccine efficacy [120]. While few studies have interrogated enteric viral co-infections in mice to date, we recently uncovered viral interference as an unexpected outcome in immunocompromised mice. We observed that chronic murine AstV infection in severely immunodeficient mice protected mice from both persistent murine NoV and murine RV infection [141]. This protection was transferable to other immunodeficient mice and was associated with the upregulation of IFN-lambda (IFN-λ) in the gut, which in turn protected the mice from infection by other enteric viruses [141]. Further details of this intrinsic defense mechanism remain to be uncovered, as do the nuances of differential viral strain effects, cell type-specific IFN-λ expression, and positive and negative regulation of heterologous antiviral responses. It is likely this is an important mechanism regulating viral outcomes in the intestine.
4. Model Systems to Study Virus Co-Infections
While mechanistic explorations of viral co-infections in the intestine have been limited, there are several useful model systems available to explore the pathogenesis of individual viruses which could be applied to co-infection studies (Figure 2). Here, we describe these models, and suggest that these may permit us to address remaining fundamental questions: How does co-infection alter the lifecycle, replication, and tropism of individual viruses? Do individual viruses promote or prevent infection by subsequent viral agents? Do co-infections ultimately alter disease outcomes? The use of animal models and in vitro tools will be key to dissecting the molecular underpinnings in enteric virus co-infections.
4.1. Non-Human Primates
Non-human primates (NHPs), including Vervet monkeys (Chlorocebus pygerythrus), Rhesus macaques (Macaca mulatta), and pigtailed macaques (Macaca nemestrina) have been used extensively to study enteric viral infections [142]. NHPs share many anatomical, immunological, and physiological similarities with humans and thus are excellent models to recapitulate the pathogenesis of enteric virus infections and/or co-infection in humans [143]. NHPs are susceptible to some human enteric viruses, making studies of human viral co-infections potentially tractable [144,145]. Indeed, many simian viruses isolated from NHPs in the wild or in captivity, including simian RV and rhesus enteric caliciviruses, are also important pathogens for in vitro and in vivo studies, such as viral receptor identification and determination of adaptive immune responses [146,147,148]. In fact, research in simian RV led to the formulation of the first commercial vaccines against human RV [148,149]. Virome studies in NHPs suggest that, especially when immunocompromised by infection with simian immunodeficiency virus, natural enteric viral co-infections may be common [150,151,152]. However, experimental co-infection with enteric viruses remains a relatively unexplored area.
4.2. Pig Models
Gnotobiotic pigs serve as a useful model to study human enteric viruses, as similarities to human gastrointestinal physiology and mucosal immune development provide significant advantages over other model systems [153,154]. The capacity to introduce defined microbial consortia and/or study the effects of infection in the absence of the microbiota also add a layer of utility to these models. Piglets have been used extensively to study human RV and NoV infection, as they exhibit diarrheal disease, gastroenteritis, and fecal viral shedding, and have been useful for antibody response studies [153,155]. For example, the detection of human NoV capsid protein in small intestinal enterocytes of infected pigs associated with increased apoptosis and epithelial barrier disruption may offer insights into diarrheal disease observed in humans [156]. In the last few decades, multiple porcine enteric viruses that cause acute diarrhea, vomiting, dehydration, and high mortality in neonatal piglets have been identified, including Coronaviridae family members, porcine epidemic diarrhea virus and porcine deltacoronavirus, as well as porcine NoV similar to human NoV GII strains [157,158,159]. These viruses show fecal–oral transmission similar to human enteric viruses, providing an opportunity to investigate the nuances of viral transmission and define factors controlling clinical outcomes [160]. Recently, enteric virus co-infections were detected in the swine industry in China wherein piglets with diarrhea showed higher rates of coinfections with porcine epidemic diarrhea virus and porcine kobuvirus [161]. Thus, pigs have substantial potential as models to study enteric virus co-infections.
4.3. Mouse Models
In a compelling survey of the gut virome to investigate whether house mice living in close proximity with humans in residential buildings carry human pathogens, multiple viruses from the Parvoviridae, Picobirnaviridae, Astroviridae, Genomoviridae, and Circoviridae families were identified [162]. Although this study did not find any human viral pathogens, it highlighted a diverse viral ecology consisting of various enteric viruses in the mouse gut, analogous to the enteric viral diversity observed in humans.
Laboratory mouse models (specific knockouts, immunodeficient, transgenic, etc.), which can be maintained free of some of these enteric pathogens, have been highly instrumental in studying enteric viral infections including pathogenesis and immune responses. Mice are easy to manipulate and can recapitulate many of the human enteric viral–host relationships [163]. Human NoV GII.4 has been shown to replicate in a BALB/c Rag-γc-deficient mice, suggesting that some human enteric viruses may directly infect small animal models to facilitate study [164], though the further development of mice susceptible to human enteric virus infection is an important target. The discovery of murine NoV in immunodeficient mice provided the opportunity to study NoV in a natural host and contributed immensely to the understanding of the cellular and molecular mechanisms of NoV pathogenesis [165,166,167,168,169]. While murine NoV does not cause diarrheal illness in adult mice, it was recently reported to induce diarrhea in some neonatal models [170]. Comparably, murine RV causes both diarrhea in neonatal mouse pups and spreads among littermates [171,172]. Recently, murine AstV were discovered in immunocompetent, as well as in chronically infected immunodeficient, mice [163,173,174]. These murine AstV generally cause asymptomatic infection and minimal pathology [163]. While natural transmission of these viruses is fecal–oral, administration of virus via alternate routes permits comparative studies and can provide useful insights into pathogenesis and antiviral mechanisms [166,175]. Importantly, studies using murine viruses have revealed that viral strains differing by only a few amino acids may have dramatically different in vivo outcomes, in cellular tropism, duration of infection, and in interactions with the host innate and adaptive immune systems [176,177,178]. These observations help to inform our understanding of strain-specific differences in human viruses as well. Thus, although mouse viral models may not mirror all clinical features of human enteric infections, they are useful in identifying key determinants of viral regulation in vivo.
Overall, there is a clear need, and opportunity for, the exploration of enteric viral co-infections in mouse models, in order to interrogate infection outcomes for individual viruses as well as innate and adaptive immune responses and how these are governed by the presence of multiple viruses during infection.
4.4. Drosophila melanogaster
Insect models, including the fruit fly D. melanogaster, can be used to study oral infection by bacterial and viral pathogens in well-characterized, fast-growing, and readily genetically-manipulatable hosts [179,180,181]. Drosophila C virus, related to mammalian enteric picornaviruses, and Flock House Virus, a persistent RNA virus, are natural enteric viral infections in D. melanogaster. Microbiota regulation of antiviral immunity can restrict enteric viral infections in D. melanogaster [182], supporting that multiple aspects of enteric viral infection in higher organisms can be effectively modeled in flies. Further, Drosophila can be infected orally by human arboviruses, permitting the exploration of both fly and human virus co-infections [183]. Taken together, Drosophila has important promise as a model for enteric viral co-infections.
4.5. Caenorhabditis elegans
The nematode C. elegans is the model organism with the most biologically simple intestine, comprised of just 20 clonal enterocytes [181]. C. elegans has a number of advantages, as it has a rapid generation time and can be efficiently screened using forward- and reverse-genetic, as well as drug or small molecule screening approaches. It has been instrumental in the discovery of multiple molecular pathways, such as RNA interference [184]. There are several natural enteric viruses that have been recently isolated from wild-caught nematodes, including Orsay virus, facilitating the study of host–virus interactions [185]. Further, C. elegans are potentially amenable to infection with non-natural nematode viruses via microinjection [186]. C. elegans is thus a potential model to identify host-pathogen interactions during enteric virus co-infections.
4.6. Immortalized In Vitro Cell Lines
Caco-2 and HT-29 lines, derived from human colorectal adenocarcinomas, are the most widely used in vitro immortalized cell systems to study mechanisms underlying host–pathogen interactions in the intestine [187,188]. The Caco-2 cell line becomes highly polarized when subjected to optimal conditions and can thus differentiate to resemble small intestine-like cells or remain undifferentiated as large intestinal cells [188,189]. This cell line has been used to cultivate a number of enteric viruses, including human NoV [190,191], human RV [192,193] and human AstV [194,195], with some success. Interestingly, Caco-2 cells have been used to perform limited co-infection studies, revealing that mixed virus infection with AstV or enterovirus interferes with RV replication [196]. Much like Caco-2 cells, HT-29 cells have been employed extensively for studies of different aspects of intestinal epithelial cell biology [197,198]. HT-29 have been used for human AdV-40 and -41 [199] and a variety of RV studies [200,201]. These cell lines thus offer highly tractable systems to interrogate virus–virus co-infections in intestinal epithelial cells.
4.7. Intestinal Organoids and Enteroids
In the last decade, pioneering work from several groups has demonstrated the capacity of pluripotent intestinal stem cells to develop into complex intestinal structures in cell culture, closely mimicking their in vivo counterparts. Two types of intestinal organoids are used for studying enteric viruses—organoids derived from pluripotent stem cells, known as human intestinal organoids (HIOs), or organoids derived from intestinal crypts grown in three dimensional cultures, known as human intestinal enteroids (HIEs) [202,203,204,205].
Both HIOs and HIEs are widely used to study enteric viral pathogenesis and innate immune responses in the intestinal epithelium. HIOs support the replication of clinical isolates of RV, and HIEs have been used to study host restriction and innate antiviral responses to RV [206,207]. Multiple human NoV strains have been successfully cultivated in HIEs derived from human intestinal biopsies [208], as have human AstV [53]. HIE models have also been pivotal in investigating antiviral signaling responses to enteroviruses and coxsackieviruses [209,210]. HIOs and HIEs thus offer efficient systems to cultivate previously non-cultivable human enteric viruses, and thus may be powerful systems to investigate enteric virus co-infections.
5. Conclusions and Future Directions
Single pathogens have been classically understood to be the etiologies of AGE. As the evidence for virus–virus co-infections grows, however, it is evident that the causative agents for this disease are often co-infecting viruses, and indeed that these co-infections may be occurring even in the absence of symptoms. However, our understanding of the consequences of enteric virus co-infection is still in its infancy. There is a great remaining need for careful exploration of the effects of co-infection on symptom severity and duration, as well as development of protective immunity, in clinical cohorts.
