
The Holin-Endolysin Lysis System of the OP2-Like Phage X2 Infecting Xanthomonas oryzae pv. oryzae


 , ,
, ,
Abstract
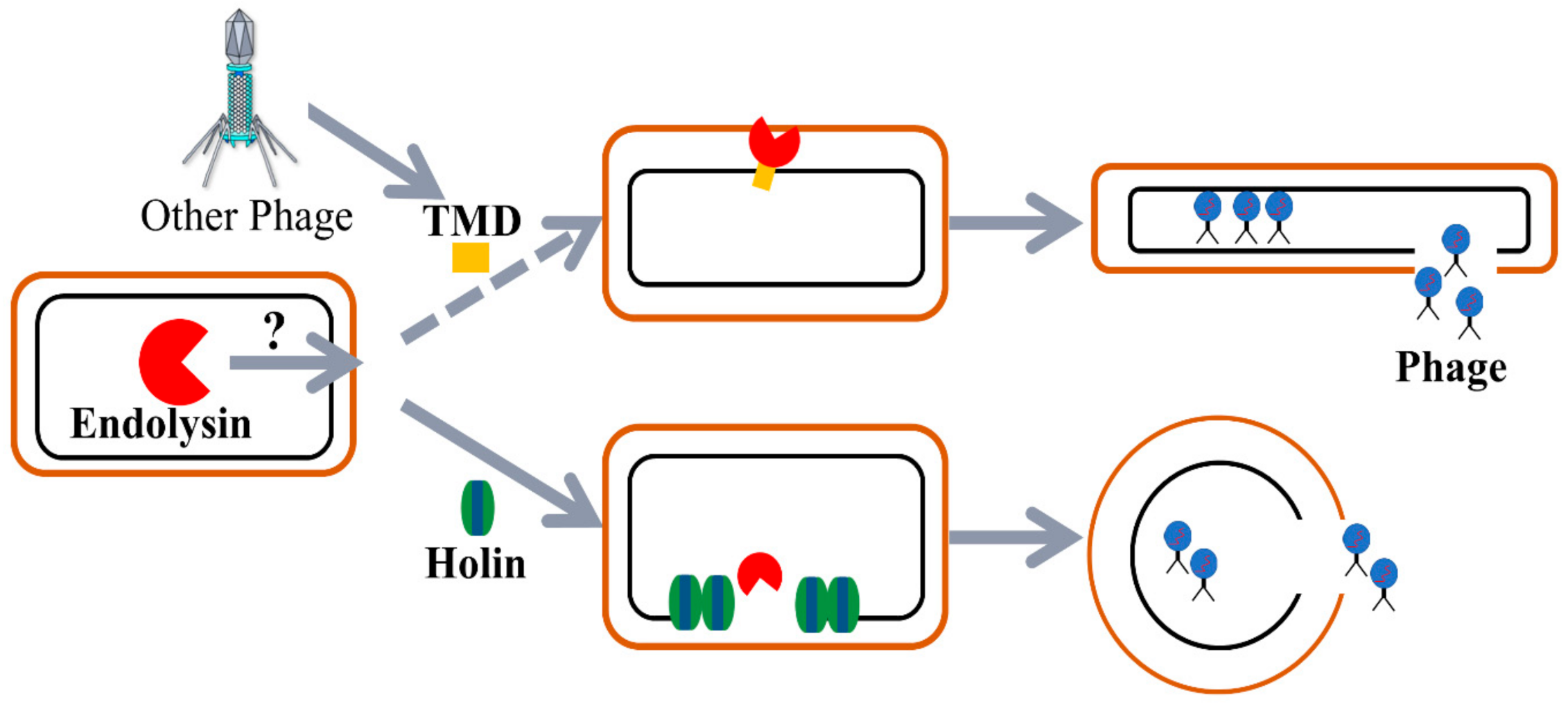
:1. Introduction
2. Materials and Methods
2.1. Bacteria, Plasmids and Growth Conditions
2.2. Genome Sequencing and Phylogenetic Analysis of Phage X2
2.3. In Silico Genome-Wide Analysis of Lysis System
2.4. Construction of Recombinant Plasmids
2.5. Growth Measurement
2.6. Live/Dead Cell Staining and Flow Cytometry Observation
2.7. Detection of β-Galactosidase Activity and Efflux of the Nucleic Acid
2.8. Transmission Electron Microscopic (TEM) Observation
2.9. Purification and Antibacterial Effect of X2-Lys
2.10. Extraction and Detection of Membrane Proteins
2.11. Statistical Analysis
3. Results and Discussion
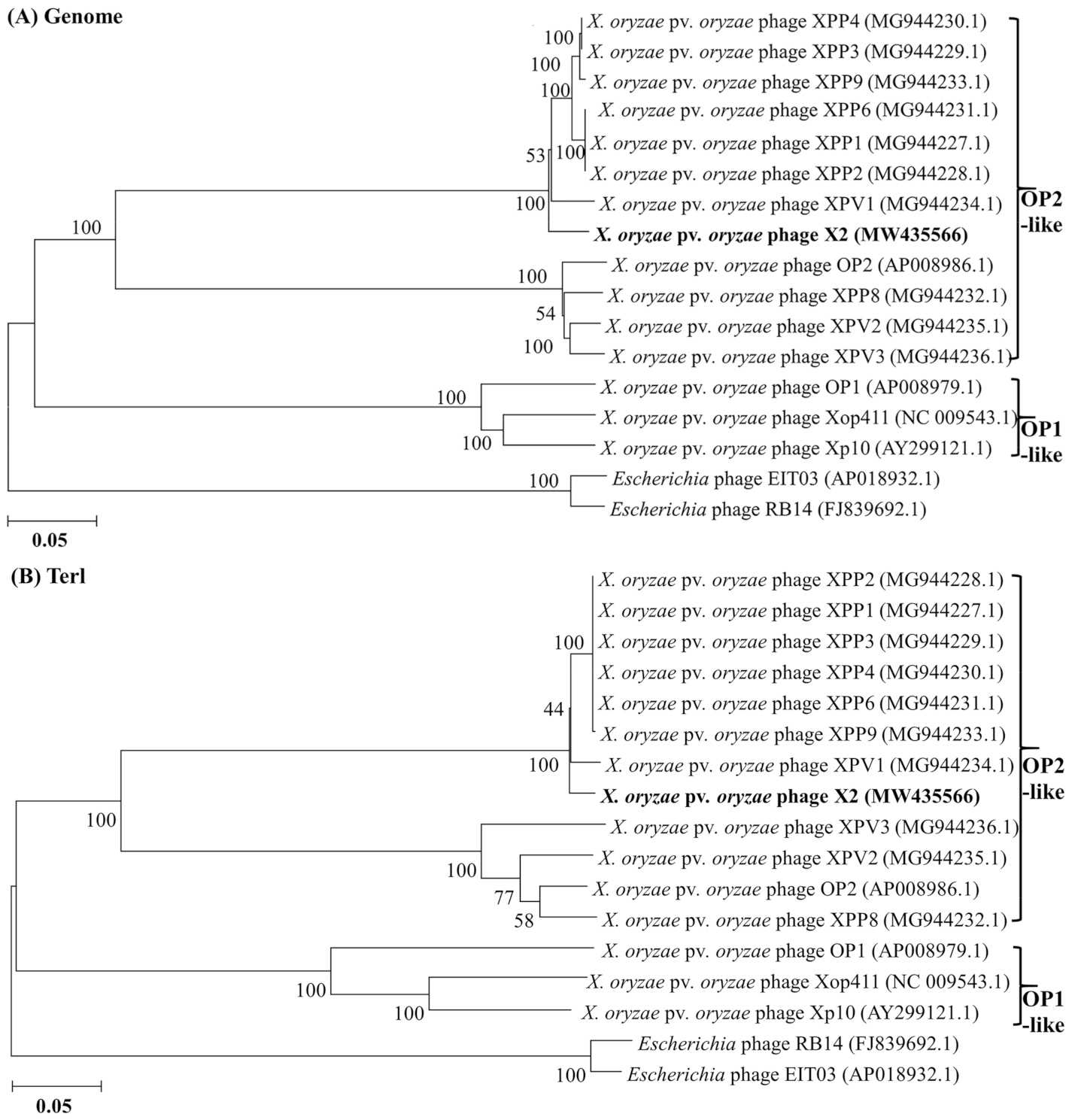
3.1. Analysis of Phage X2 Genome
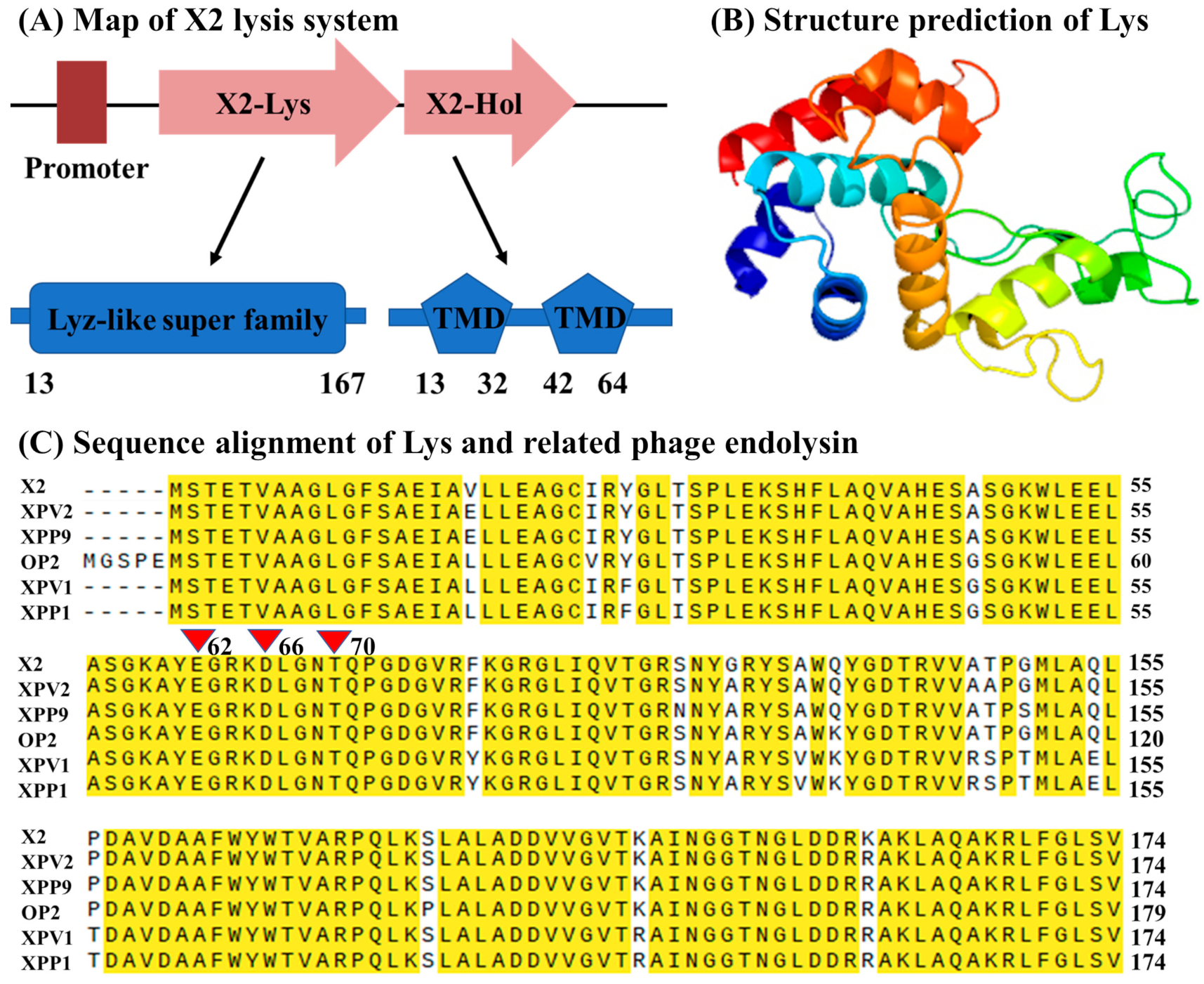
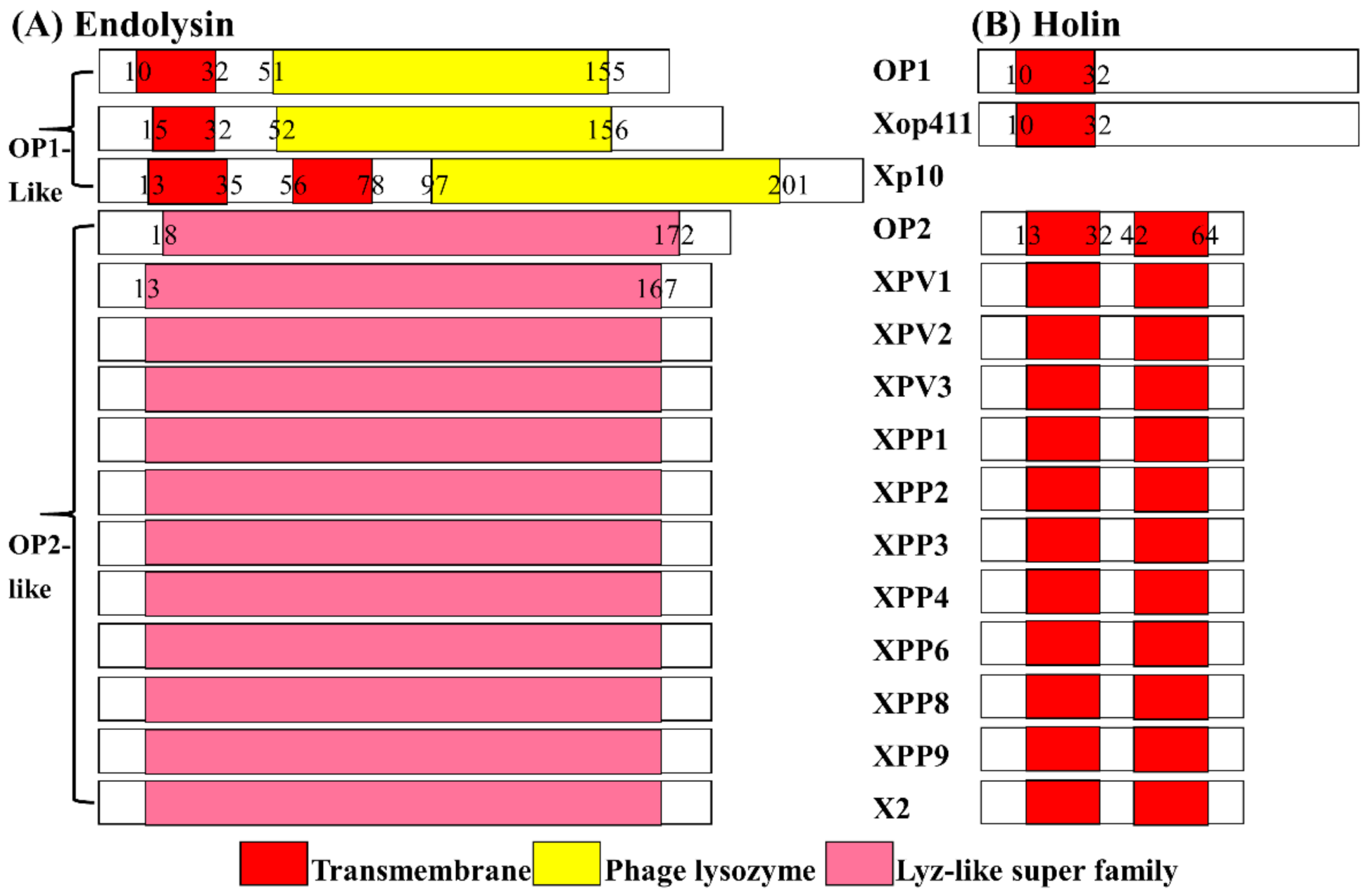
3.2. In Silico Identification of Phage X2 Lysis System
3.3. Phage X2 Lysis System Similar to OP2- Other Than OP1-Like Phages
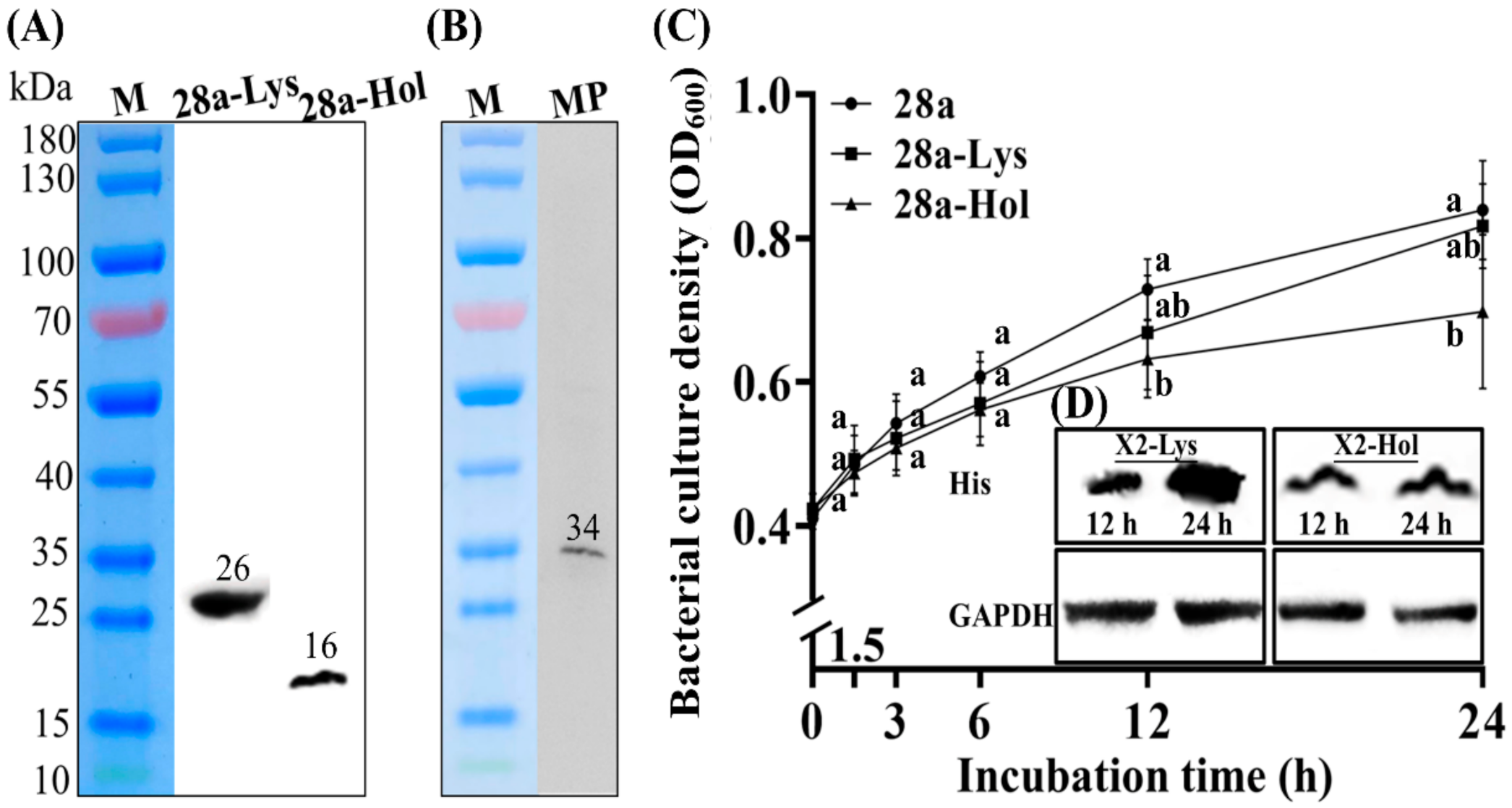
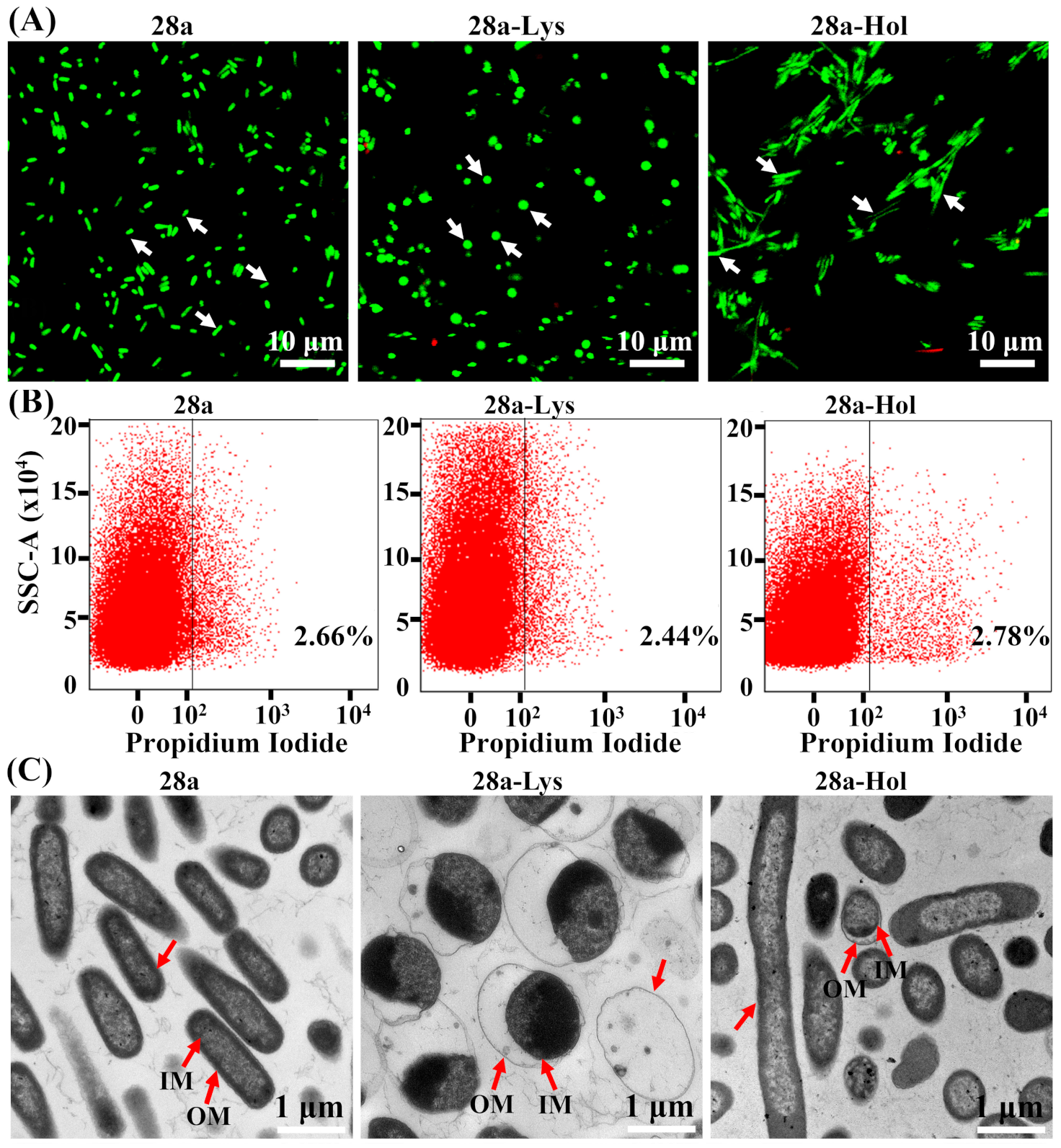
3.4. Subcellular Localization and Antibacterial Activity of X2-Lys and X2-Hol
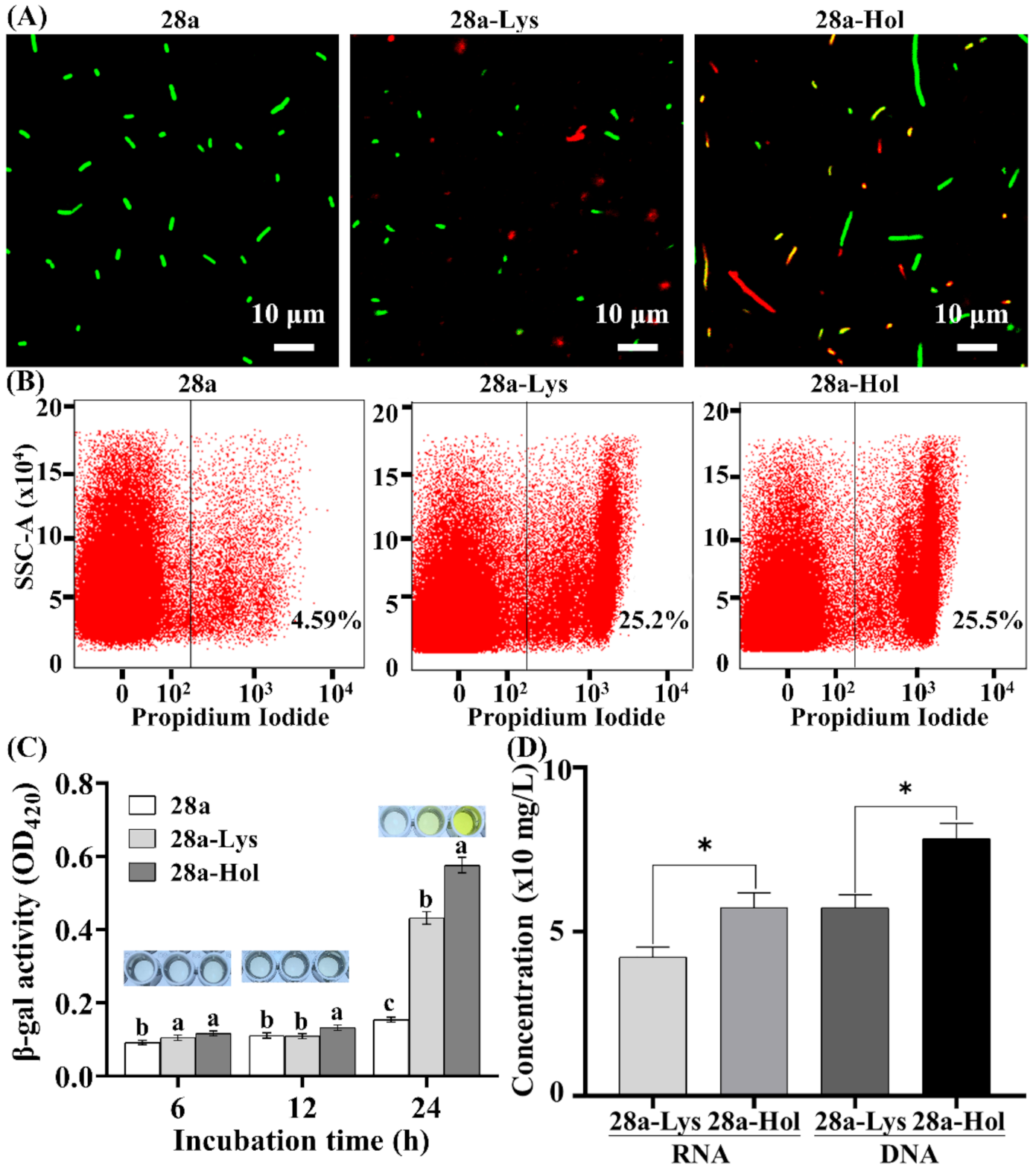
3.5. Changes in Bacterial Morphology Induced by X2-Lys and X2-Hol Expression
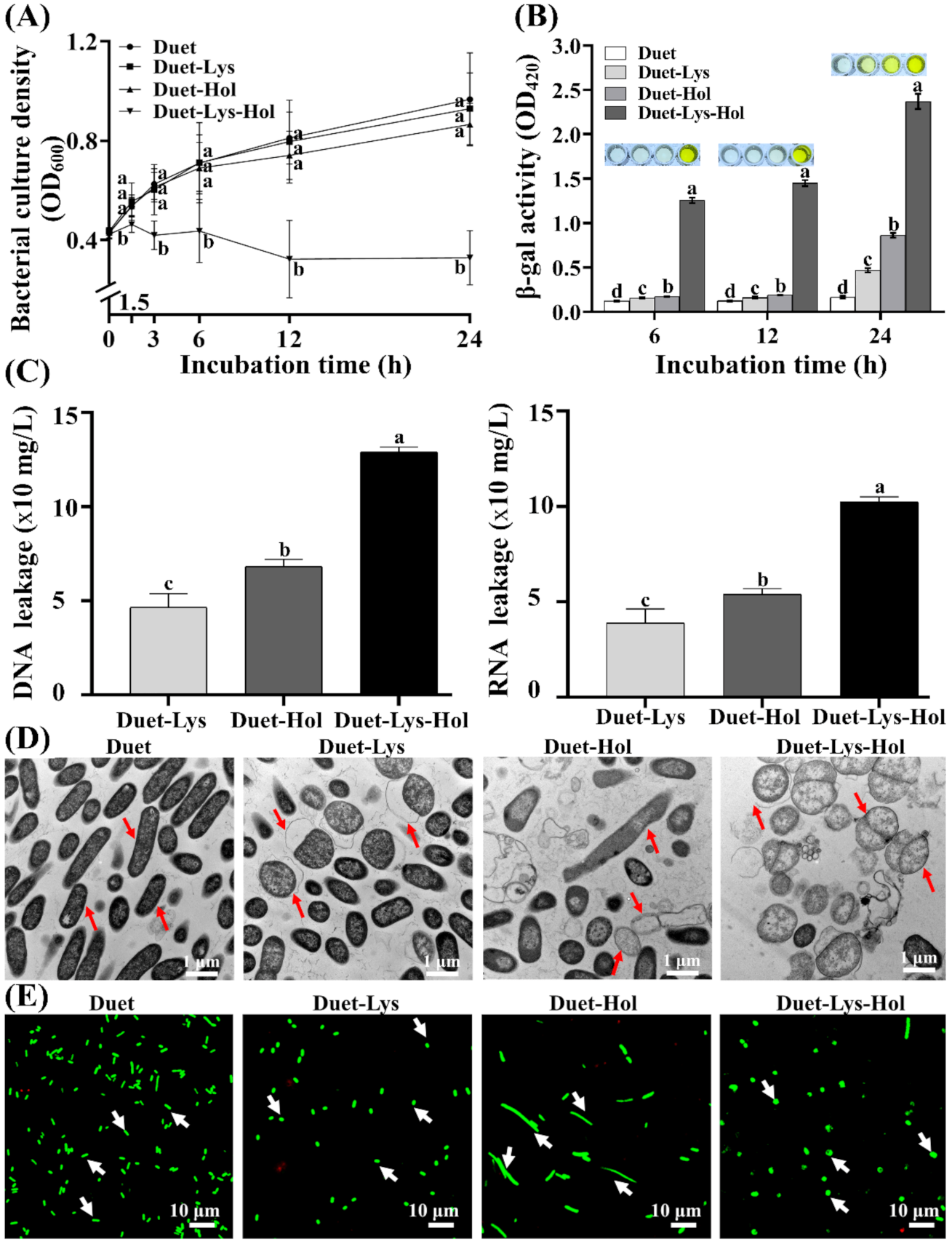
3.6. Effects of Co-Expression of X2-Hol and X2-Lys on Cells
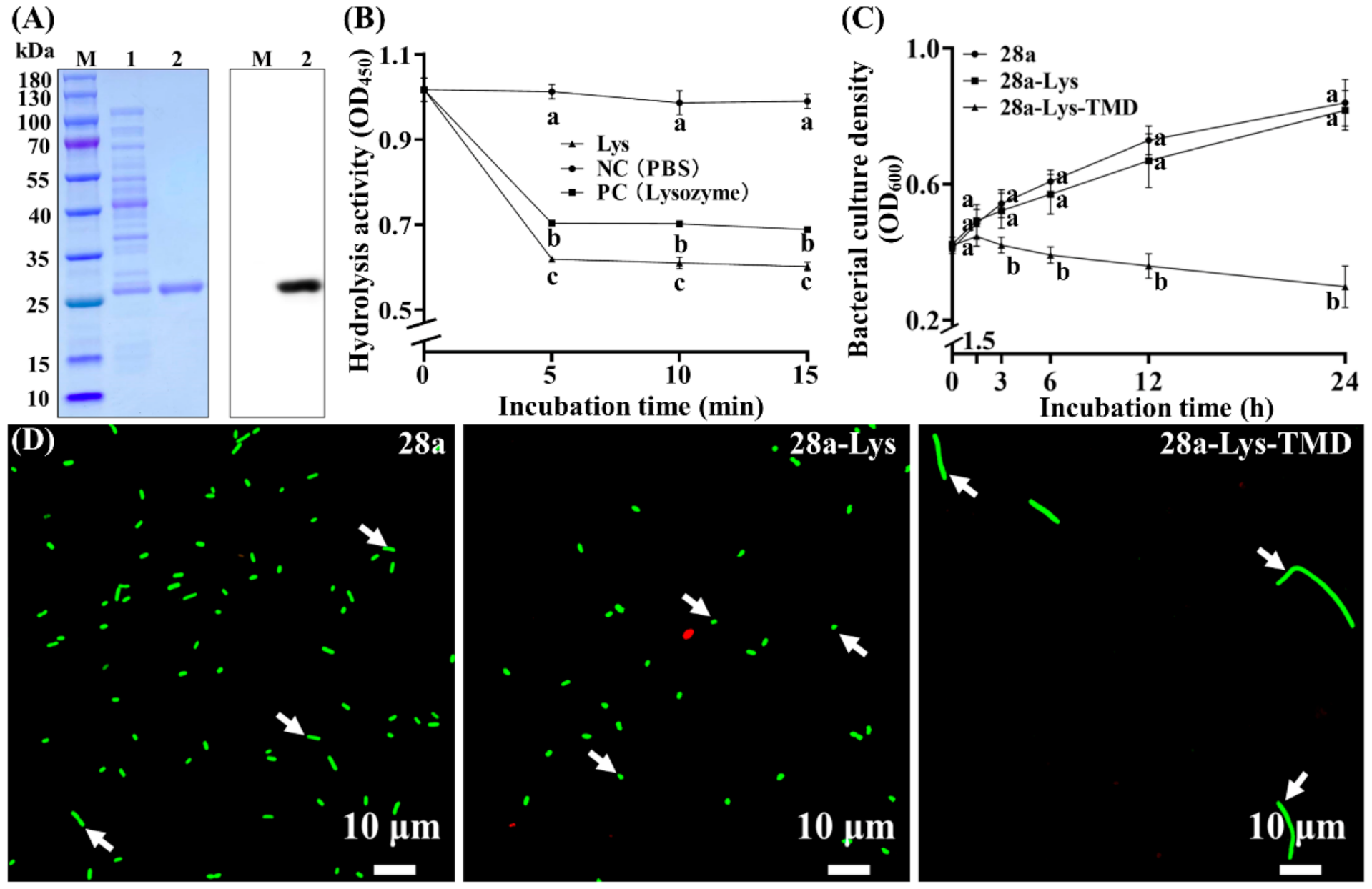
3.7. Purified X2-Lys Shows Antibacterial Activity
3.8. Potential Multiple Roles of the Transmembrane Domain in X2-Lys for Transport and Division Inhibition
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kronheim, S.; Daniel-Ivad, M.; Duan, Z.; Hwang, S.; Wong, A.I.; Mantel, I.; Nodwell, J.R.; Maxwell, K.L. A chemical defence against phage infection. Nature 2018, 564, 283–286. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.N.; Lin, J.W.; Chow, T.Y.; Tseng, Y.H.; Weng, S.F. A novel lysozyme from Xanthomonas oryzae phage varphiXo411 active against Xanthomonas and Stenotrophomonas. Protein Expr. Purif. 2006, 50, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Lim, J.A.; Kong, M.; Ryu, S.; Rhee, S. Structure of bacteriophage SPN1S endolysin reveals an unusual two-module fold for the peptidoglycan lytic and binding activity. Mol. Microbiol. 2014, 92, 316–325. [Google Scholar] [CrossRef]
