Loop-Mediated Isothermal Amplification (LAMP) Assay for Rapid and Accurate Confirmatory Diagnosis of HTLV-1/2 Infection

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Population Data
2.2. Design of LAMP Primers
2.3. HTLV-1/2 LAMP Assays
2.4. Quantification of HTLV-1/2 Proviral Load
2.5. PCR-RFLP for HTLV-1/2
2.6. Statistical Analysis
3. Results
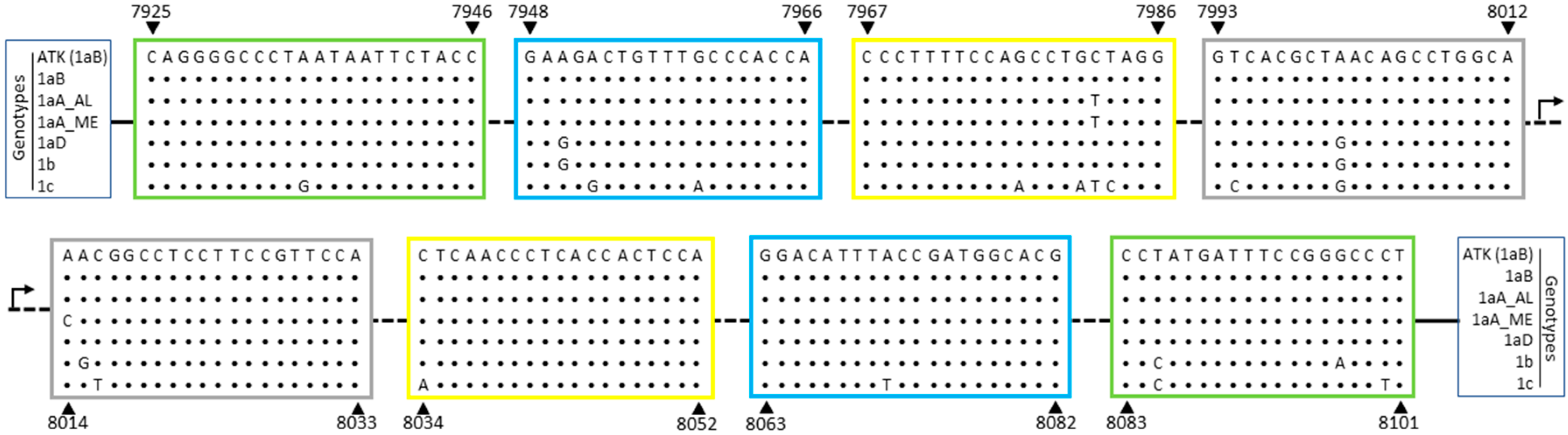
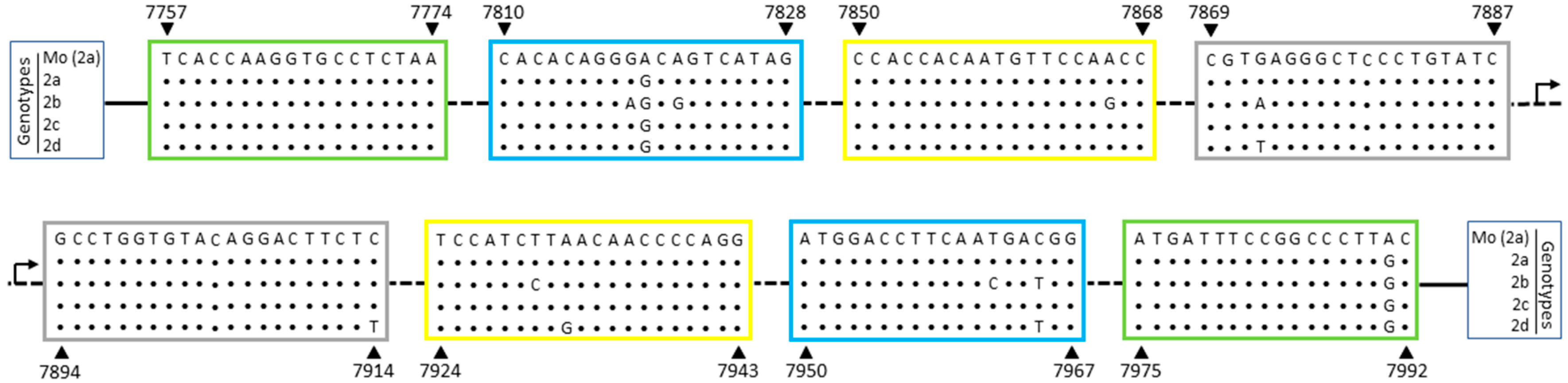
3.1. Genetic Polymorphisms within LAMP Primers for HTLV-1/2
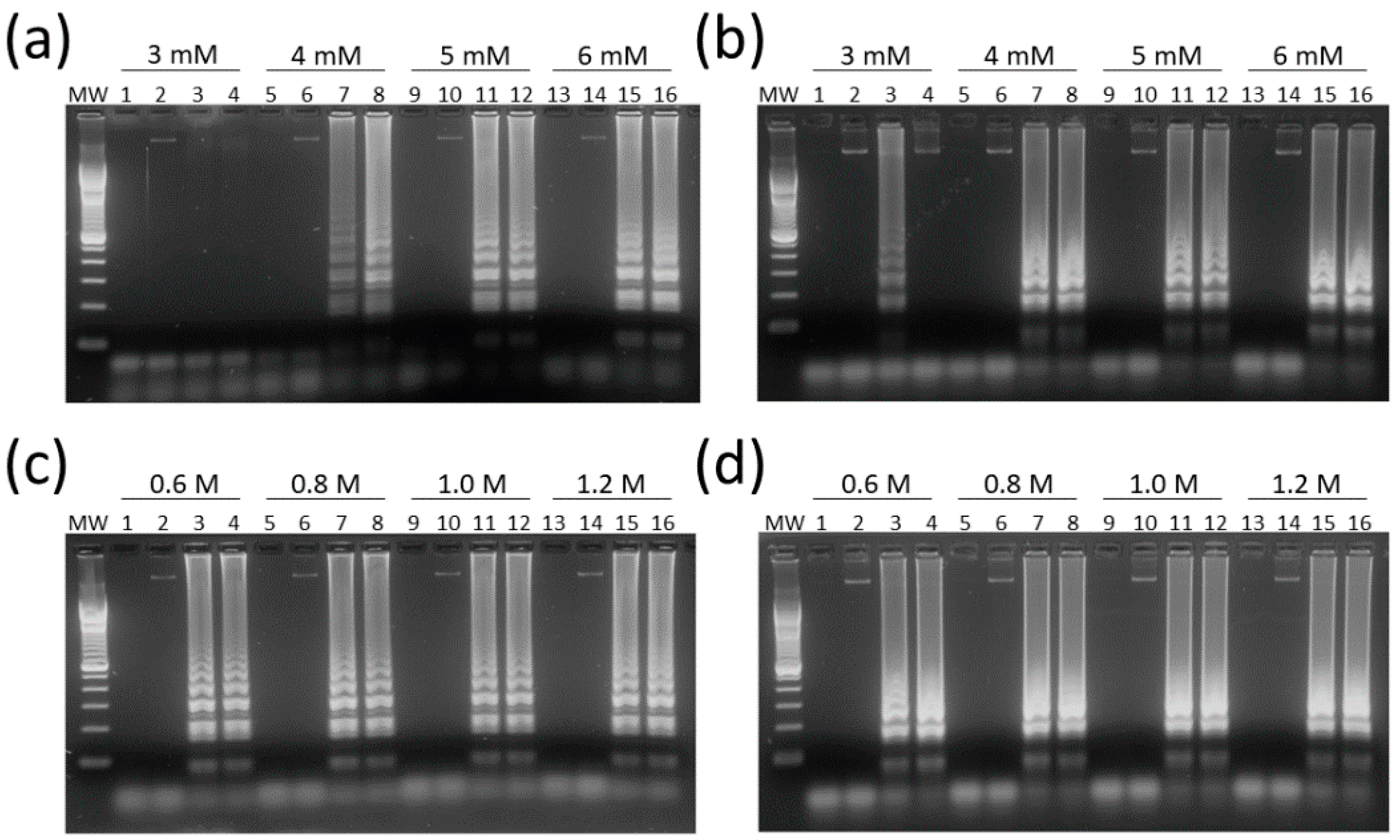
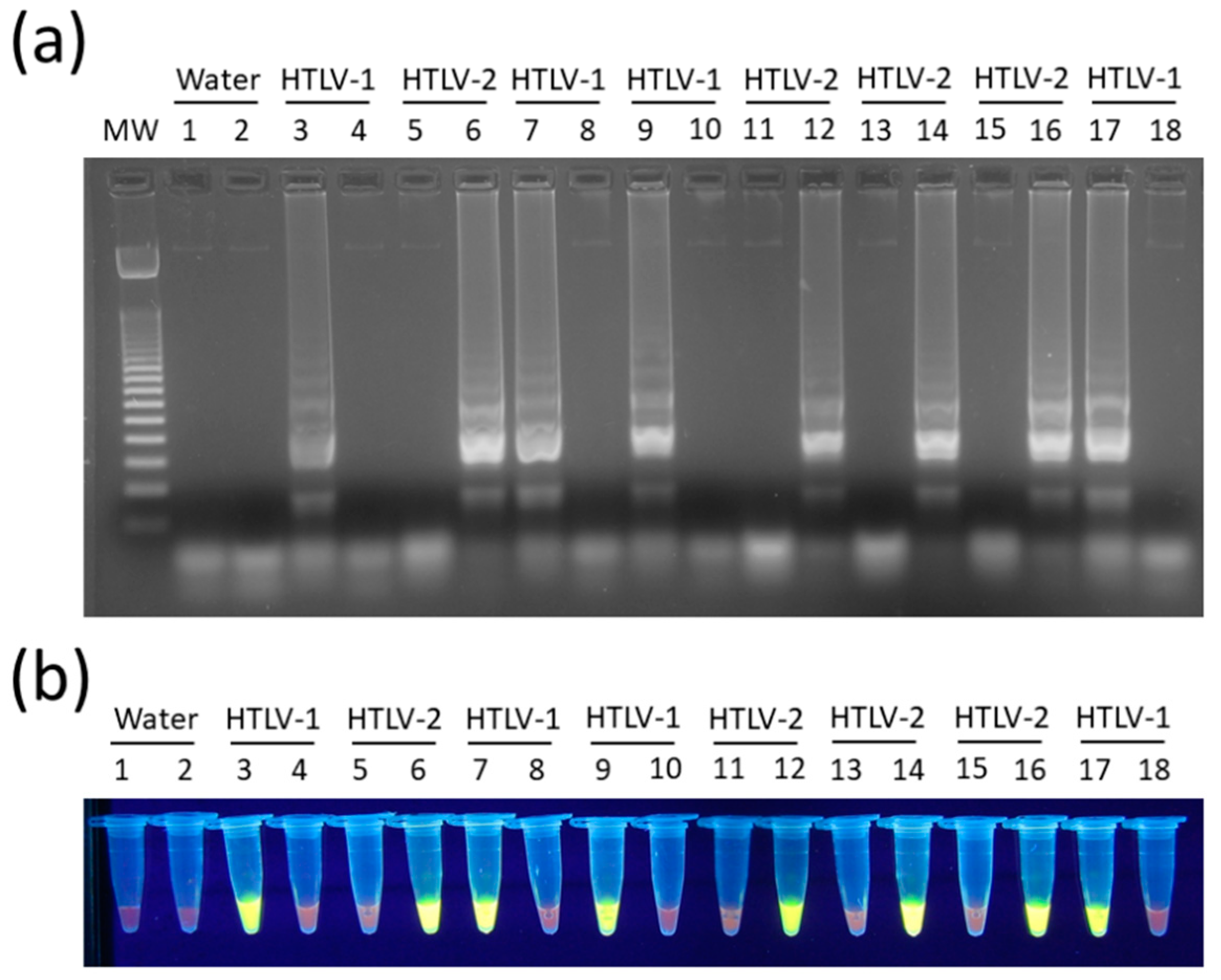
3.2. Optimization and Target Specificity of HTLV-1/2 LAMP Reactions
3.3. Overall Accuracy and Sensitivity of HTLV-1/2 LAMP Assays
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Proietti, F.A.; Carneiro-Proietti, A.B.F.; Catalan-Soares, B.C.; Murphy, E.L. Global epidemiology of HTLV-I infection and associated diseases. Oncogene 2005, 24, 6058–6068. [Google Scholar] [CrossRef] [Green Version]