Notably, the frequency of co-infection, and the identity of implicated viruses, can be drastically altered with the introduction of immunization programs. For example, PV and RV vaccination programs dramatically reduced infection rates in communities worldwide. Thus, as vaccines are developed for other enteric viral pathogens, there may continue to be robust shifts in viral co-infection prevalence. We are also now developing an increasing appreciation that the commensal microbiota, diet, and host genetics may have complex regulatory effects on enteric viruses, all of which may variably contribute to disease outcomes from infection as well. Thus, there remains much to be discovered in terms of physiological effects of co-infections and other host factors on disease severity, immune defense mechanisms and vaccine responses. Although the different models discussed in this review have been used to study the pathogenesis of individual enteric viruses, our knowledge regarding enteric viral co-infections lags far behind. We suggest that these models could be used as a toolkit for targeted studies exploring clinically relevant enteric virus co-infections, thereby allowing us to carefully dissect the individual contributions of the many complex factors that control these pathogens.
Author Contributions
H.M., H.I., and M.T.B. wrote, read, and edited the manuscript. All authors have read and agreed to the final version of the manuscript.
Funding
H.I. was supported by the Children’s Discovery Institute of Washington University and St. Louis Children’s Hospital Postdoctoral Research grant (MI-F-2018–712). M.T.B. was supported by NIH grants R01 AI127552, R01 AI139314, and R01 AI141478, a Children’s Discovery Institute of Washington University and St. Louis Children’s Hospital Interdisciplinary Research Initiative grant (MI-II-2019–790), the Pew Biomedical Scholars Program, and The Mathers Foundation.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Parashar, U.D.; Hummelman, E.G.; Bresee, J.S.; Miller, M.A.; Glass, R.I. Global illness and deaths caused by rotavirus disease in children. Emerg. Infect. Dis. 2003, 9, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Collaborators, G.B.D. Estimates of global, regional, and national morbidity, mortality, and aetiologies of diarrhoeal diseases: A systematic analysis for the global burden of disease study 2015. Lancet Infect. Dis. 2017, 17, 909–948. [Google Scholar] [CrossRef]
- Mills, J.A.P.; Liu, J.; Rogawski, E.T.; Kabir, F.; Lertsethtakarn, P.; Siguas, M.; Investigators, M.-E.N. Use of quantitative molecular diagnostic methods to assess the aetiology, burden, and clinical characteristics of diarrhoea in children in low-resource settings: A reanalysis of the MAL-ED cohort study. Lancet Glob. Health 2018, 6, e1309–e1318. [Google Scholar] [CrossRef] [Green Version]
- Graves, N.S. Acute gastroenteritis. Prim. Care 2013, 40, 727–741. [Google Scholar] [CrossRef] [PubMed]
- Eckardt, A.J.; Baumgart, D.C. Viral gastroenteritis in adults. Recent Pat. Antiinfect. Drug Discov. 2011, 6, 54–63. [Google Scholar] [CrossRef]
- Burnett, E.; Jonesteller, C.L.; Tate, J.E.; Yen, C.; Parashar, U.D. Global impact of rotavirus vaccination on childhood hospitalizations and mortality from diarrhea. J. Infect. Dis. 2017, 215, 1666–1672. [Google Scholar] [CrossRef]
- Vu, D.L.; Bosch, A.; Pinto, R.M.; Guix, S. Epidemiology of classic and novel human astrovirus: Gastroenteritis and beyond. Viruses 2017, 9, 33. [Google Scholar] [CrossRef]
- Aktas, O.; Aydin, H.; Timurkan, M.O. A molecular study on the prevalence and co-infections of rotavirus, norovirus, astrovirus and adenovirus in children with gastroenteritis. Minerva Pediatr. 2019, 71, 431–437. [Google Scholar] [CrossRef]
- Ahmed, S.M.; Hall, A.J.; Robinson, A.E.; Verhoef, L.; Premkumar, P.; Parashar, U.D.; Lopman, B.A. Global prevalence of norovirus in cases of gastroenteritis: A systematic review and meta-analysis. Lancet Infect. Dis. 2014, 14, 725–730. [Google Scholar] [CrossRef] [Green Version]
- Xing, W.; Liao, Q.; Viboud, C.; Zhang, J.; Sun, J.; Wu, J.T.; Yu, H. Hand, foot, and mouth disease in China, 2008–2012: An epidemiological study. Lancet Infect. Dis. 2014, 14, 308–318. [Google Scholar] [CrossRef] [Green Version]
- Jevsnik, M.; Steyer, A.; Zrim, T.; Pokorn, M.; Mrvic, T.; Grosek, S.; Petrovec, M. Detection of human coronaviruses in simultaneously collected stool samples and nasopharyngeal swabs from hospitalized children with acute gastroenteritis. Virol. J. 2013, 10, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, T.; Perlot, T.; Rehman, A.; Trichereau, J.; Ishiguro, H.; Paolino, M.; Penninger, J.M. ACE2 links amino acid malnutrition to microbial ecology and intestinal inflammation. Nature 2012, 487, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Burgueno, J.F.; Reich, A.; Hazime, H.; Quintero, M.A.; Fernandez, I.; Fritsch, J.; Abreu, M.T. Expression of SARS-CoV-2 entry molecules ACE2 and TMPRSS2 in the gut of patients with IBD. Inflamm. Bowel Dis. 2020, 26, 797–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holtz, L.R.; Cao, S.; Zhao, G.; Bauer, I.K.; Denno, D.M.; Klein, E.J.; Wang, D. Geographic variation in the eukaryotic virome of human diarrhea. Virology 2014, 468–470, 556–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foca, A.; Liberto, M.C.; Quirino, A.; Marascio, N.; Zicca, E.; Pavia, G. Gut inflammation and immunity: What is the role of the human gut virome? Mediat. Inflamm. 2015, 2015, 326032. [Google Scholar] [CrossRef] [PubMed]
- Neil, J.A.; Cadwell, K. The Intestinal virome and immunity. J. Immunol. 2018, 201, 1615–1624. [Google Scholar] [CrossRef] [Green Version]
- Reyes, A.; Haynes, M.; Hanson, N.; Angly, F.E.; Heath, A.C.; Rohwer, F.; Gordon, J.I. Viruses in the faecal microbiota of monozygotic twins and their mothers. Nature 2010, 466, 334–338. [Google Scholar] [CrossRef]
- Roux, S.; Hawley, A.K.; Beltran, M.T.; Scofield, M.; Schwientek, P.; Stepanauskas, R.; Woyke, T.; Hallam, S.J.; Sullivan, M.B. Ecology and evolution of viruses infecting uncultivated SUP05 bacteria as revealed by single-cell- and meta-genomics. eLife 2014, 3, e03125. [Google Scholar] [CrossRef]
- Labonte, J.M.; Swan, B.K.; Poulos, B.; Luo, H.; Koren, S.; Hallam, S.J.; Sullivan, M.B.; Woyke, T.; Wommack, E.C.; Stepanauskas, R. Single-cell genomics-based analysis of virus-host interactions in marine surface bacterioplankton. ISME J. 2015, 9, 2386–2399. [Google Scholar] [CrossRef]
- Munoz, S.L.D. Viral co-infection is shaped by host ecology and virus-virus interactions across diverse microbial taxa and environments. Virus Evol. 2017, 3. [Google Scholar] [CrossRef] [Green Version]
- Dzunkova, M.; Low, S.J.; Daly, J.N.; Deng, L.; Rinke, C.; Hugenholtz, P. Defining the human gut host-phage network through single-cell viral tagging. Nat. Microbiol. 2019, 4, 2192–2203. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Zhao, C.; Zhang, H.; Mattei, L.; Mix, S.S.; Bittinger, K.; Kessler, L.R.; Wu, G.D.; Baldassano, R.N.; DeRusso, P.; et al. The stepwise assembly of the neonatal virome is modulated by breastfeeding. Nature 2020, 581, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Maqsood, R.; Rodgers, R.; Rodriguez, C.; Handley, S.A.; Ndao, I.M.; Tarr, P.I.; Holtz, L.R. Discordant transmission of bacteria and viruses from mothers to babies at birth. Microbiome 2019, 7, 156. [Google Scholar] [CrossRef] [PubMed]
- Lim, E.S.; Zhou, Y.; Zhao, G.; Bauer, I.K.; Droit, L.; Ndao, I.M.; Holtz, L.R. Early life dynamics of the human gut virome and bacterial microbiome in infants. Nat. Med. 2015, 21, 1228–1234. [Google Scholar] [CrossRef]
- Pijnacker, R.; Pelt, W.V.; Vennema, H.; Kortbeek, L.M.; Notermans, D.W.; Franz, E.; Gras, L.M. Clinical relevance of enteropathogen co-infections in preschool children-a population-based repeated cross-sectional study. Clin. Microbiol. Infect. 2019, 25, 1013–1037. [Google Scholar] [CrossRef]
- Kapusinszky, B.; Minor, P.; Delwart, E. Nearly constant shedding of diverse enteric viruses by two healthy infants. J. Clin. Microbiol. 2012, 50, 3427–3434. [Google Scholar] [CrossRef] [Green Version]
- Tandukar, S.; Sherchand, J.B.; Karki, S.; Malla, B.; Shrestha, R.G.; Bhandari, D.; Haramoto, E. Co-infection by waterborne enteric viruses in children with gastroenteritis in nepal. Healthcare 2019, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Mengelle, C.; Mansuy, J.M.; Prere, M.F.; Grouteau, E.; Claudet, I.; Kamar, N.; Izopet, J. Simultaneous detection of gastrointestinal pathogens with a multiplex Luminex-based molecular assay in stool samples from diarrhoeic patients. Clin. Microbiol. Infect. 2013, 19, E458–E465. [Google Scholar] [CrossRef] [Green Version]
- Eibach, D.; Krumkamp, R.; Hahn, A.; Sarpong, N.; Sarkodie, Y.A.; Leva, A.; Tannich, E. Application of a multiplex PCR assay for the detection of gastrointestinal pathogens in a rural African setting. BMC Infect. Dis. 2016, 16, 150. [Google Scholar] [CrossRef] [Green Version]