- Bai, J.; Lee, S.; Ryu, S. Identification and in vitro Characterization of a Novel Phage Endolysin that Targets Gram-Negative Bacteria. Microorganisms 2020, 8, 447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Y.; Li, X.; Wang, L.; Li, G.; Cong, C.; Li, R.; Cui, H.; Murtaza, B.; Xu, Y. The endolysin of the Acinetobacter baumannii phage vB_AbaP_D2 shows broad antibacterial activity. Microb. Biotechnol. 2021, 14, 403–418. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, T.; Molnar, J.; Varga, I.; Nagy, I.K.; Valappil, S.K.; Papp, S.; Vera Cruz, C.M.; Oliva, R.; Vizi, T.; Schneider, G.; et al. Complete Genome Sequences of 10 Xanthomonas oryzae pv. oryzae Bacteriophages. Microbiol. Resour. Announc. 2019, 8, e00334-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buttimer, C.; McAuliffe, O.; Ross, R.P.; Hill, C.; O’Mahony, J.; Coffey, A. Bacteriophages and Bacterial Plant Diseases. Front. Microbiol. 2017, 8, 34. [Google Scholar] [CrossRef] [Green Version]
- Chang, R.Y.K.; Wallin, M.; Lin, Y.; Leung, S.S.Y.; Wang, H.; Morales, S.; Chan, H.K. Phage therapy for respiratory infections. Adv. Drug Deliv. Rev. 2018, 133, 76–86. [Google Scholar] [CrossRef]
- Muturi, P.; Yu, J.; Maina, A.N.; Kariuki, S.; Mwaura, F.B.; Wei, H. Bacteriophages Isolated in China for the Control of Pectobacterium carotovorum Causing Potato Soft Rot in Kenya. Virol. Sin. 2019, 34, 287–294. [Google Scholar] [CrossRef]
- Nachimuthu, R.; Royam, M.M.; Manohar, P.; Leptihn, S. Application of bacteriophages and endolysins in aquacultures a biocontrol measure. Biol. Control. 2021, 160, 104678. [Google Scholar] [CrossRef]
- Nelson, D.; Loomis, L.; Fischetti, V.A. Prevention and elimination of upper respiratory colonization of mice by group A streptococci by using a bacteriophage lytic enzyme. Proc. Natl. Acad. Sci. USA 2001, 98, 4107–4112. [Google Scholar] [CrossRef] [Green Version]
- Cha, Y.; Son, B.; Ryu, S. Effective removal of staphylococcal biofilms on various food contact surfaces by Staphylococcus aureus phage endolysin LysCSA13. Food Microbiol. 2019, 84, 103245. [Google Scholar] [CrossRef]
- Wang, I.N.; Smith, D.L.; Young, R. Holins: The protein clocks of bacteriophage infections. Annu. Rev. Microbiol. 2000, 54, 799–825. [Google Scholar] [CrossRef]
- Catalao, M.J.; Gil, F.; Moniz-Pereira, J.; Sao-Jose, C.; Pimentel, M. Diversity in bacterial lysis systems: Bacteriophages show the way. FEMS Microbiol. Rev. 2013, 37, 554–571. [Google Scholar] [CrossRef] [Green Version]