- Hall, W.W.; Kubo, T.; Ijichi, S.; Takahashi, H.; Zhu, S.W. Human T cell leukemia/lymphoma virus, type II (HTLV-II): Emergence of an important newly recognized pathogen. Seminars Virol. 1994, 5, 165–178. [Google Scholar] [CrossRef]
- Hall, W.W.; Ishak, R.; Zhu, S.W.; Novoa, P.; Eiraku, N.; Takahashi, H.; da Ferreira, M.C.; Azevedo, V.; Ishak, M.O.; da Ferreira, O.C.; et al. Human T lymphotropic virus type II (HTLV-II): Epidemiology, molecular properties, and clinical features of infection. J. Acquir. Immune Defic. Syndr. Hum. Retrovirol. 1996, 1, S204–S214. [Google Scholar] [CrossRef]
- Catalan-Soares, B.; de Carneiro-Proietti, A.B.F.; Proietti, F.A. Heterogeneous geographic distribution of human T-cell lymphotropic viruses I and II (HTLV-I/II): Serological screening prevalence rates in blood donors from large urban areas in Brazil. Cad. Saúde Pública 2005, 21, 926–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauclère, P.; Afonso, P.V.; Meertens, L.; Plancoulaine, S.; Calattini, S.; Froment, A.; Van Beveren, M.; de Thé, G.; Quintana-Murci, L.; Mahieux, R.; et al. HTLV-2B Strains, Similar to Those Found in Several Amerindian Tribes, Are Endemic in Central African Bakola Pygmies. J. Infect. Dis. 2011, 203, 1316–1323. [Google Scholar] [CrossRef] [PubMed]
- Gessain, A.; Cassar, O. Epidemiological Aspects and World Distribution of HTLV-1 Infection. Front. Microbiol. 2012, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Einsiedel, L.; Woodman, R.J.; Flynn, M.; Wilson, K.; Cassar, O.; Gessain, A. Human T-Lymphotropic Virus type 1 infection in an Indigenous Australian population: Epidemiological insights from a hospital-based cohort study. BMC Public Health 2016, 16, 787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Time to Eradicate HTLV-1: An Open Letter to WHO. Available online: http://www.gvn.org/who (accessed on 14 May 2020).