- Mokomane, M.; Tate, J.E.; Steenhoff, A.P.; Esona, M.D.; Bowen, M.D.; Lechiile, K.; Goldfarb, D.M. Evaluation of the influence of gastrointestinal coinfections on rotavirus vaccine effectiveness in botswana. Pediatr. Infect. Dis. J. 2018, 37, e58–e62. [Google Scholar] [CrossRef] [Green Version]
- Mills, J.A.P.; Babji, S.; Bodhidatta, L.; Gratz, J.; Haque, R.; Havt, A.; Investigators, M.-E.N. Pathogen-specific burdens of community diarrhoea in developing countries: A multisite birth cohort study (MAL-ED). Lancet Glob. Health. 2015, 3, e564–e575. [Google Scholar] [CrossRef] [Green Version]
- Clark, A.; Black, R.; Tate, J.; Roose, A.; Kotloff, K.; Lam, D.; Steele, D. Estimating global, regional and national rotavirus deaths in children aged <5 years: Current approaches, new analyses and proposed improvements. PLoS ONE 2017, 12, e0183392. [Google Scholar] [CrossRef]
- Deen, J.; Lopez, A.L.; Kanungo, S.; Wang, X.Y.; Anh, D.D.; Tapia, M.; Grais, R.F. Improving rotavirus vaccine coverage: Can newer-generation and locally produced vaccines help? Hum. Vaccin. Immunother. 2018, 14, 495–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pindyck, T.; Tate, J.E.; Parashar, U.D. A decade of experience with rotavirus vaccination in the United States -vaccine uptake, effectiveness, and impact. Expert Rev. Vaccines 2018, 17, 593–606. [Google Scholar] [CrossRef] [PubMed]
- Ramig, R.F. Pathogenesis of intestinal and systemic rotavirus infection. J. Virol. 2004, 78, 10213–10220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McAtee, C.L.; Webman, R.; Gilman, R.H.; Mejia, C.; Bern, C.; Apaza, S.; Torrico, F. Burden of norovirus and rotavirus in children after rotavirus vaccine introduction, Cochabamba, Bolivia. Am. J. Trop Med. Hyg. 2016, 94, 212–217. [Google Scholar] [CrossRef] [Green Version]
- Hassan, E.; Baldridge, M.T. Norovirus encounters in the gut: Multifaceted interactions and disease outcomes. Mucosal Immunol. 2019, 12, 1259–1267. [Google Scholar] [CrossRef]
- Graaf, M.d.; Beek, J.V.; Koopmans, M.P. Human norovirus transmission and evolution in a changing world. Nat. Rev. Microbiol. 2016, 14, 421–433. [Google Scholar] [CrossRef]
- Zhang, S.X.; Zhou, Y.M.; Xu, W.; Tian, L.G.; Chen, J.X.; Chen, S.H.; Zhou, X.N. Impact of co-infections with enteric pathogens on children suffering from acute diarrhea in southwest China. Infect. Dis. Poverty 2016, 5, 64. [Google Scholar] [CrossRef] [Green Version]
- Green, K.Y.; Kaufman, S.S.; Nagata, B.M.; Chaimongkol, N.; Kim, D.Y.; Levenson, E.A.; Sosnovtsev, S.V. Human norovirus targets enteroendocrine epithelial cells in the small intestine. Nat. Commun. 2020, 11, 2759. [Google Scholar] [CrossRef]
- Karandikar, U.C.; Crawford, S.E.; Ajami, N.J.; Murakami, K.; Kou, B.; Ettayebi, K.; Estes, M.K. Detection of human norovirus in intestinal biopsies from immunocompromised transplant patients. J. Gen. Virol. 2016, 97, 2291–2300. [Google Scholar] [CrossRef] [PubMed]
- Dreps, S.B.; Bucardo, F.; Vinje, J. Sapovirus: An important cause of acute gastroenteritis in children. Lancet Child. Adolesc. Health 2019, 3, 758–759. [Google Scholar] [CrossRef]
- Varela, M.F.; Rivadulla, E.; Lema, A.; Romalde, J.L. Human sapovirus among outpatients with acute gastroenteritis in spain: A one-year study. Viruses 2019, 11, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roels, G.L.; Cramer, J.P.; Mendelman, P.M.; Sherwood, J.; Clemens, R.; Aerssens, A.; Damme, P.V. Safety and immunogenicity of different formulations of norovirus vaccine candidate in healthy adults: A randomized, controlled, double-blind clinical trial. J. Infect. Dis. 2018, 217, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Hallowell, B.D.; Parashar, U.D.; Hall, A.J. Epidemiologic challenges in norovirus vaccine development. Hum. Vaccin Immunother. 2019, 15, 1279–1283. [Google Scholar] [CrossRef] [PubMed]
- Koopmans, M.P.; Bijen, M.H.; Monroe, S.S.; Vinje, J. Age-stratified seroprevalence of neutralizing antibodies to astrovirus types 1 to 7 in humans in The Netherlands. Clin. Diagn. Lab. Immunol. 1998, 5, 33–37. [Google Scholar] [CrossRef] [Green Version]
- Kriston, S.; Willcocks, M.M.; Carter, M.J.; Cubitt, W.D. Seroprevalence of astrovirus types 1 and 6 in London, determined using recombinant virus antigen. Epidemiol. Infect. 1996, 117, 159–164. [Google Scholar] [CrossRef] [Green Version]
- Wunderli, W.; Meerbach, A.; Gungor, T.; Berger, C.; Greiner, O.; Caduff, R.; Tapparel, C. Astrovirus infection in hospitalized infants with severe combined immunodeficiency after allogeneic hematopoietic stem cell transplantation. PLoS ONE 2011, 6, e27483. [Google Scholar] [CrossRef] [Green Version]
- Quan, P.L.; Wagner, T.A.; Briese, T.; Torgerson, T.R.; Hornig, M.; Tashmukhamedova, A.; Lipkin, W.I. Astrovirus encephalitis in boy with X-linked agammaglobulinemia. Emerg. Infect. Dis. 2010, 16, 918–925. [Google Scholar] [CrossRef]
- Brown, J.R.; Morfopoulou, S.; Hubb, J.; Emmett, W.A.; Ip, W.; Shah, D.; Breuer, J. Astrovirus VA1/HMO-C: An increasingly recognized neurotropic pathogen in immunocompromised patients. Clin. Infect. Dis. 2015, 60, 881–888. [Google Scholar] [CrossRef]
- Cordey, S.; Vu, D.L.; Schibler, M.; L’Huillier, A.G.; Brito, F.; Docquier, M.; Kaiser, L. Astrovirus MLB2, a new gastroenteric virus associated with meningitis and disseminated infection. Emerg. Infect. Dis. 2016, 22, 846–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, M.; Kuroda, M.; Kasai, M.; Matsui, H.; Fukuyama, T.; Katano, H.; Taya, K.T. Acute encephalopathy in an immunocompromised boy with astrovirus-MLB1 infection detected by next generation sequencing. J. Clin. Virol. 2016, 78, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Kolawole, A.O.; Mirabelli, C.; Hill, D.R.; Svoboda, S.A.; Janowski, A.B.; Passalacqua, K.D.; Wobus, C.E. Astrovirus replication in human intestinal enteroids reveals multi-cellular tropism and an intricate host innate immune landscape. PLoS Pathog. 2019, 15, e1008057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehndiratta, M.M.; Mehndiratta, P.; Pande, R. Poliomyelitis: Historical facts, epidemiology, and current challenges in eradication. Neurohospitalist 2014, 4, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Lancaster, K.Z.; Pfeiffer, J.K. Limited trafficking of a neurotropic virus through inefficient retrograde axonal transport and the type I interferon response. PLoS Pathog. 2010, 6, e1000791. [Google Scholar] [CrossRef] [PubMed]
- Mwengee, W.; Okeibunor, J.; Poy, A.; Shaba, K.; Kinuani, L.M.; Minkoulou, E.; Mkanda, P. Polio eradication initiative: Contribution to improved communicable diseases surveillance in WHO African region. Vaccine 2016, 34, 5170–5174. [Google Scholar] [CrossRef] [Green Version]
- Salk, J.E. Considerations in the preparation and use of poliomyelitis virus vaccine. J. Am. Med. Assoc. 1955, 158, 1239–1248. [Google Scholar] [CrossRef]
- Krugman, S.; Warren, J.; Eiger, M.S.; Berman, P.H.; Michaels, R.M.; Sabin, A.B. Immunization with live attenuated poliovirus vaccine. Am. J. Dis. Child. 1961, 101, 23–29. [Google Scholar] [CrossRef]
- Shin, W.J.; Hara, D.; Gbormittah, F.; Chang, H.; Chang, B.S.; Jung, J.U. Development of thermostable lyophilized sabin inactivated poliovirus vaccine. mBio 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Platt, L.R.; Estivariz, C.F.; Sutter, R.W. Vaccine-associated paralytic poliomyelitis: A review of the epidemiology and estimation of the global burden. J. Infect. Dis. 2014, 210, S380–S389. [Google Scholar] [CrossRef]
- Jorba, J.; Diop, O.M.; Iber, J.; Henderson, E.; Zhao, K.; Quddus, A.; Burns, C.C. Update on vaccine-derived poliovirus outbreaks-worldwide, January 2018–June 2019. MMWR Morb. Mortal. Wkly. Rep. 2019, 68, 1024–1028. [Google Scholar] [CrossRef] [PubMed]
- Koh, W.M.; Bogich, T.; Siegel, K.; Jin, J.; Chong, E.Y.; Tan, C.Y.; Cook, A.R. The epidemiology of hand, foot and mouth disease in Asia: A systematic review and analysis. Pediatr. Infect. Dis. J. 2016, 35, e285–e300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Wu, S.; Xiong, Y.; Li, T.; Wen, Z.; Yan, M.; Wu, J. Co-circulation and genomic recombination of coxsackievirus A16 and enterovirus 71 during a large outbreak of hand, foot, and mouth disease in Central China. PLoS ONE 2014, 9, e96051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.F.; Zhang, Y.; Hou, P.Q.; Zhu, S.Y.; Wu, X.Y.; Zhao, H.; Qin, C.F. Human enterovirus co-infection in severe HFMD patients in China. J. Clin. Virol. 2014, 61, 621–622. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Du, Z.; Feng, M.; Che, Y.; Li, Q. A severe case of co-infection with Enterovirus 71 and vaccine-derived Poliovirus type II. J. Clin. Virol. 2015, 72, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Patil, P.R.; Chitambar, S.D.; Gopalkrishna, V. Molecular surveillance of non-polio enterovirus infections in patients with acute gastroenteritis in Western India: 2004–2009. J. Med. Virol. 2015, 87, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Chansaenroj, J.; Tuanthap, S.; Thanusuwannasak, T.; Duang, A.I.; Klinfueng, S.; Thaneskongtong, N.; Poovorawan, Y. Human enteroviruses associated with and without diarrhea in Thailand between 2010 and 2016. PLoS ONE 2017, 12, e0182078. [Google Scholar] [CrossRef] [Green Version]