- Young, R. Phage lysis: Do we have the hole story yet? Curr. Opin. Microbiol. 2013, 16, 790–797. [Google Scholar] [CrossRef] [Green Version]
- White, R.; Chiba, S.; Pang, T.; Dewey, J.S.; Savva, C.G.; Holzenburg, A.; Pogliano, K.; Young, R. Holin triggering in real time. Proc. Natl. Acad. Sci. USA 2011, 108, 798–803. [Google Scholar] [CrossRef] [Green Version]
- Sao-Jose, C.; Parreira, R.; Vieira, G.; Santos, M.A. The N-terminal region of the Oenococcus oeni bacteriophage fOg44 lysin behaves as a bona fide signal peptide in Escherichia coli and as a cis-inhibitory element, preventing lytic activity on oenococcal cells. J. Bacteriol. 2000, 182, 5823–5831. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Struck, D.K.; Deaton, J.; Wang, I.N.; Young, R. A signal-arrest-release sequence mediates export and control of the phage P1 endolysin. Proc. Natl. Acad. Sci. USA 2004, 101, 6415–6420. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Arulandu, A.; Struck, D.K.; Swanson, S.; Sacchettini, J.C.; Young, R. Disulfide isomerization after membrane release of its SAR domain activates P1 lysozyme. Science 2005, 307, 113–117. [Google Scholar] [CrossRef]
- Briers, Y.; Peeters, L.M.; Volckaert, G.; Lavigne, R. The lysis cassette of bacteriophage varphiKMV encodes a signal-arrest-release endolysin and a pinholin. Bacteriophage 2011, 1, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Chen, J. Research on Identification and Function of a New Type of Holin-Endolysin Lysis system in Acidovorax Oryzae Phage AP1. Master’s Thesis, Hangzhou Normal University, Hangzhou, China, 2019. [Google Scholar]
- Wang, L. Research on isolation, identification and characterization of phages of two main pathogenic bacteria in rice. Master’s Thesis, Hangzhou Normal University, Hangzhou, China, 2016. [Google Scholar]
- Ogunyemi, S.O.; Chen, J.; Zhang, M.C.; Wang, L.; Masum, M.M.I.; Yan, C.Q.; An, Q.L.; Li, B.; Chen, J.P. Identification and characterization of five new OP2-related Myoviridae bacteriophages infecting different strains of Xanthomonas oryzae pv. oryzae. J. Plant Pathol. 2019, 101, 263–273. [Google Scholar] [CrossRef]