- Tagaya, Y.; Matsuoka, M.; Gallo, R. 40 years of the human T-cell leukemia virus: Past, present, and future. F1000Res 2019, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.B.; Kaidarova, Z.; Hindes, D.; Bravo, M.; Kiely, N.; Kamel, H.; Dubay, D.; Hoose, B.; Murphy, E.L. Seroprevalence and demographic determinants of human T-lymphotropic virus type 1 and 2 infections among first-time blood donors—United States, 2000–2009. J. Infect. Dis. 2014, 209, 523–531. [Google Scholar] [CrossRef]
- Caterino-de-Araujo, A.; Alves, F.A.; Campos, K.R.; Lemos, M.F.; Moreira, R.C. Making the invisible visible: Searching for human T-cell lymphotropic virus types 1 and 2 (HTLV-1 and HTLV-2) in Brazilian patients with viral hepatitis B and C. Mem. Inst. Oswaldo. Cruz. 2018, 113, 130–134. [Google Scholar] [CrossRef]
- Beilke, M.A. Retroviral coinfections: HIV and HTLV: Taking stock of more than a quarter century of research. AIDS Res. Hum. Retrovir. 2012, 28, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.T.; de Melo Espíndola, O.; Bezerra Leite, A.C.C.; Araújo, A. Neurological aspects of HIV/human T lymphotropic virus coinfection. AIDS Rev. 2009, 11, 71–78. [Google Scholar]
- Goubau, P.; Wantzin, P.; Gessain, A.; Jeannel, D.; Coste, J. Seroepidemiology of the Human T-Cell Leukaemia/Lymphoma Viruses in Europe. JAIDS J. Acquir. Immune Defic. Syndr. 1996, 13, 68. [Google Scholar]
- Ramos, G.; Ricardo, L.; Umberto, B.; Cardoso, T.; Tetelbom, S.; Simon, D. Short Communication: Prevalence and Risk Factors for Human T Cell Lymphotropic Virus Infection in Southern Brazilian HIV-Positive Patients. AIDS Res. Hum. Retrovir. 2014, 30, 9. [Google Scholar] [CrossRef]
- Araujo, A.Q.C.; Silva, M.T.T. The HTLV-1 neurological complex. Lancet Neurol. 2006, 5, 1068–1076. [Google Scholar] [CrossRef]
- Araujo, A.Q.C.; Wedemann, D. HTLV-1 Associated Neurological Complex. What is Hidden below the Water? AIDS Rev. 2019, 21, 211–217. [Google Scholar] [CrossRef]
- Watanabe, T. Adult T-cell leukemia: Molecular basis for clonal expansion and transformation of HTLV-1–infected T cells. Blood 2017, 129, 1071–1081. [Google Scholar] [CrossRef] [Green Version]
- Hirata, T.; Uchima, N.; Kishimoto, K.; Zaha, O.; Kinjo, N.; Hokama, A.; Sakugawa, H.; Kinjo, F.; Fujita, J. Impairment of host immune response against strongyloides stercoralis by human T cell lymphotropic virus type 1 infection. Am. J. Trop. Med. Hyg. 2006, 74, 246–249. [Google Scholar] [CrossRef] [Green Version]
- Okajima, R.; Oliveira, A.C.P.; Smid, J.; Casseb, J.; Sanches, J.A. High prevalence of skin disorders among HTLV-1 infected individuals independent of clinical status. PLoS Negl. Trop. Dis. 2013, 7, e2546. [Google Scholar] [CrossRef] [Green Version]
- Einsiedel, L.J.; Pepperill, C.; Wilson, K. Crusted scabies: A clinical marker of human T-lymphotropic virus type 1 infection in central Australia. Med. J. Aust. 2014, 200, 633–634. [Google Scholar] [CrossRef] [Green Version]
- Martin, F.; Taylor, G.P.; Jacobson, S. Inflammatory manifestations of HTLV-1 and their therapeutic options. Expert Rev. Clin. Immunol. 2014, 10, 1531–1546. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, N.B.; de Lourdes Bastos, M.; Souza, A.S.; Netto, E.M.; Arruda, S.; Santos, S.B.; Carvalho, E.M. Impaired TNF, IL-1β, and IL-17 production and increased susceptibility to Mycobacterium tuberculosis infection in HTLV-1 infected individuals. Tuberculosis (Edinb) 2018, 108, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Thorstensson, R.; Albert, J.; Andersson, S. Strategies for diagnosis of HTLV-I and -II. Transfusion 2002, 42, 780–791. [Google Scholar] [CrossRef] [PubMed]
- Costa, E.A.S.; Magri, M.C.; Caterino-de-Araujo, A. The best algorithm to confirm the diagnosis of HTLV-1 and HTLV-2 in at-risk individuals from São Paulo, Brazil. J. Virol. Methods 2011, 173, 280–286. [Google Scholar] [CrossRef]
- Poiesz, B.J.; Dube, S.; Choi, D.; Esteban, E.; Ferrer, J.; Leon-Ponte, M.; de Perez, G.E.; Glaser, J.; Devare, S.G.; Vallari, A.S.; et al. Comparative performances of an HTLV-I/II EIA and other serologic and PCR assays on samples from persons at risk for HTLV-II infection. Transfusion 2000, 40, 924–930. [Google Scholar] [CrossRef]
- Campos, K.R.; Santos, F.L.N.; da Silva Brito, V.; Gonçalves, N.L.S.; Araujo, T.H.A.; Galvão-Castro, B.; Caterino-de-Araujo, A. Line Immunoassay for Confirmation and Discrimination of Human T-Cell Lymphotropic Virus Infections in Inconclusive Western Blot Serum Samples from Brazil. J. Clin. Microbiol. 2019, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garin, B.; Gosselin, S.; de Thé, G.; Gessain, A. HTLV-I/II infection in a high viral endemic area of Zaire, Central Africa: Comparative evaluation of serology, PCR, and significance of indeterminate western blot pattern. J. Med. Virol. 1994, 44, 104–109. [Google Scholar] [CrossRef]