- Kumthip, K.; Khamrin, P.; Ushijima, H.; Maneekarn, N. Multiple enterovirus genotypes circulating in children hospitalized with acute gastroenteritis in Thailand. Infect. Genet. Evol. 2017, 55, 324–331. [Google Scholar] [CrossRef]
- Lynch, J.P.; Fishbein, M.; Echavarria, M. Adenovirus. Semin. Respir. Crit. Care Med. 2011, 32, 494–511. [Google Scholar] [CrossRef]
- Qiu, F.Z.; Shen, X.X.; Li, G.X.; Zhao, L.; Chen, C.; Duan, S.X.; Ma, X.J. Adenovirus associated with acute diarrhea: A case-control study. BMC Infect. Dis. 2018, 18, 450. [Google Scholar] [CrossRef]
- Kumthip, K.; Khamrin, P.; Ushijima, H.; Maneekarn, N. Enteric and non-enteric adenoviruses associated with acute gastroenteritis in pediatric patients in Thailand, 2011 to 2017. PLoS ONE 2019, 14, e0220263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, A.D.P.; Manna, B.; Sarkar, M.C. Molecular characterization of enteric adenovirus genotypes 40 and 41 identified in children with acute gastroenteritis in Kolkata, India during 2013–2014. J. Med. Virol. 2017, 89, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Uhnoo, I.; Wadell, G.; Svensson, L.; Johansson, M.E. Importance of enteric adenoviruses 40 and 41 in acute gastroenteritis in infants and young children. J. Clin. Microbiol. 1984, 20, 365–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filho, E.P.; Faria, N.R.d.C.; Fialho, A.M.; Assis, R.S.d.; Almeida, M.M.S.; Rocha, M.; Leite, J.P.G. Adenoviruses associated with acute gastroenteritis in hospitalized and community children up to 5 years old in Rio de Janeiro and Salvador, Brazil. J. Med. Microbiol. 2007, 56, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Kosulin, K. Intestinal HAdV infection: Tissue specificity, persistence, and implications for antiviral therapy. Viruses 2019, 11, 804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lion, T. Adenovirus infections in immunocompetent and immunocompromised patients. Clin. Microbiol. Rev. 2014, 27, 441–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Echavarria, M. Adenoviruses in immunocompromised hosts. Clin. Microbiol. Rev. 2008, 21, 704–715. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.Y.; Luo, Y.P.; Huang, D.D.; Fan, H.; Lu, Q.B.; Wo, Y.; Liu, W. Fatal pneumonia cases caused by human adenovirus 55 in immunocompetent adults. Infect. Dis. 2016, 48, 40–47. [Google Scholar] [CrossRef]
- Thomas, P.D.; Pollok, R.C.; Gazzard, B.G. Enteric viral infections as a cause of diarrhoea in the acquired immunodeficiency syndrome. HIV Med. 1999, 1, 19–24. [Google Scholar] [CrossRef]
- Roy, S.; Calcedo, R.; Jaszek, A.M.; Keough, M.; Peng, H.; Wilson, J.M. Adenoviruses in lymphocytes of the human gastro-intestinal tract. PLoS ONE 2011, 6, e24859. [Google Scholar] [CrossRef]
- Holshue, M.L.; DeBolt, C.; Lindquist, S.; Lofy, K.H.; Wiesman, J.; Bruce, H. First case of 2019 novel coronavirus in the United States. N. Engl. J. Med. 2020, 382, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Tang, M.; Zheng, X.; Liu, Y.; Li, X.; Shan, H. Evidence for gastrointestinal infection of SARS-CoV-2. Gastroenterology 2020, 158, 1831–1833. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Li, X.; Zhu, B.; Liang, H.; Fang, C.; Gong, Y.; Gong, S. Characteristics of pediatric SARS-CoV-2 infection and potential evidence for persistent fecal viral shedding. Nat. Med. 2020, 26, 502–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munster, V.J.; Feldmann, F.; Williamson, B.N.; Doremalen, N.V.; Perez, L.P.; Schulz, J.; Wit, E.d. Respiratory disease in rhesus macaques inoculated with SARS-CoV-2. Nature 2020. [Google Scholar] [CrossRef]
- Kim, Y.I.; Kim, S.G.; Kim, S.M.; Kim, E.H.; Park, S.J.; Yu, K.M.; Choi, Y.K. Infection and rapid transmission of SARS-CoV-2 in Ferrets. Cell Host Microbe 2020, 27, 704–709. [Google Scholar] [CrossRef]
- Lu, X.; Zhang, L.; Du, H.; Zhang, J.; Li, Y.Y.; Qu, J.; Chinese pediatric novel coronavirus study, T. SARS-CoV-2 infection in children. N. Engl. J. Med. 2020, 382, 1663–1665. [Google Scholar] [CrossRef] [Green Version]
- Nowak, M.D.; Sordillo, E.M.; Gitman, M.R.; Paniz Mondolfi, A.E. Co-infection in SARS-CoV-2 infected patients: Where are influenza virus and rhinovirus/enterovirus? J. Med. Virol. 2020. [Google Scholar] [CrossRef]
- Zang, R.; Castro, M.F.G.; McCune, B.T.; Zeng, Q.; Rothlauf, P.W.; Sonnek, N.M.; Ding, S. TMPRSS2 and TMPRSS4 promote SARS-CoV-2 infection of human small intestinal enterocytes. Sci. Immunol. 2020, 5. [Google Scholar] [CrossRef]
- Xiao, F.; Sun, J.; Xu, Y.; Li, F.; Huang, X.; Li, H.; Zhao, J. Infectious SARS-CoV-2 in feces of patient with severe COVID-19. Emerg. Infect. Dis. 2020, 26. [Google Scholar] [CrossRef]
- Ruggeri, F.M.; Delogu, R.; Petouchoff, T.; Tcheremenskaia, O.; Petris, S.D.; Fiore, L. RotaNet-Italy study group molecular characterization of rotavirus strains from children with diarrhea in Italy, 2007–2009. J. Med. Virol. 2011, 83, 1657–1668. [Google Scholar] [CrossRef] [Green Version]
- Eiden, J.; Losonsky, G.A.; Johnson, J.; Yolken, R.H. Rotavirus RNA variation during chronic infection of immunocompromised children. Pediatr. Infect. Dis. 1985, 4, 632–637. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Apaza, S.G.; Espetia, S.; Velasquez, D.; Cabrera, L.; Loli, S. Multiple norovirus infections in a birth cohort in a Peruvian Periurban community. Clin. Infect. Dis. 2014, 58, 483–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruber, J.F.; Bowman, N.M.; Dreps, S.B.; Reyes, Y.; Belson, C.; Michaels, K.C.; Bucardo, F. Risk factors for norovirus gastroenteritis among nicaraguan children. Am. J. Trop. Med. Hyg. 2017, 97, 937–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menon, V.K.; George, S.; Sarkar, R.; Giri, S.; Samuel, P.; Vivek, R.; Kang, G. Norovirus gastroenteritis in a birth cohort in Southern India. PLoS ONE 2016, 11, e0157007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, D.C.; Babu, M.A.; Raghavendra, A.; Dhananjaya, D.; Kumar, S.; Maiya, P.P. Non-polio enteroviruses and their association with acute diarrhea in children in India. Infect. Genet. Evol. 2013, 17, 153–161. [Google Scholar] [CrossRef]
- Wu, D.; Changwen, K.; Wei, L.; Monagin, C.; Jin, Y.; Cong, M.; Jun, S. A large outbreak of hand, foot, and mouth disease caused by EV71 and CAV16 in Guangdong, China, 2009. Arch. Virol. 2011, 156, 945–953. [Google Scholar] [CrossRef]
- Rasanen, S.; Lappalainen, S.; Kaikkonen, S.; Hamalainen, M.; Salminen, M.; Vesikari, T. Mixed viral infections causing acute gastroenteritis in children in a waterborne outbreak. Epidemiol. Infect. 2010, 138, 1227–1234. [Google Scholar] [CrossRef]
- Chitambar, S.; Gopalkrishna, V.; Chhabra, P.; Patil, P.; Verma, H.; Lahon, A.; Pawar, S. Diversity in the enteric viruses detected in outbreaks of gastroenteritis from Mumbai, Western India. Int. J. Environ. Res. Public Health 2012, 9, 895–915. [Google Scholar] [CrossRef]
- Roman, E.; Wilhelmi, I.; Colomina, J.; Villar, J.; Cilleruelo, M.L.; Nebreda, V.; Fauquier, A.S. Acute viral gastroenteritis: Proportion and clinical relevance of multiple infections in Spanish children. J. Med. Microbiol. 2001, 52, 435–440. [Google Scholar] [CrossRef] [Green Version]
- Bon, F.; Fascia, P.; Dauvergne, M.; Tenenbaum, D.; Planson, H.; Petion, A.M.; Kohli, E. Prevalence of group a rotavirus, human calicivirus, astrovirus, and adenovirus type 40 and 41 infections among children with acute gastroenteritis in Dijon, France. J. Clin. Microbiol. 1999, 37, 3055–3058. [Google Scholar] [CrossRef] [Green Version]
- Leblanc, D.; Inglis, G.D.; Boras, V.F.; Brassard, J.; Houde, A. The prevalence of enteric RNA viruses in stools from diarrheic and non-diarrheic people in southwestern Alberta, Canada. Arch. Virol. 2017, 162, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Vu, D.L.; Sabria, A.; Aregall, N.; Michl, K.; Garrido, V.R.; Goterris, L.; Guix, S. Novel human astroviruses: Prevalence and association with common enteric viruses in undiagnosed gastroenteritis cases in Spain. Viruses 2019, 11, 585. [Google Scholar] [CrossRef] [Green Version]
- Wayman, S.D.; Fahle, G.; Palmore, T.; Green, K.Y.; Prevots, D.R. Norovirus, astrovirus, and sapovirus among immunocompromised patients at a tertiary care research hospital. Diagn. Microbiol. Infect. Dis. 2018, 92, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.R.; Shah, D.; Breuer, J. Viral gastrointestinal infections and norovirus genotypes in a paediatric UK hospital, 2014–2015. J. Clin. Virol. 2016, 84, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Green, K.Y. Norovirus infection in immunocompromised hosts. Clin. Microbiol. Infect. 2014, 20, 717–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beek, J.V.; Graaf, M.d.; Smits, S.; Schapendonk, C.M.E.; Verjans, G.; Vennema, H.; Koopmans, M. Whole-genome next-generation sequencing to study within-host evolution of norovirus (NoV) among immunocompromised patients with chronic NoV infection. J. Infect. Dis. 2017, 216, 1513–1524. [Google Scholar] [CrossRef] [PubMed]