- Shi, J.; Ma, X.; Gao, Y.; Fan, D.; Zhu, C.; Mi, Y.; Xue, W. Hydroxylation of human type III collagen alpha chain by recombinant coexpression with a viral prolyl 4-hydroxylase in Escherichia coli. Protein J. 2017, 36, 322–331. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, F.; Li, B.; Yang, Y.Z.; Ibrahim, M.; Fang, Y.S.; Qiu, W.; Masum, M.M.I.; Oliva, R. Characterization and functional analysis of clpB gene from Acidovorax avenae subsp avenae RS-1. Plant Pathol. 2017, 66, 1369–1379. [Google Scholar] [CrossRef]
- Masum, M.M.I.; Siddiqa, M.M.; Ali, K.A.; Zhang, Y.; Abdallah, Y.; Ibrahim, E.; Qiu, W.; Yan, C.Q.; Li, B. Biogenic Synthesis of Silver Nanoparticles Using Phyllanthus emblica Fruit Extract and Its Inhibitory Action Against the Pathogen Acidovorax oryzae Strain RS-2 of Rice Bacterial Brown Stripe. Front. Microbiol. 2019, 10, 820. [Google Scholar] [CrossRef]
- Wu, J.; Abbas, H.M.K.; Li, J.; Yuan, Y.; Liu, Y.; Wang, G.; Dong, A.W. Cell Membrane-Interrupting Antimicrobial Peptides from Isatis indigotica Fortune Isolated by a Bacillus subtilis Expression System. Biomolecules 2019, 10, 30. [Google Scholar] [CrossRef] [Green Version]
- Abdallah, Y.; Liu, M.J.; Ogunyemi, S.O.; Ahmed, T.; Fouad, H.; Abdelazez, A.; Yan, C.Q.; Yang, Y.; Chen, J.P.; Li, B. Bioinspired Green Synthesis of Chitosan and Zinc Oxide Nanoparticles with Strong Antibacterial Activity against Rice Pathogen Xanthomonas oryzae pv. oryzae. Molecules 2020, 25, 4795. [Google Scholar] [CrossRef]
- Mikoulinskaia, G.V.; Odinokova, I.V.; Zimin, A.A.; Lysanskaya, V.Y.; Feofanov, S.A.; Stepnaya, O.A. Identification and characterization of the metal ion-dependent L-alanoyl-D-glutamate peptidase encoded by bacteriophage T5. FEBS J. 2009, 276, 7329–7342. [Google Scholar] [CrossRef]
- Lim, J.A.; Shin, H.; Kang, D.H.; Ryu, S. Characterization of endolysin from a Salmonella Typhimurium-infecting bacteriophage SPN1S. Res. Microbiol. 2012, 163, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.W.; Gao, C.; Jiang, Y.; Wang, M.; Zhou, X.H.; Shao, H.B.; Gong, Z.; McMinn, A. Metagenomic Characterization of the Viral Community of the South Scotia Ridge. Viruses 2019, 11, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.R.; Liu, Y.D.; Wang, M.; Wang, M.W.; Jiang, T.; Sun, J.H.; Gao, C.; Jiang, Y.; Guo, C.; Shao, H.B.; et al. Characterization and Genome Analysis of a Novel Marine Alteromonas Phage P24. Curr. Microbiol. 2020, 77, 2813–2820. [Google Scholar] [CrossRef] [PubMed]