- Morimoto, H.K.; Morimoto, A.A.; Reiche, E.M.V.; Ueda, L.T.; Matsuo, T.; Reiche, F.V.; Caterino-de-Araujo, A. Difficulties in the diagnosis of HTLV-2 infection in HIV/AIDS patients from Brazil: Comparative performances of serologic and molecular assays, and detection of HTLV-2b subtype. Rev. Inst. Med. Trop. São Paulo 2007, 49, 225–230. [Google Scholar] [CrossRef] [Green Version]
- Novoa, P.; Penalva de Oliveira, A.C.; Posada Vergara, M.P.; da Silva Duarte, A.J.; Casseb, J. Molecular characterization of human T-cell lymphotropic virus type 2 (HTLV-II) from people living in urban areas of Sao Paulo city: Evidence of multiple subtypes circulation. J. Med. Virol. 2007, 79, 182–187. [Google Scholar] [CrossRef]
- Olah, I.; Fukumori, L.M.I.; Smid, J.; de Oliveira, A.C.P.; Duarte, A.J.S.; Casseb, J. Neither molecular diversity of the envelope, immunosuppression status, nor proviral load causes indeterminate HTLV western blot profiles in samples from human T-cell lymphotropic virus type 2 (HTLV-2)-infected individuals. J. Med. Virol. 2010, 82, 837–842. [Google Scholar] [CrossRef]
- Jacob, F.; Santos-Fortuna, E.; Azevedo, R.S.; Caterino-de-Araujo, A. Serological patterns and temporal trends of HTLV-1/2 infection in high-risk populations attending Public Health Units in São Paulo, Brazil. J. Clin. Virol. 2008, 42, 149–155. [Google Scholar] [CrossRef] [PubMed]
- da Silva Brito, V.; Santos, F.L.N.; Gonçalves, N.L.S.; Araujo, T.H.A.; Nascimento, D.S.V.; Pereira, F.M.; Boa-Sorte, N.C.A.; Grassi, M.F.R.; Caterino-de-Araujo, A.; Galvão-Castro, B. Performance of Commercially Available Serological Screening Tests for Human T-Cell Lymphotropic Virus Infection in Brazil. J. Clin. Microbiol. 2018, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, K.R.; Gonçalves, M.G.; Costa, N.A.; Caterino-de-Araujo, A. Comparative performances of serologic and molecular assays for detecting human T lymphotropic virus type 1 and type 2 (HTLV-1 and HTLV-2) in patients infected with human immunodeficiency virus type 1 (HIV-1). Braz. J. Infect. Dis. 2017, 21, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, E63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, T.; Lou, G.; Yang, J.; Xu, D.; Meng, Z. Development and evaluation of real-time loop-mediated isothermal amplification for hepatitis B virus DNA quantification: A new tool for HBV management. J. Clin. Virol. 2008, 41, 270–276. [Google Scholar] [CrossRef]
- Curtis, K.A.; Rudolph, D.L.; Owen, S.M. Rapid detection of HIV-1 by reverse-transcription, loop-mediated isothermal amplification (RT-LAMP). J. Virol. Methods 2008, 151, 264–270. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, J.; Hu, J.; Chen, H.; Du, L.; Wu, L.; Ding, Y.; Xiong, S.; Huang, X.; Zhang, Y.; et al. Rapid detection of hepatitis C virus RNA by a reverse transcription loop-mediated isothermal amplification assay. FEMS Immunol. Med. Microbiol. 2011, 63, 144–147. [Google Scholar] [CrossRef]
- Baek, Y.H.; Um, J.; Antigua, K.J.C.; Park, J.-H.; Kim, Y.; Oh, S.; Kim, Y.-I.; Choi, W.-S.; Kim, S.G.; Jeong, J.H.; et al. Development of a reverse transcription-loop-mediated isothermal amplification as a rapid early-detection method for novel SARS-CoV-2. Emerg. Microbes Infect. 2020, 9, 998–1007. [Google Scholar] [CrossRef] [Green Version]
- Reddy, A.K.; Balne, P.K.; Reddy, R.K.; Mathai, A.; Kaur, I. Loop-mediated isothermal amplification assay for the diagnosis of retinitis caused by herpes simplex virus-1. Clin. Microbiol. Infect. 2011, 17, 210–213. [Google Scholar] [CrossRef] [Green Version]
- Kubo, T.; Agoh, M.; Mai, L.Q.; Fukushima, K.; Nishimura, H.; Yamaguchi, A.; Hirano, M.; Yoshikawa, A.; Hasebe, F.; Kohno, S.; et al. Development of a reverse transcription-loop-mediated isothermal amplification assay for detection of pandemic (H1N1) 2009 virus as a novel molecular method for diagnosis of pandemic influenza in resource-limited settings. J. Clin. Microbiol. 2010, 48, 728–735. [Google Scholar] [CrossRef] [Green Version]
- Reddy, A.K.; Balne, P.K.; Reddy, R.K.; Mathai, A.; Kaur, I. Development and evaluation of loop-mediated isothermal amplification assay for rapid and inexpensive detection of cytomegalovirus DNA in vitreous specimens from suspected cases of viral retinitis. J. Clin. Microbiol. 2010, 48, 2050–2052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parida, M.; Sannarangaiah, S.; Dash, P.K.; Rao, P.V.L.; Morita, K. Loop mediated isothermal amplification (LAMP): A new generation of innovative gene amplification technique; perspectives in clinical diagnosis of infectious diseases. Rev. Med. Virol. 2008, 18, 407–421. [Google Scholar] [CrossRef] [PubMed]
- PrimerExplorer V5. Available online: http://primerexplorer.jp/lampv5e/index.html. (accessed on 10 October 2019).