- Munir, N.; Liu, P.; Gastanaduy, P.; Montes, J.; Shane, A.; Moe, C. Norovirus infection in immunocompromised children and children with hospital-acquired acute gastroenteritis. J. Med. Virol. 2014, 86, 1203–1209. [Google Scholar] [CrossRef]
- Chong, P.P.; Atmar, R.L. Norovirus in health care and implications for the immunocompromised host. Curr. Opin. Infect. Dis. 2019, 32, 348–355. [Google Scholar] [CrossRef]
- Liste, M.B.; Natera, I.; Suarez, J.A.; Pujol, F.H.; Liprandi, F.; Ludert, J.E. Enteric virus infections and diarrhea in healthy and human immunodeficiency virus-infected children. J. Clin. Microbiol. 2000, 38, 2873–2877. [Google Scholar] [CrossRef] [Green Version]
- Portes, S.A.R.; Costa, F.A.C.; Rocha, M.S.; Fumian, T.M.; Maranhao, A.G.; Assis, R.M.d.; Volotao, E.M. Enteric viruses in HIV-1 seropositive and HIV-1 seronegative children with diarrheal diseases in Brazil. PLoS ONE 2017, 12, e0183196. [Google Scholar] [CrossRef] [Green Version]
- Lewandowska, D.W.; Capaul, R.; Prader, S.; Zagordi, O.; Geissberger, F.D.; Kugler, M.; Huber, M. Persistent mammalian orthoreovirus, coxsackievirus and adenovirus co-infection in a child with a primary immunodeficiency detected by metagenomic sequencing: A case report. BMC Infect. Dis. 2018, 18, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, H.C.; Borges, F.P.; Arantes, A.M.; Souza, M. Detection of human adenovirus (species-C, -D and -F) in an allogeneic stem cell transplantation recipient: A case report. Rev. Bras. Hematol. Hemoter. 2017, 39, 60–62. [Google Scholar] [CrossRef] [Green Version]
- Bodhidatta, L.; McDaniel, P.; Sornsakrin, S.; Srijan, A.; Serichantalergs, O.; Mason, C.J. Case-control study of diarrheal disease etiology in a remote rural area in Western Thailand. Am. J. Trop Med. Hyg. 2010, 83, 1106–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Ryan, M.L.; Lucero, Y.; Prado, V.; Santolaya, M.E.; Rabello, M.; Solis, Y.; Mamani, N. Symptomatic and asymptomatic rotavirus and norovirus infections during infancy in a Chilean birth cohort. Pediatr. Infect. Dis. J. 2009, 28, 879–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okitsu, S.; Khamrin, P.; Takanashi, S.; Thongprachum, A.; Hoque, S.A.; Takeuchi, H.; Ushijima, H. Molecular detection of enteric viruses in the stool samples of children without diarrhea in Bangladesh. Infect. Genet. Evol. 2020, 77, 104055. [Google Scholar] [CrossRef] [PubMed]
- Ayukekbong, J.; Lindh, M.; Nenonen, N.; Tah, F.; Akenji, T.N.; Bergstrom, T. Enteric viruses in healthy children in Cameroon: Viral load and genotyping of norovirus strains. J. Med. Virol. 2011, 83, 2135–2142. [Google Scholar] [CrossRef]
- Akihara, S.; Phan, T.G.; Nguyen, T.A.; Hansman, G.; Okitsu, S.; Ushijima, H. Existence of multiple outbreaks of viral gastroenteritis among infants in a day care center in Japan. Arch. Virol. 2005, 150, 2061–2075. [Google Scholar] [CrossRef]
- Nathanson, N.; Kew, O.M. From emergence to eradication: The epidemiology of poliomyelitis deconstructed. Am. J. Epidemiol. 2010, 172, 1213–1229. [Google Scholar] [CrossRef]
- Ramani, S.; Harris, V. Going viral! Unraveling the impact of nonpolio enteroviruses on oral vaccine responses. J. Infect. Dis. 2019, 219, 1173–1175. [Google Scholar] [CrossRef]
- Parker, E.P.; Kampmann, B.; Kang, G.; Grassly, N.C. Influence of enteric infections on response to oral poliovirus vaccine: A systematic review and meta-analysis. J. Infect. Dis. 2014, 210, 853–864. [Google Scholar] [CrossRef] [Green Version]
- Praharaj, I.; Parker, E.P.K.; Giri, S.; Allen, D.J.; Silas, S.; Revathi, R.; Kang, G. Influence of nonpolio enteroviruses and the bacterial gut microbiota on oral poliovirus vaccine response: A study from South India. J. Infect. Dis. 2019, 219, 1178–1186. [Google Scholar] [CrossRef] [PubMed]
- Taniuchi, M.; Mills, J.A.P.; Begum, S.; Uddin, M.J.; Sobuz, S.U.; Liu, J.; Houpt, E.R. Impact of enterovirus and other enteric pathogens on oral polio and rotavirus vaccine performance in Bangladeshi infants. Vaccine 2016, 34, 3068–3075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, E.P.K.; Praharaj, I.; Zekavati, A.; Lazarus, R.P.; Giri, S.; Operario, D.J.; Grassly, N.C. Influence of the intestinal microbiota on the immunogenicity of oral rotavirus vaccine given to infants in south India. Vaccine 2018, 36, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.; Steele, A.D.; Parashar, U.D. Influence of oral polio vaccines on performance of the monovalent and pentavalent rotavirus vaccines. Vaccine 2012, A30–A35. [Google Scholar] [CrossRef] [PubMed]
- Groome, M.J.; Page, N.; Cortese, M.M.; Moyes, J.; Zar, H.J.; Kapongo, C.N.; Madhi, S.A. Effectiveness of monovalent human rotavirus vaccine against admission to hospital for acute rotavirus diarrhoea in South African children: A case-control study. Lancet Infect. Dis. 2014, 14, 1096–1104. [Google Scholar] [CrossRef]
- Praharaj, I.; Mills, J.A.P.; Taneja, S.; Antony, K.; Yuhas, K.; Flores, J.; Kang, G. Diarrheal etiology and impact of coinfections on rotavirus vaccine efficacy estimates in a clinical trial of a monovalent human-bovine (116E) oral rotavirus vaccine, Rotavac, India. Clin. Infect. Dis. 2019, 69, 243–250. [Google Scholar] [CrossRef] [Green Version]
- Brouwer, L.; Benschop, K.S.M.; Nguyen, D.; Kamau, E.; Pajkrt, D.; Simmonds, P.; Wolthers, K.C. Recombination analysis of non-poliovirus members of the enterovirus C species; restriction of recombination events to members of the same 3DPol cluster. Viruses 2020, 12, 706. [Google Scholar] [CrossRef]
- Muslin, C.; Kain, A.M.; Bessaud, M.; Blondel, B.; Delpeyroux, F. Recombination in enteroviruses, a multi-step modular evolutionary process. Viruses 2019, 11, 859. [Google Scholar] [CrossRef] [Green Version]
- Bessaud, M.; Joffret, M.L.; Blondel, B.; Delpeyroux, F. Exchanges of genomic domains between poliovirus and other cocirculating species C enteroviruses reveal a high degree of plasticity. Sci. Rep. 2016, 6, 38831. [Google Scholar] [CrossRef]
- Wohlgemuth, N.; Honce, R.; Cherry, S.S. Astrovirus evolution and emergence. Infect. Genet. Evol. 2019, 69, 30–37. [Google Scholar] [CrossRef]
- Begall, L.F.L.; Mauroy, A.; Thiry, E. Norovirus recombinants: Recurrent in the field, recalcitrant in the lab-a scoping review of recombination and recombinant types of noroviruses. J. Gen. Virol. 2018, 99, 970–988. [Google Scholar] [CrossRef] [PubMed]
- McDonald, S.M.; Nelson, M.I.; Turner, P.E.; Patton, J.T. Reassortment in segmented RNA viruses: Mechanisms and outcomes. Nat. Rev. Microbiol. 2016, 14, 448–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Liu, W.J.; Xu, W.; Jin, T.; Zhao, Y.; Song, J.; Gao, G.F. A bat-derived putative cross-family recombinant coronavirus with a reovirus gene. PLoS Pathog. 2016, 12, e1005883. [Google Scholar] [CrossRef] [PubMed]
- Santiana, M.; Ghosh, S.; Ho, B.A.; Rajasekaran, V.; Du, W.L.; Mutsafi, Y.; Bonnet, N.A. Vesicle-cloaked virus clusters are optimal units for inter-organismal viral transmission. Cell Host Microbe 2018, 24, 208–220. [Google Scholar] [CrossRef] [Green Version]
- Aguilera, E.R.; Erickson, A.K.; Jesudhasan, P.R.; Robinson, C.M.; Pfeiffer, J.K. Plaques formed by mutagenized viral populations have elevated co-infection frequencies. mBio 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Aguilera, E.R.; Pfeiffer, J.K. Strength in numbers: Mechanisms of viral co-infection. Virus Res. 2019, 265, 43–46. [Google Scholar] [CrossRef]
- Moshe, A.; Gorovits, R. Virus-induced aggregates in infected cells. Viruses 2012, 4, 2218–2232. [Google Scholar] [CrossRef] [Green Version]
- Moreno, I.A.; Sanjuan, R. Collective infection of cells by viral aggregates promotes early viral proliferation and reveals a cellular-level allee effect. Curr. Biol. 2018, 28, 3212–3219.e3214. [Google Scholar] [CrossRef] [Green Version]
- Erickson, A.K.; Jesudhasan, P.R.; Mayer, M.J.; Narbad, A.; Winter, S.E.; Pfeiffer, J.K. Bacteria facilitate enteric virus co-infection of mammalian cells and promote genetic recombination. Cell Host Microbe 2018, 23, 77–88. [Google Scholar] [CrossRef] [Green Version]
- Dianzani, F. Viral interference and interferon. Ric. Clin. Lab. 1975, 5, 196–213. [Google Scholar] [CrossRef]
- Ingle, H.; Lee, S.; Ai, T.; Orvedahl, A.; Rodgers, R.; Zhao, G.; Baldridge, M.T. Viral complementation of immunodeficiency confers protection against enteric pathogens via interferon-lambda. Nat. Microbiol. 2019, 4, 1120–1128. [Google Scholar] [CrossRef] [PubMed]
- Sestak, K. Non-human primate models of enteric viral infections. Viruses 2018, 10, 544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chege, G.K.; Steele, A.D.; Hart, C.A.; Snodgrass, D.R.; Omolo, E.O.; Mwenda, J.M. Experimental infection of non-human primates with a human rotavirus isolate. Vaccine 2005, 23, 1522–1528. [Google Scholar] [CrossRef] [PubMed]
- Bentes, G.A.; Guimaraes, J.R.; Volotao, E.M.; Fialho, A.M.; Hooper, C.; Ganime, A.C.; Pinto, M.A. Cynomolgus monkeys (Macaca fascicularis) as an experimental infection model for human group a rotavirus. Viruses 2018, 10, 355. [Google Scholar] [CrossRef] [Green Version]