- Lavigne, R.; Darius, P.; Summer, E.J.; Seto, D.; Mahadevan, P.; Nilsson, A.S.; Ackermann, H.W.; Kropinski, A.M. Classification of Myoviridae bacteriophages using protein sequence similarity. BMC Microbiol. 2009, 9, 224. [Google Scholar] [CrossRef] [Green Version]
- Adams, M.J.; Carstens, E.B. Ratification vote on taxonomic proposals to the International Committee on Taxonomy of Viruses (2012). Arch. Virol. 2012, 157, 1411–1422. [Google Scholar] [CrossRef] [Green Version]
- Yuzenkova, J.; Nechaev, S.; Berlin, J.; Rogulja, D.; Kuznedelov, K.; Inman, R.; Mushegian, A.; Severinov, K. Genome of Xanthomonas oryzae bacteriophage Xp10: An odd T-odd phage. J. Mol. Biol. 2003, 330, 735–748. [Google Scholar] [CrossRef]
- Weng, S.F.; Fu, Y.C.; Lin, J.W.; Tseng, T.T. Identification of a Broad-Spectrum Peptidoglycan Hydrolase Associated with the Particle of Xanthomonas oryzae Phage Xop411. J. Mol. Microbiol. Biotechnol. 2018, 28, 78–86. [Google Scholar] [CrossRef]
- Reddy, B.L.; Saier, M.H. Topological and phylogenetic analyses of bacterial holin families and superfamilies. BBA-Biomembr. 2013, 1828, 2654–2671. [Google Scholar] [CrossRef] [Green Version]
- Young, R. Phage Lysis: Three Steps, Three Choices, One Outcome. J. Microbiol. 2014, 52, 243–258. [Google Scholar] [CrossRef]
- To, K.H.; Dewey, J.; Weaver, J.; Park, T.; Young, R. Functional Analysis of a Class I Holin, P2 Y. J. Bacteriol. 2013, 195, 1346–1355. [Google Scholar] [CrossRef] [Green Version]
- Kuroki, R.; Weaver, L.H.; Matthews, B.W. Structural basis of the conversion of T4 lysozyme into a transglycosidase by reengineering the active site. Proc. Natl. Acad. Sci. USA 1999, 96, 8949–8954. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, H.; Melo, L.D.; Santos, S.B.; Nobrega, F.L.; Ferreira, E.C.; Cerca, N.; Azeredo, J.; Kluskens, L.D. Molecular aspects and comparative genomics of bacteriophage endolysins. J. Virol. 2013, 87, 4558–4570. [Google Scholar] [CrossRef] [Green Version]
- Altrichter, S.; Haase, M.; Loh, B.; Kuhn, A.; Leptihn, S. Mechanism of the Spontaneous and Directional Membrane Insertion of a 2-Transmembrane Ion Channel. ACS Chem. Biol. 2017, 12, 380–388. [Google Scholar] [CrossRef]
- Kuhn, A.; Haase, M.; Leptihn, S. Assisted and Unassisted Protein Insertion into Liposomes. Biophys. J. 2017, 113, 1187–1193. [Google Scholar] [CrossRef] [PubMed]