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tateno, M.; Kondo, N.; Itoh, T.; Chubachi, T.; Togashi, T.; Yoshiki, T. Rat lymphoid cell lines with human T cell leukemia virus production. I. Biological and serological characterization. J. Exp. Med. 1984, 159, 1105–1116. [Google Scholar] [CrossRef] [PubMed]
- Kashima, S.; Alcantara, L.C.; Takayanagui, O.M.; Cunha, M.A.V.; Castro, B.G.; Pombo-de-Oliveira, M.S.; Zago, M.A.; Covas, D.T. Distribution of human T cell lymphotropic virus type 1 (HTLV-1) subtypes in Brazil: Genetic characterization of LTR and tax region. AIDS Res. Hum. Retrovir. 2006, 22, 953–959. [Google Scholar] [CrossRef]
- Paiva, A.; Casseb, J.; Paiva, A.; Casseb, J. Origin and prevalence of human T-lymphotropic virus type 1 (HTLV-1) and type 2 (HTLV-2) among indigenous populations in the Americas. Rev. Inst. Med. Trop. São Paulo 2015, 57, 01–14. [Google Scholar] [CrossRef]
- Braço, I.L.J.; de Sá, K.S.G.; Waqasi, M.; Queiroz, M.A.F.; da Silva, A.N.R.; Cayres-Vallinoto, I.M.V.; Lima, S.S.; de Oliveira Guimarães Ishak, M.; Ishak, R.; Guerreiro, J.F.; et al. High prevalence of human T-lymphotropic virus 2 (HTLV-2) infection in villages of the Xikrin tribe (Kayapo), Brazilian Amazon region. BMC Infect. Dis. 2019, 19, 459. [Google Scholar] [CrossRef] [Green Version]
- Meagher, R.J.; Priye, A.; Light, Y.K.; Huang, C.; Wang, E. Impact of Primer Dimers and Self-Amplifying Hairpins on Reverse Transcription Loop-Mediated Isothermal Amplification Detection of Viral RNA. Analyst 2018, 143, 1924–1933. [Google Scholar] [CrossRef]
- Wang, X.; Yin, F.; Bi, Y.; Cheng, G.; Li, J.; Hou, L.; Li, Y.; Yang, B.; Liu, W.; Yang, L. Rapid and sensitive detection of Zika virus by reverse transcription loop-mediated isothermal amplification. J. Virol. Methods 2016, 238, 86–93. [Google Scholar] [CrossRef]
- Gessain, A.; Yanagihara, R.; Franchini, G.; Garruto, R.M.; Jenkins, C.L.; Ajdukiewicz, A.B.; Gallo, R.C.; Gajdusek, D.C. Highly divergent molecular variants of human T-lymphotropic virus type I from isolated populations in Papua New Guinea and the Solomon Islands. Proc. Natl. Acad. Sci. USA 1991, 88, 7694–7698. [Google Scholar] [CrossRef] [Green Version]
- Afonso, P.V.; Cassar, O.; Gessain, A. Molecular epidemiology, genetic variability and evolution of HTLV-1 with special emphasis on African genotypes. Retrovirology 2019, 16, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassar, O.; Charavay, F.; Touzain, F.; Jeannin, P.; Grangeon, J.-P.; Laumond, S.; Chungue, E.; Martin, P.M.V.; Gessain, A. A Novel Human T-lymphotropic Virus Type 1c Molecular Variant in an Indigenous Individual from New Caledonia, Melanesia. PLoS Negl. Trop. Dis. 2017, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassar, O.; Einsiedel, L.; Afonso, P.V.; Gessain, A. Human T-Cell Lymphotropic Virus Type 1 Subtype C Molecular Variants among Indigenous Australians: New Insights into the Molecular Epidemiology of HTLV-1 in Australo-Melanesia. PLoS Negl. Trop. Dis. 2013, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassar, O.; Capuano, C.; Bassot, S.; Charavay, F.; Duprez, R.; Afonso, P.V.; Abel, M.; Walter, H.; Mera, W.; Martin, P.M.V.; et al. Human T lymphotropic virus type 1 subtype C melanesian genetic variants of the Vanuatu Archipelago and Solomon Islands share a common ancestor. J. Infect. Dis. 2007, 196, 510–521. [Google Scholar] [CrossRef] [PubMed]