- Rockx, B.H.; Bogers, W.M.; Heeney, J.L.; Amerongen, G.V.; Koopmans, M.P. Experimental norovirus infections in non-human primates. J. Med. Virol. 2005, 75, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Farkas, T.; Yang, K.; Pendu, J.L.; Baines, J.D.; Cardin, R.D. The coxsackievirus and adenovirus receptor, a required host factor for recovirus infection, is a putative enteric calicivirus receptor. J. Virol. 2019, 93. [Google Scholar] [CrossRef]
- Sestak, K.; McNeal, M.M.; Choi, A.; Cole, M.J.; Ramesh, G.; Alvarez, X.; Ward, R.L. Defining T-cell-mediated immune responses in rotavirus-infected juvenile rhesus macaques. J. Virol. 2004, 78, 10258–10264. [Google Scholar] [CrossRef] [Green Version]
- McNeal, M.M.; Sestak, K.; Choi, A.H.; Basu, M.; Cole, M.J.; Aye, P.P.; Ward, R.L. Development of a rotavirus-shedding model in rhesus macaques, using a homologous wild-type rotavirus of a new P genotype. J. Virol. 2005, 79, 944–954. [Google Scholar] [CrossRef] [Green Version]
- Hoshino, Y.; Honma, S.; Jones, R.W.; Santos, N.; Nakagomi, O.; Nakagomi, T.; Thouless, M.E. A rotavirus strain isolated from pig-tailed macaque (Macaca nemestrina) with diarrhea bears a P6[1]: G8 specificity. Virology 2006, 345, 1–12. [Google Scholar] [CrossRef] [Green Version]
- D’Arc, M.; Furtado, C.; Siqueira, J.D.; Seuanez, H.N.; Ayouba, A.; Peeters, M.; Soares, M.A. Assessment of the gorilla gut virome in association with natural simian immunodeficiency virus infection. Retrovirology 2018, 15, 19. [Google Scholar] [CrossRef]
- Handley, S.A.; Thackray, L.B.; Zhao, G.; Presti, R.; Miller, A.D.; Droit, L.; Virgin, H.W. Pathogenic simian immunodeficiency virus infection is associated with expansion of the enteric virome. Cell 2012, 151, 253–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handley, S.A.; Desai, C.; Zhao, G.; Droit, L.; Monaco, C.L.; Schroeder, A.C.; Virgin, H.W. SIV infection-mediated changes in gastrointestinal bacterial microbiome and virome are associated with immunodeficiency and prevented by vaccination. Cell Host Microbe 2016, 19, 323–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saif, L.J.; Ward, L.A.; Yuan, L.; Rosen, B.I.; To, T.L. The gnotobiotic piglet as a model for studies of disease pathogenesis and immunity to human rotaviruses. Arch. Virol. Suppl. 1996, 12, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, A.; Mao, J.; Lei, S.; Twitchell, E.; Shiraz, A.; Jiang, X.; Yuan, A.L. Parenterally administered P24-VP8* nanoparticle vaccine conferred strong protection against rotavirus diarrhea and virus shedding in gnotobiotic pigs. Vaccines 2019, 7, 177. [Google Scholar] [CrossRef] [Green Version]
- Yuan, L.; Ward, L.A.; Rosen, B.I.; To, T.L.; Saif, L.J. Systematic and intestinal antibody-secreting cell responses and correlates of protective immunity to human rotavirus in a gnotobiotic pig model of disease. J. Virol. 1996, 70, 3075–3083. [Google Scholar] [CrossRef] [Green Version]
- Troeger, H.; Loddenkemper, C.; Schneider, T.; Schreier, E.; Epple, H.J.; Zeitz, M.; Schulzke, J.D. Structural and functional changes of the duodenum in human norovirus infection. Gut 2009, 58, 1070–1077. [Google Scholar] [CrossRef]
- Sugieda, M.; Nakajima, S. Viruses detected in the caecum contents of healthy pigs representing a new genetic cluster in genogroup II of the genus “Norwalk-like viruses”. Virus Res. 2002, 87, 165–172. [Google Scholar] [CrossRef]
- Poel, W.H.v.D.; Vinje, J.; Heide, R.v.D.; Herrera, M.I.; Vivo, A.; Koopmans, M.P. Norwalk-like calicivirus genes in farm animals. Emerg. Infect. Dis. 2000, 6, 36–41. [Google Scholar] [CrossRef]
- Wang, Q.H.; Han, M.G.; Cheetham, S.; Souza, M.; Funk, J.A.; Saif, L.J. Porcine noroviruses related to human noroviruses. Emerg. Infect. Dis. 2005, 11, 1874–1881. [Google Scholar] [CrossRef]
- Koonpaew, S.; Teeravechyan, S.; Frantz, P.N.; Chailangkarn, T.; Jongkaewwattana, A. PEDV and PDCoV pathogenesis: The interplay between host innate immune responses and porcine enteric coronaviruses. Front. Vet. Sci. 2019, 6, 34. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.P.; Yang, Z.; Lin, W.D.; Wang, W.Y.; Yang, J.; Jin, W.J.; Qin, A.J. The rate of co-infection for piglet diarrhea viruses in China and the genetic characterization of porcine epidemic diarrhea virus and porcine kobuvirus. Acta Virol. 2016, 60, 55–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, S.H.; Che, X.; Garcia, J.A.; Klena, J.D.; Lee, B.; Muller, D.; Lipkin, W.I. Viral diversity of house mice in New York City. mBio 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortez, V.; Sharp, B.; Yao, J.; Livingston, B.; Vogel, P.; Cherry, S.S. Characterizing a murine model for astrovirus using viral isolates from persistently infected immunocompromised mice. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taube, S.; Kolawole, A.O.; Hohne, M.; Wilkinson, J.E.; Handley, S.A.; Perry, J.W.; Wobus, C.E. A mouse model for human norovirus. mBio 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Karst, S.M.; Wobus, C.E.; Lay, M.; Davidson, J.; Virgin, H.W. STAT1-dependent innate immunity to a Norwalk-like virus. Science 2003, 299, 1575–1578. [Google Scholar] [CrossRef]
- Baldridge, M.T.; Nice, T.J.; McCune, B.T.; Yokoyama, C.C.; Kambal, A.; Wheadon, M.; Virgin, H.W. Commensal microbes and interferon-lambda determine persistence of enteric murine norovirus infection. Science 2015, 347, 266–269. [Google Scholar] [CrossRef] [Green Version]
- Orchard, R.C.; Wilen, C.B.; Doench, J.G.; Baldridge, M.T.; McCune, B.T.; Lee, Y.C.; Virgin, H.W. Discovery of a proteinaceous cellular receptor for a norovirus. Science 2016, 353, 933–936. [Google Scholar] [CrossRef] [Green Version]
- Nice, T.J.; Baldridge, M.T.; McCune, B.T.; Norman, J.M.; Lazear, H.M.; Artyomov, M.; Virgin, H.W. Interferon-lambda cures persistent murine norovirus infection in the absence of adaptive immunity. Science 2015, 347, 269–273. [Google Scholar] [CrossRef] [Green Version]
- Wilen, C.B.; Lee, S.; Hsieh, L.L.; Orchard, R.C.; Desai, C.; Hykes, B.L.J.; Virgin, H.W. Tropism for tuft cells determines immune promotion of norovirus pathogenesis. Science 2018, 360, 204–208. [Google Scholar] [CrossRef] [Green Version]
- Roth, A.N.; Helm, E.W.; Mirabelli, C.; Kirsche, E.; Smith, J.C.; Eurell, L.B.; Karst, S.M. Norovirus infection causes acute self-resolving diarrhea in wild-type neonatal mice. Nat. Commun. 2020, 11, 2968. [Google Scholar] [CrossRef]
- Broome, R.L.; Vo, P.T.; Ward, R.L.; Clark, H.F.; Greenberg, H.B. Murine rotavirus genes encoding outer capsid proteins VP4 and VP7 are not major determinants of host range restriction and virulence. J. Virol. 1993, 67, 2448–2455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knipping, K.; McNeal, M.M.; Crienen, A.; Amerongen, G.V.; Garssen, J.; Land, B.V. A gastrointestinal rotavirus infection mouse model for immune modulation studies. Virol. J. 2011, 8, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortez, V.; Boyd, D.F.; Crawford, J.C.; Sharp, B.; Livingston, B.; Rowe, H.M.; Cherry, S.S. Astrovirus infects actively secreting goblet cells and alters the gut mucus barrier. Nat. Commun. 2020, 11, 2097. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, C.C.; Loh, J.; Zhao, G.; Stappenbeck, T.S.; Wang, D.; Huang, H.V.; Thackray, L.B. Adaptive immunity restricts replication of novel murine astroviruses. J. Virol. 2012, 86, 12262–12270. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Wilen, C.B.; Orvedahl, A.; McCune, B.T.; Kim, K.W.; Orchard, R.C.; Virgin, H.W. Norovirus cell tropism is determined by combinatorial action of a viral non-structural protein and host cytokine. Cell Host Microbe 2017, 22, 449–459. [Google Scholar] [CrossRef] [Green Version]
- Nice, T.J.; Strong, D.W.; McCune, B.T.; Pohl, C.S.; Virgin, H.W. A single-amino-acid change in murine norovirus NS1/2 is sufficient for colonic tropism and persistence. J. Virol. 2013, 87, 327–334. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Regev, D.; Watanabe, M.; Hickman, D.; Moussatche, N.; Jesus, D.M.; Karst, S.M. Identification of immune and viral correlates of norovirus protective immunity through comparative study of intra-cluster norovirus strains. PLoS Pathog. 2013, 9, e1003592. [Google Scholar] [CrossRef] [Green Version]
- Tomov, V.T.; Palko, O.; Lau, C.W.; Pattekar, A.; Sun, Y.; Tacheva, R.; Wherry, E.J. Differentiation and protective capacity of virus-specific CD8(+) T cells suggest murine norovirus persistence in an immune-privileged enteric niche. Immunity 2017, 47, 723–738. [Google Scholar] [CrossRef] [Green Version]
- Kounatidis, I.; Ligoxygakis, P. Drosophila as a model system to unravel the layers of innate immunity to infection. Open Biol. 2012, 2, 120075. [Google Scholar] [CrossRef] [Green Version]
- Lemaitre, B.; Hoffmann, J. The host defense of drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef] [Green Version]
- Segrist, E.; Cherry, S. Using diverse model systems to define intestinal epithelial defenses to enteric viral infections. Cell Host Microbe 2020, 27, 329–344. [Google Scholar] [CrossRef] [PubMed]