- Park, T.; Struck, D.K.; Dankenbring, C.A.; Young, R. The pinholin of lambdoid phage 21: Control of lysis by membrane depolarization. J. Bacteriol. 2007, 189, 9135–9139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakimoto, S. Classification of strains of Xanthomonas oryzae on the basis of their susceptibility against bacteriophages. Jpn. J. Phytopathol. 2009, 25, 193–198. [Google Scholar] [CrossRef]
- Lee, C.N.; Hu, R.M.; Chow, T.Y.; Lin, J.W.; Chen, H.Y.; Tseng, Y.H.; Weng, S.F. Comparison of genomes of three Xanthomonas oryzae bacteriophages. BMC Genom. 2007, 8, 442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, X.P.; Guiltinan, M.J. DNA-Binding Specificity of the Wheat Bzip Protein Embp-1. Nucleic Acids Res. 1994, 22, 4969–4978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gründling, A.; Smith, D.L.; Bläsi, U.; Young, R. Dimerization between the holin and holin inhibitor of phage lambda. J. Bacteriol. 2000, 182, 6075–6081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turnbull, L.; Toyofuku, M.; Hynen, A.L.; Kurosawa, M.; Pessi, G.; Petty, N.K.; Osvath, S.R.; Carcamo-Oyarce, G.; Gloag, E.S.; Shimoni, R.; et al. Explosive cell lysis as a mechanism for the biogenesis of bacterial membrane vesicles and biofilms. Nat. Commun. 2016, 7, 11220. [Google Scholar] [CrossRef] [Green Version]
- Attai, H.; Rimbey, J.; Smith, G.P.; Brown, P.J.B. Expression of a Peptidoglycan Hydrolase from Lytic Bacteriophages Atu_ph02 and Atu_ph03 Triggers Lysis of Agrobacterium tumefaciens. Appl. Environ. Microb. 2017, 83, e01498-17. [Google Scholar] [CrossRef] [Green Version]
- Ning, H.Q.; Lin, H.; Wang, J.X. Synergistic effects of endolysin Lysqdvp001 and epsilon-poly-lysine in controlling Vibrio parahaemolyticus and its biofilms. Int. J. Food Microbiol. 2021, 343, 109112. [Google Scholar] [CrossRef]
- Adamczyk-Poplawska, M.; Tracz-Gaszewska, Z.; Lasota, P.; Kwiatek, A.; Piekarowicz, A. Haemophilus influenzae HP1 Bacteriophage Encodes a Lytic Cassette with a Pinholin and a Signal-Arrest-Release Endolysin. Int. J. Mol. Sci. 2020, 21, 4013. [Google Scholar] [CrossRef]
- Dewey, J.S.; Savva, C.G.; White, R.L.; Vitha, S.; Holzenburg, A.; Young, R. Micron-scale holes terminate the phage infection cycle. Proc. Natl. Acad. Sci. USA 2010, 107, 2219–2223. [Google Scholar] [CrossRef] [Green Version]
- Hermoso, J.A.; García, J.L.; García, P. Taking aim on bacterial pathogens: From phage therapy to enzybiotics. Curr. Opin. Microbiol. 2007, 10, 461–472. [Google Scholar] [CrossRef]