- Soldan, S.S.; Graf, M.D.; Waziri, A.; Flerlage, A.N.; Robinson, S.M.; Kawanishi, T.; Leist, T.P.; Lehky, T.J.; Levin, M.C.; Jacobson, S. HTLV-I/II seroindeterminate Western blot reactivity in a cohort of patients with neurological disease. J. Infect. Dis. 1999, 180, 685–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cánepa, C.; Salido, J.; Ruggieri, M.; Fraile, S.; Pataccini, G.; Berini, C.; Biglione, M. Low Proviral Load is Associated with Indeterminate Western Blot Patterns in Human T-Cell Lymphotropic Virus Type 1 Infected Individuals: Could Punctual Mutations be Related? Viruses 2015, 7, 5643–5658. [Google Scholar] [CrossRef] [Green Version]
- Kuramitsu, M.; Sekizuka, T.; Yamochi, T.; Firouzi, S.; Sato, T.; Umeki, K.; Sasaki, D.; Hasegawa, H.; Kubota, R.; Sobata, R.; et al. Proviral Features of Human T Cell Leukemia Virus Type 1 in Carriers with Indeterminate Western Blot Analysis Results. J. Clin. Microbiol. 2017, 55, 2838–2849. [Google Scholar] [CrossRef] [Green Version]
- Martins, M.L.; da Santos, A.C.S.; Namen-Lopes, M.S.; Barbosa-Stancioli, E.F.; Utsch, D.G.; de Carneiro-Proietti, A.B.F. Long-term serological follow-up of blood donors with an HTLV-indeterminate western blot: Antibody profile of seroconverters and individuals with false reactions. J. Med. Virol. 2010, 82, 1746–1753. [Google Scholar] [CrossRef]
- Glowacka, I.; Korn, K.; Potthoff, S.A.; Lehmann, U.; Kreipe, H.H.; Ivens, K.; Barg-Hock, H.; Schulz, T.F.; Heim, A. Delayed seroconversion and rapid onset of lymphoproliferative disease after transmission of human T-cell lymphotropic virus type 1 from a multiorgan donor. Clin. Infect. Dis. 2013, 57, 1417–1424. [Google Scholar] [CrossRef] [Green Version]
- Dube, S.; Saksena, N.; Spicer, T.; Healey, J.; Benz, P.; Dube, D.K.; Poiesz, B.J. Delayed seroconversion to STLV-1 infection is associated with mutations in the pol and rex genes. Virol. J. 2013, 10, 282. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.; Ma, G.; Nosaka, K.; Tanabe, J.; Satou, Y.; Koito, A.; Wain-Hobson, S.; Vartanian, J.-P.; Matsuoka, M. APOBEC3G generates nonsense mutations in human T-cell leukemia virus type 1 proviral genomes in vivo. J. Virol. 2010, 84, 7278–7287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazato, P.; Matsuo, M.; Katsuya, H.; Satou, Y. Transcriptional and Epigenetic Regulatory Mechanisms Affecting HTLV-1 Provirus. Viruses 2016, 8, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillet, N.A.; Malani, N.; Melamed, A.; Gormley, N.; Carter, R.; Bentley, D.; Berry, C.; Bushman, F.D.; Taylor, G.P.; Bangham, C.R.M. The host genomic environment of the provirus determines the abundance of HTLV-1-infected T-cell clones. Blood 2011, 117, 3113–3122. [Google Scholar] [CrossRef] [PubMed]
- Koyanagi, Y.; Itoyama, Y.; Nakamura, N.; Takamatsu, K.; Kira, J.; Iwamasa, T.; Goto, I.; Yamamoto, N. In vivo infection of human T-cell leukemia virus type I in non-T cells. Virology 1993, 196, 25–33. [Google Scholar] [CrossRef]
- Lee, T.-H.; Chafets, D.M.; Busch, M.P.; Murphy, E.L. Quantitation of HTLV-I and II proviral load using real-time quantitative PCR with SYBR Green chemistry. J. Clin. Virol. 2004, 31, 275–282. [Google Scholar] [CrossRef]
- Murphy, E.L. Infection with human T-lymphotropic virus types-1 and -2 (HTLV-1 and -2): Implications for blood transfusion safety. Transfus. Clin. Biol. 2016, 23, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Montanheiro, P.A.; Penalva de Oliveira, A.C.; Posada-Vergara, M.P.; Milagres, A.C.; Tauil, C.; Marchiori, P.E.; Duarte, A.J.S.; Casseb, J. Human T-cell lymphotropic virus type I (HTLV-I) proviral DNA viral load among asymptomatic patients and patients with HTLV-I-associated myelopathy/tropical spastic paraparesis. Braz. J. Med. Biol. Res. 2005, 38, 1643–1647. [Google Scholar] [CrossRef] [Green Version]
- Kwok, S.; Kellogg, D.; Ehrlich, G.; Poiesz, B.; Bhagavati, S.; Sninsky, J.J. Characterization of a sequence of human T cell leukemia virus type I from a patient with chronic progressive myelopathy. J. Infect. Dis. 1988, 158, 1193–1197. [Google Scholar] [CrossRef]