- Sansone, C.L.; Cohen, J.; Yasunaga, A.; Xu, J.; Osborn, G.; Subramanian, H.; Cherry, S. Microbiota-dependent priming of antiviral intestinal immunity in drosophila. Cell Host Microbe 2015, 18, 571–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Hopkins, K.; Sabin, L.; Yasunaga, A.; Subramanian, H.; Lamborn, I.; Cherry, S. ERK signaling couples nutrient status to antiviral defense in the insect gut. Proc. Natl. Acad. Sci. USA 2013, 110, 15025–15030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Felix, M.A.; Wang, D. Natural viruses of caenorhabditis nematodes. Annu. Rev. Genet. 2019, 53, 313–326. [Google Scholar] [CrossRef]
- Martin, A.; Rex, E.A.; Ishidate, T.; Lin, R.; Gammon, D.B. Infection of Caenorhabditis elegans with vesicular stomatitis virus via microinjection. Bio. Protoc. 2017, 7. [Google Scholar] [CrossRef]
- Hewes, S.A.; Wilson, R.L.; Estes, M.K.; Shroyer, N.F.; Blutt, S.E.; Allen, K.J.G. In vitro models of the small intestine: Engineering challenges and engineering solutions. Tissue Eng. Part B Rev. 2020. [Google Scholar] [CrossRef]
- Pearce, S.C.; Coia, H.G.; Karl, J.P.; Feliciano, I.G.P.; Zachos, N.C.; Racicot, K. Intestinal in vitro and ex vivo models to study host-microbiome interactions and acute stressors. Front. Physiol. 2018, 9, 1584. [Google Scholar] [CrossRef] [Green Version]
- Le, M.L.V.; Servin, A.L. Pathogenesis of human enterovirulent bacteria: Lessons from cultured, fully differentiated human colon cancer cell lines. Microbiol. Mol. Biol. Rev. 2013, 77, 380–439. [Google Scholar] [CrossRef] [Green Version]
- Oka, T.; Stoltzfus, G.T.; Zhu, C.; Jung, K.; Wang, Q.; Saif, L.J. Attempts to grow human noroviruses, a sapovirus, and a bovine norovirus in vitro. PLoS ONE 2018, 13, e0178157. [Google Scholar] [CrossRef]
- Straub, T.M.; Bartholomew, R.A.; Valdez, C.O.; Valentine, N.B.; Dohnalkova, A.; Ozanich, R.M.; Call, D.R. Human norovirus infection of caco-2 cells grown as a three-dimensional tissue structure. J. Water Health 2011, 9, 225–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svensson, L.; Finlay, B.B.; Bass, D.; Bonsdorff, C.H.V.; Greenberg, H.B. Symmetric infection of rotavirus on polarized human intestinal epithelial (Caco-2) cells. J. Virol. 1991, 65, 4190–4197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaibi, C.; Laffitte, J.C.; Sandre, C.; Esclatine, A.; Servin, A.L.; Quero, A.M.; Legendre, M.G. Rotavirus induces apoptosis in fully differentiated human intestinal Caco-2 cells. Virology 2005, 332, 480–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vu, D.L.; Bosch, A.; Pinto, R.M.; Ribes, E.; Guix, S. Human astrovirus MLB replication in vitro: Persistence in extraintestinal cell lines. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [Green Version]
- Meliopoulos, V.A.; Marvin, S.A.; Freiden, P.; Moser, L.A.; Nighot, P.; Ali, R.; Cherry, S.S. Oral administration of astrovirus capsid protein is sufficient to induce acute diarrhea in vivo. mBio 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Moon, S.; Wang, Y.; Jiang, B. Multiple virus infection alters rotavirus replication and expression of cytokines and Toll-like receptors in intestinal epithelial cells. Virus Res. 2012, 167, 48–55. [Google Scholar] [CrossRef]
- Altamimi, M.; Abdelhay, O.; Rastall, R.A. Effect of oligosaccharides on the adhesion of gut bacteria to human HT-29 cells. Anaerobe 2016, 39, 136–142. [Google Scholar] [CrossRef]
- Raja, S.B.; Murali, M.R.; Devaraj, H.; Devaraj, S.N. Differential expression of gastric MUC5AC in colonic epithelial cells: TFF3-wired IL1 beta/Akt crosstalk-induced mucosal immune response against shigella dysenteriae infection. J. Cell Sci. 2012, 125, 703–713. [Google Scholar] [CrossRef] [Green Version]
- Rajan, A.; Persson, B.D.; Frangsmyr, L.; Olofsson, A.; Sandblad, L.; Heino, J.; Arnberg, N. Enteric species F human adenoviruses use laminin-binding integrins as co-receptors for infection of Ht-29 Cells. Sci. Rep. 2018, 8, 10019. [Google Scholar] [CrossRef]
- Chattopadhyay, S.; Basak, T.; Nayak, M.K.; Bhardwaj, G.; Mukherjee, A.; Bhowmick, R.; Sarkar, M.C. Identification of cellular calcium binding protein calmodulin as a regulator of rotavirus A infection during comparative proteomic study. PLoS ONE 2013, 8, e56655. [Google Scholar] [CrossRef] [Green Version]
- Chanda, S.; Nandi, S.; Sarkar, M.C. Rotavirus-induced miR-142-5p elicits proviral milieu by targeting non-canonical transforming growth factor beta signalling and apoptosis in cells. Cell Microbiol. 2016, 18, 733–747. [Google Scholar] [CrossRef]
- Sato, T.; Vries, R.G.; Snippert, H.J.; Wetering, M.V.d.; Barker, N.; Stange, D.E.; Clevers, H. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature 2009, 459, 262–265. [Google Scholar] [CrossRef]
- Sato, T.; Stange, D.E.; Ferrante, M.; Vries, R.G.; Van, J.H.; Brink, S.V.D.; Clevers, H. Long-term expansion of epithelial organoids from human colon, adenoma, adenocarcinoma, and Barrett’s epithelium. Gastroenterology 2011, 141, 1762–1772. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Clevers, H. Growing self-organizing mini-guts from a single intestinal stem cell: Mechanism and applications. Science 2013, 340, 1190–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zachos, N.C.; Kovbasnjuk, O.; Abel, J.F.; In, J.; Blutt, S.E.; Jonge, H.R.d.; Donowitz, M. Human enteroids/colonoids and intestinal organoids functionally recapitulate normal intestinal physiology and pathophysiology. J. Biol. Chem. 2016, 291, 3759–3766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxena, K.; Blutt, S.E.; Ettayebi, K.; Zeng, X.L.; Broughman, J.R.; Crawford, S.E.; Estes, M.K. Human intestinal enteroids: A new model to study human rotavirus infection, host restriction, and pathophysiology. J. Virol. 2015, 90, 43–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxena, K.; Simon, L.M.; Zeng, X.L.; Blutt, S.E.; Crawford, S.E.; Sastri, N.P.; Estes, M.K. A paradox of transcriptional and functional innate interferon responses of human intestinal enteroids to enteric virus infection. Proc. Natl. Acad. Sci. USA 2017, 114, E570–E579. [Google Scholar] [CrossRef] [Green Version]
- Ettayebi, K.; Crawford, S.E.; Murakami, K.; Broughman, J.R.; Karandikar, U.; Tenge, V.R.; Estes, M.K. Replication of human noroviruses in stem cell-derived human enteroids. Science 2016, 353, 1387–1393. [Google Scholar] [CrossRef]
- Drummond, C.G.; Bolock, A.M.; Ma, C.; Luke, C.J.; Good, M.; Coyne, C.B. Enteroviruses infect human enteroids and induce antiviral signaling in a cell lineage-specific manner. Proc. Natl. Acad. Sci. USA 2017, 114, 1672–1677. [Google Scholar] [CrossRef] [Green Version]
- Good, C.; Wells, A.I.; Coyne, C.B. Type III interferon signaling restricts enterovirus 71 infection of goblet cells. Sci. Adv. 2019, 5, 4255. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Potential consequences of enteric viral co-infections. (a) Co-infection of multiple viral strains in a single cell facilitates the recombination and generation of novel strains. (b) Extracellular vesicles containing multiple virions of a single virus type, one means of co-infecting new cells. Whether co-infection with unrelated viruses could lead to co-packaging is unknown. (c) An example of enteric viral interference, wherein murine AstV infection inhibits murine NoV infection of tuft cells via IFN-λ production.
Figure 1.
Potential consequences of enteric viral co-infections. (a) Co-infection of multiple viral strains in a single cell facilitates the recombination and generation of novel strains. (b) Extracellular vesicles containing multiple virions of a single virus type, one means of co-infecting new cells. Whether co-infection with unrelated viruses could lead to co-packaging is unknown. (c) An example of enteric viral interference, wherein murine AstV infection inhibits murine NoV infection of tuft cells via IFN-λ production.

Figure 2.
In vivo and in vitro co-infection study models. In vivo models for enteric virus studies include mouse (murine NoV, AstV, and RV), fly (Drosophila C virus, Flock House virus), pig (human RV, human NoV, and porcine epidemic diarrhea virus), C. elegans (Orsay virus) and non-human primate (human RV and NoV, simian RV, and rhesus enteric caliciviruses) models with physiological experimental read-outs. In vitro models that could be used for enteric co-infection studies include organoids/enteroids and immortalized cell lines for detailed analyses in simplified systems. Illustration created using BioRender (BioRender.com).
Figure 2.
In vivo and in vitro co-infection study models. In vivo models for enteric virus studies include mouse (murine NoV, AstV, and RV), fly (Drosophila C virus, Flock House virus), pig (human RV, human NoV, and porcine epidemic diarrhea virus), C. elegans (Orsay virus) and non-human primate (human RV and NoV, simian RV, and rhesus enteric caliciviruses) models with physiological experimental read-outs. In vitro models that could be used for enteric co-infection studies include organoids/enteroids and immortalized cell lines for detailed analyses in simplified systems. Illustration created using BioRender (BioRender.com).