- Fischetti, V.A. Bacteriophage lytic enzymes: Novel anti-infectives. Trends Microbiol. 2005, 13, 491–496. [Google Scholar] [CrossRef]
- Son, B.; Kong, M.; Lee, Y.; Ryu, S. Development of a Novel Chimeric Endolysin, Lys109 With Enhanced Lytic Activity Against Staphylococcus aureus. Front. Microbiol. 2020, 11, 615887. [Google Scholar] [CrossRef]
- Lu, Y.F.; Wang, Y.R.; Wang, J.; Zhao, Y.; Zhong, Q.; Li, G.; Fu, Z.F.; Lu, S.G. Phage Endolysin LysP108 Showed Promising Antibacterial Potential Against Methicillin-resistant Staphylococcus aureus. Front. Cell Infect. Microbiol. 2021, 11, 668430. [Google Scholar] [CrossRef]
- Briers, Y.; Walmagh, M.; Van Puyenbroeck, V.; Cornelissen, A.; Cenens, W.; Aertsen, A.; Oliveira, H.; Azeredo, J.; Verween, G.; Pirnay, J.P.; et al. Engineered endolysin-based "Artilysins" to combat multidrug-resistant gram-negative pathogens. mBio 2014, 5, e01379-01314. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Wang, M.; Yu, J.; Wei, H. Antibacterial Activity of a Novel Peptide-Modified Lysin Against Acinetobacter baumannii and Pseudomonas aeruginosa. Front. Microbiol. 2015, 6, 1471. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Feng, C.; Ren, J.; Zhuang, X.; Zhang, Y.; Zhu, Y.; Dong, K.; He, P.; Guo, X.; Qin, J. A Novel Antimicrobial Endolysin, LysPA26, against Pseudomonas aeruginosa. Front. Microbiol. 2017, 8, 293. [Google Scholar] [CrossRef] [Green Version]
- Alakomi, H.L.; Paananen, A.; Suihko, M.L.; Helander, I.M.; Saarela, M. Weakening effect of cell permeabilizers on gram-negative bacteria causing biodeterioration. Appl. Environ. Microb. 2006, 72, 4695–4703. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasmids | Description | Sources |
|---|---|---|
| pET28a | KmR; cloning vector | Novagen |
| pET28a-Lys | KmR; recombinant expression vector with a X2-Lys | This study |
| pET28a-Hol | KmR; recombinant expression vector with a X2-Hol | This study |
| pET28a-Lys-TMD | KmR; recombinant expression vector with a X2-Lys-TMD | This study |
| pETDuet-1 | AmpR; cloning vector | Laboratory collection |
| pETDuet-Lys | AmpR; recombinant expression vector with a X2-Lys | This study |
| pETDuet-Hol | AmpR; recombinant expression vector with a X2-Hol | This study |
| pETDuet-Lys-Hol | AmpR; recombinant expression vector with a X2-Lys and X2-Hol | This study |