- Quyen, T.L.; Ngo, T.A.; Bang, D.D.; Madsen, M.; Wolff, A. Classification of Multiple DNA Dyes Based on Inhibition Effects on Real-Time Loop-Mediated Isothermal Amplification (LAMP): Prospect for Point of Care Setting. Front. Microbiol. 2019, 10, 2234. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Primers 1 | Sequence (5′→3′) 2 | Positions 2,3 |
|---|---|---|---|
| HTLV-1 | FIP-1 | 5′-tgccaggctgttagcgtgacgaagactgtttgcccacca-3′ | (F1c: 8012–7993) (F2: 7948–7966) |
| BIP-1 | 5′-aacggcctccttccgttccacgtgccatcggtaaatgtcc-3′ | (B1c: 8014–8033) (B2: 8082–8063) | |
| F3-1 | 5′-caggggccctaataattctacc-3′ | 7925–7946 | |
| B3-1 | 5′-agggcccggaaatcatagg-3′ | 8101–8083 | |
| LF-1 | 5′-cctagcaggctggaaaaggg-3′ | 7986–7967 | |
| LB-1 | 5′-ctcaaccctcaccactcca-3′ | 8034–8052 | |
| HTLV-2 | FIP-2 | 5′-gatacagggagccctcacgcacacaggggcagtcatag-3′ | (F1c: 7887–7869) (F2: 7810–7828) |
| BIP-2 | 5′-gcctggtgtacaggacttctcccgtcattgaaggtccat-3′ | (B1c: 7894–7914) (B2: 7967–7950) | |
| F3-2 | 5′-tcaccaaggtgcctctaa-3′ | 7757–7774 | |
| B3-2 | 5′-gcaagggccggaaatcat-3′ | 7992–7975 | |
| LF-2 | 5′-ggttggaacattgtggtgg-3′ | 7868–7850 | |
| LB-2 | 5′-tccatcttaacaaccccagg-3′ | 7924–7943 |
| Groups | LAMP | qPCR | PCR-RFLP | ||||
|---|---|---|---|---|---|---|---|
| n | Accuracy | Sensitivity | Accuracy | Sensitivity | Accuracy | Sensitivity | |
| Total 1 | 226 | 80.5 | 80.1 | 75.7 | 75.1 | 84.9 | 84.6 |
| (74.8–85.5) | (74.2–85.2) | (69.5–81.1) | (68.9–80.7) | (79.5–89.3) | (79.1–90.0) | ||
| HTLV-1 | 164 | 92.0 | 89.0 | 88.1 | 83.5 | 94.7 | 93.9 |
| (87.7–95.2) | (83.2–93.4) | (83.1–92.0) | (77.0–88.9) | (90.9–97.2) | (89.0–97.0) | ||
| HTLV-2 | 54 | 89.8 | 57.4 | 88.9 | 53.7 | 89.8 | 57.4 |
| (85.1–93.4) | (43.2–70.8) | (84.1–92.7) | (40.0–67.4) | (85.1–93.4) | (43.2–70.8) | ||
| HTLV-1/2 mono-infected | 144 | 88.9 | 88.9 | 79.9 | 79.9 | 93.1 | 93.1 |
| (82.6–93.5) | (82.6–93.5) | (72.4–86.1) | (72.4–86.1) | (87.6–96.6) | (87.6–96.6) | ||
| HIV-coinfected | 82 | 65.9 | 63.6 | 68.3 | 66.2 | 70.4 | 68.4 |
| (54.6–76.0) | (51.9–74.3) | (57.1–78.1) | (54.6–76.6) | (59.2–80.0) | (56.8–78.6) | ||
| Cochran’s Q-Test | Post-Hoc McNemar | |
|---|---|---|
| All patients (n = 226) | ||
| LAMP × PCR-RFLP | 14.4 (<0.001) | 2.8 (0.284) |
| LAMP × qPCR | 4.5 (0.103) | |
| PCR-RFLP × qPCR | 10.5 (0.004) | |
| HTLV-1/2 mono-infected (n = 144) | ||
| LAMP × PCR-RFLP | 21.8 (<0.001) | 2.0 (0.471) |
| LAMP × qPCR | 11.3 (0.003) | |
| PCR-RFLP × qPCR | 19 (<0.001) | |
| HIV-coinfected (n = 82) | ||
| LAMP × PCR-RFLP | 1.2 (0.549) | N.A. |
| LAMP × qPCR | N.A. | |
| PCR-RFLP × qPCR | N.A. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes, Y.; Caterino-de-Araujo, A.; Campos, K.; Gonçalves, M.G.; Leite, A.C.; Lima, M.A.; Araújo, A.; Silva, M.T.; Espíndola, O. Loop-Mediated Isothermal Amplification (LAMP) Assay for Rapid and Accurate Confirmatory Diagnosis of HTLV-1/2 Infection. Viruses 2020, 12, 981. https://0-doi-org.brum.beds.ac.uk/10.3390/v12090981
Gomes Y, Caterino-de-Araujo A, Campos K, Gonçalves MG, Leite AC, Lima MA, Araújo A, Silva MT, Espíndola O. Loop-Mediated Isothermal Amplification (LAMP) Assay for Rapid and Accurate Confirmatory Diagnosis of HTLV-1/2 Infection. Viruses. 2020; 12(9):981. https://0-doi-org.brum.beds.ac.uk/10.3390/v12090981
Chicago/Turabian StyleGomes, Yago, Adele Caterino-de-Araujo, Karoline Campos, Maria Gisele Gonçalves, Ana Claudia Leite, Marco Antonio Lima, Abelardo Araújo, Marcus Tulius Silva, and Otávio Espíndola. 2020. "Loop-Mediated Isothermal Amplification (LAMP) Assay for Rapid and Accurate Confirmatory Diagnosis of HTLV-1/2 Infection" Viruses 12, no. 9: 981. https://0-doi-org.brum.beds.ac.uk/10.3390/v12090981