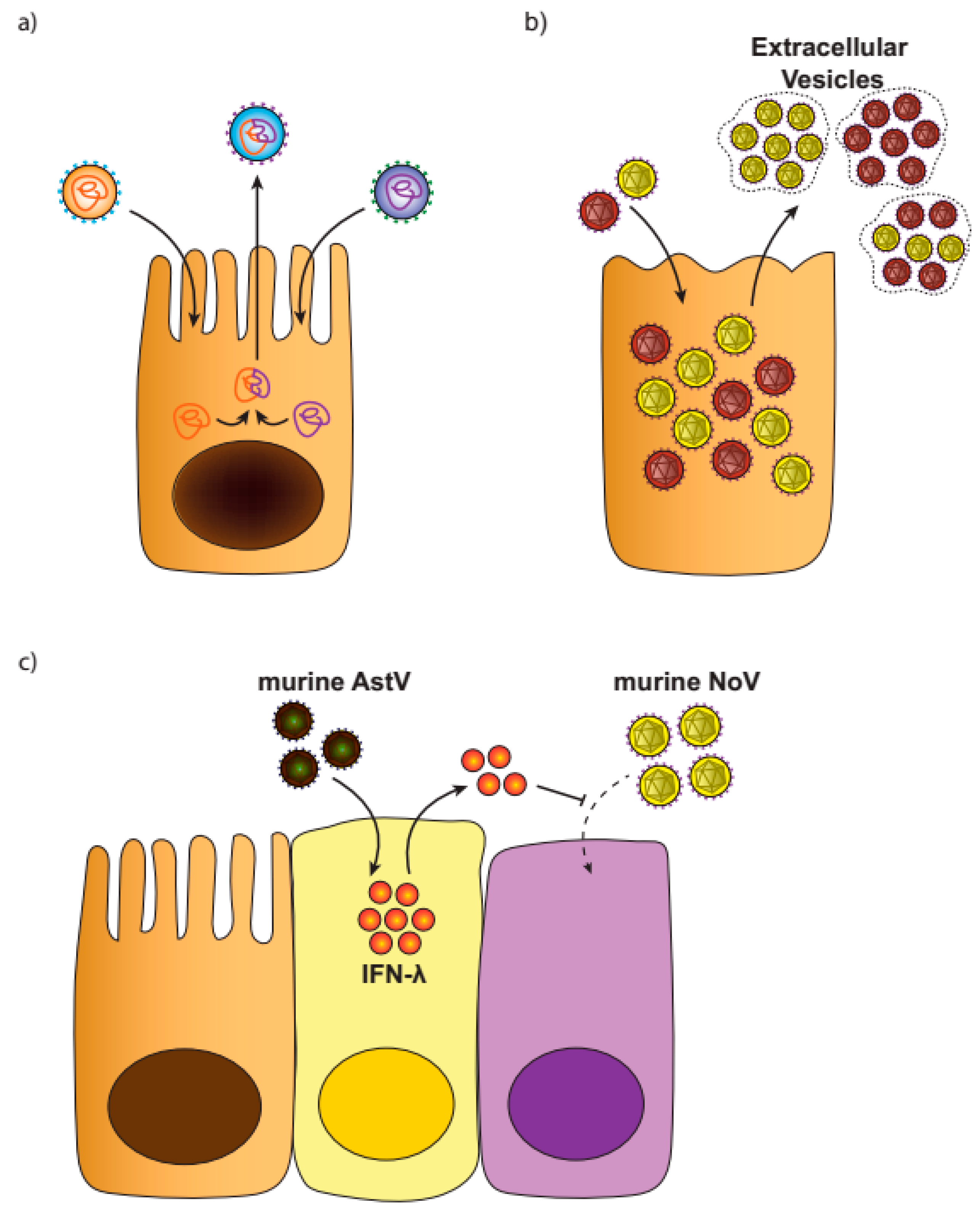{kind=link}
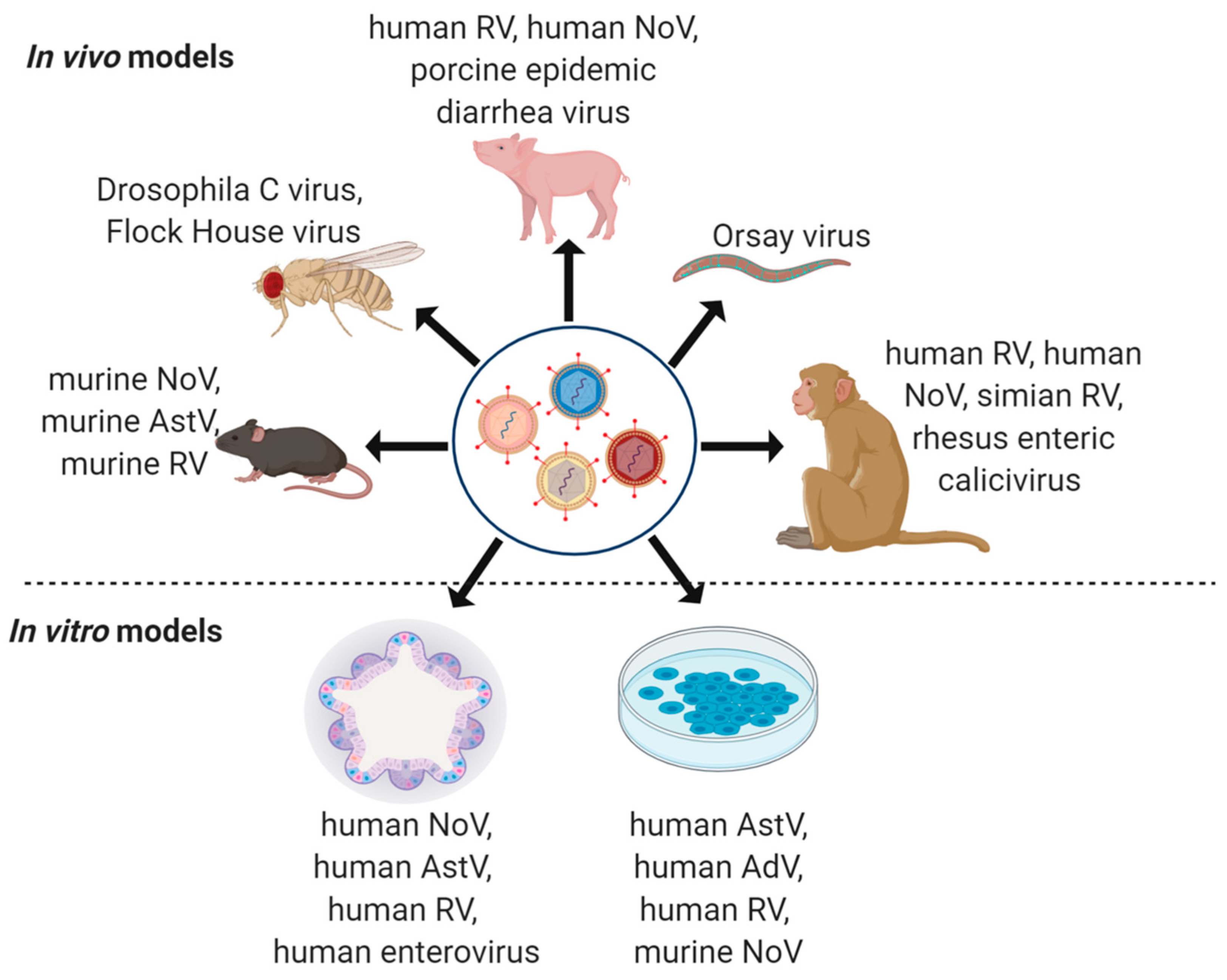{kind=link}
Table 1.
Summary of major enteric viral pathogens, many of which have been frequently implicated in virus–virus co-infection events.
Table 1.
Summary of major enteric viral pathogens, many of which have been frequently implicated in virus–virus co-infection events.
| Classification | Genome | Major Agents | Disease | Prevalence | Reference |
|---|---|---|---|---|---|
| Class I Adenoviridae | dsDNA 30–40 kb | Adenovirus (AdV) serotypes 40, 41 (species F) | Gastroenteritis | 15% in children and 1.5 to 5.4% in adults | [5] |
| Class III Reoviridae | dsRNA 16–27 kb | Rotavirus (RV) | Gastroenteritis | In children <5: pre-vaccine 40% post-vaccine 19% | [5,6] |
| Class IV Astroviridae | ssRNA (+) 6.8–7.9 kb | Astrovirus (AstV) | Nosocomial or epidemic diarrhea | ~5% in children with AGE, 3–9% in hospitalized patients with diarrhea | [5,7] |
| Class IV Caliciviridae | ssRNA (+) 7.4–8.3 kb | Norovirus (NoV), Sapovirus (SaV) | Epidemic diarrhea | 18% in all AGE cases, as well as 24% in community | [5,8,9] |
| Class IV Picornaviridae | ssRNA (+) 7.1–8.9 kb | Enterovirus species A and B (Enteroviruses and Coxsackieviruses), species C (Polioviruses) | Gastroenteritis, Hand, foot, and mouth disease (HFMD), Poliomyelitis | ~5% in children with AGE; ~4% in children with HFMD | [10] |
| Class IV Coronaviridae | ssRNA (+) 27–30 kb | Severe acute respiratory syndrome (SARS-CoV) and severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) | Acute diarrhea, severe acute respiratory syndrome, and coronavirus disease 2019 (COVID-19) | Ongoing global pandemic—to be determined | [11,12,13] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Makimaa, H.; Ingle, H.; Baldridge, M.T. Enteric Viral Co-Infections: Pathogenesis and Perspective. Viruses 2020, 12, 904. https://0-doi-org.brum.beds.ac.uk/10.3390/v12080904
AMA Style
Makimaa H, Ingle H, Baldridge MT. Enteric Viral Co-Infections: Pathogenesis and Perspective. Viruses. 2020; 12(8):904. https://0-doi-org.brum.beds.ac.uk/10.3390/v12080904
Chicago/Turabian StyleMakimaa, Heyde, Harshad Ingle, and Megan T. Baldridge. 2020. "Enteric Viral Co-Infections: Pathogenesis and Perspective" Viruses 12, no. 8: 904. https://0-doi-org.brum.beds.ac.uk/10.3390/v12080904
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.