| Primer Name | Nucleotide Sequence (5′–3′) | Characterization |
|---|---|---|
| 28a-Lys-F | CGGGATCCATGTCAACTGAGACTGTCGCAG (B) | Gene of Lys from X2 phage |
| 28a-Lys-R | CCCAAGCTTTACGCTCAGTCCAAACAGGCG (H) | |
| 28a-Hol-F | CGGGATCCATGACCCCTGAACCTCGTAATATCG (B) | Gene of Hol from X2 phage |
| 28a-Hol-R | CCCAAGCTTGCCTGCATTGTTTGCGCC (H) | |
| TMD-F | CCCAAGCTTAGCCTCGGCAACTGGC (H) | Gene of TMD |
| TMD-R | CCGCTCGAGTGACCACCCCTCTCGCC (X) | |
| Duet-Lys-F | AACTGCAGATGTCAACTGAGACTGTCGCAG (P) | Gene of Lys from X2 phage |
| Duet-Lys-R | CCCAAGCTTTACGCTCAGTCCAAACAGGCG (H) | |
| Duet-Hol-F | CTATACATATGATGACCCCTGAACCTCGTAATATCG (N) | Gene of Hol from X2 phage |
| Duet-Hol-R | GAAGATCTTCAGCCTGCATTGTTTGCGCC (Bg) |
| Type | Name | Length (bp) | G+C Content (%) | Number of ORF | Accession No. |
|---|---|---|---|---|---|
| OP1- | |||||
| OP1 | 43,785 | 51 | 59 | AP008979.1 | |
| Xop411 | 44,520 | 52 | 60 | NC_009543.1 | |
| Xp10 | 44,373 | 52 | 60 | AY299121.1 | |
| OP2- | |||||
| OP2 | 46,643 | 61 | 62 | AP008986.1 | |
| XPP1 | 46,195 | 62 | 73 | MG944227 | |
| XPP2 | 46,204 | 61 | 72 | MG944228 | |
| XPP3 | 46,201 | 62 | 78 | MG944229 | |
| XPP4 | 46,200 | 62 | 73 | MG944230 | |
| XPP6 | 46,204 | 61 | 72 | MG944231 | |
| XPP8 | 46,184 | 62 | 74 | MG944232 | |
| XPP9 | 46,201 | 61 | 80 | MG944233 | |
| XPV1 | 46,503 | 60 | 77 | MG944234 | |
| XPV2 | 45,969 | 64 | 74 | MG944235 | |
| XPV3 | 47,046 | 60 | 76 | MG944236 | |
| X2 | 45,966 | 61 | 74 | MW435566 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Z.; Zhang, Y.; Xu, X.; Ahmed, T.; Yang, Y.; Loh, B.; Leptihn, S.; Yan, C.; Chen, J.; Li, B. The Holin-Endolysin Lysis System of the OP2-Like Phage X2 Infecting Xanthomonas oryzae pv. oryzae. Viruses 2021, 13, 1949. https://0-doi-org.brum.beds.ac.uk/10.3390/v13101949
Wu Z, Zhang Y, Xu X, Ahmed T, Yang Y, Loh B, Leptihn S, Yan C, Chen J, Li B. The Holin-Endolysin Lysis System of the OP2-Like Phage X2 Infecting Xanthomonas oryzae pv. oryzae. Viruses. 2021; 13(10):1949. https://0-doi-org.brum.beds.ac.uk/10.3390/v13101949
Chicago/Turabian StyleWu, Zhifeng, Yang Zhang, Xinyang Xu, Temoor Ahmed, Yong Yang, Belinda Loh, Sebastian Leptihn, Chenqi Yan, Jianping Chen, and Bin Li. 2021. "The Holin-Endolysin Lysis System of the OP2-Like Phage X2 Infecting Xanthomonas oryzae pv. oryzae" Viruses 13, no. 10: 1949. https://0-doi-org.brum.beds.ac.uk/10.3390/v13101949