
Identification of Novel Rodent-Borne Orthohantaviruses in an Endemic Area of Chronic Kidney Disease of Unknown Etiology (CKDu) in Sri Lanka

 , , , , , , , , and
, , , , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
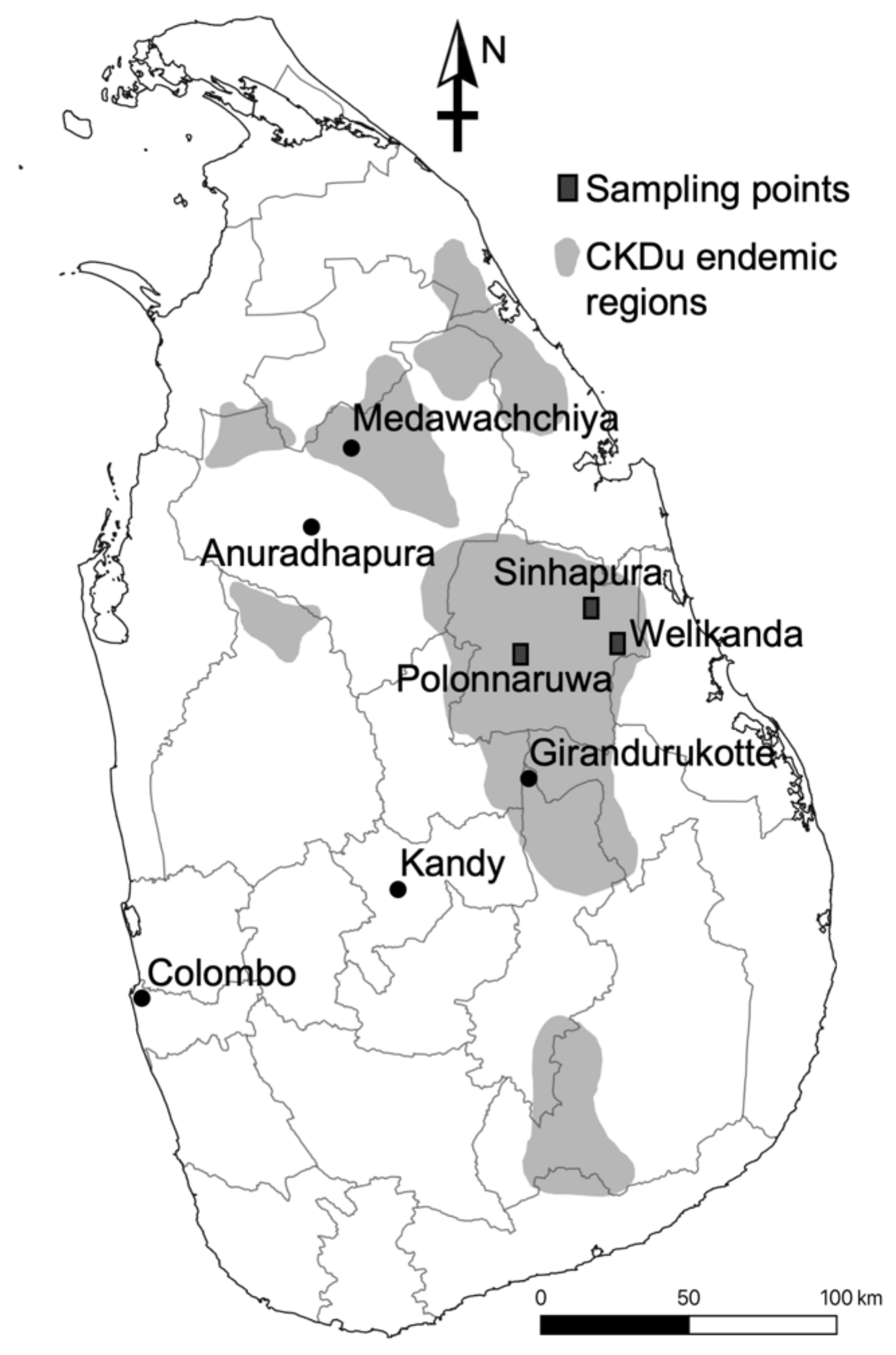
2.1. Sample Collection
2.2. DNA Extraction and Rodent Species Identification
2.3. Indirect Immunofluorescence Assay (IFA)
2.4. RNA Extraction, cDNA Synthesis, and Hantavirus Screening PCR
2.5. Genomic Sequencing
2.6. Sequence Alignment and Phylogenetic Analysis
2.7. Quantification of Viral RNA
3. Results
3.1. Animal Species Identification
3.2. Sero-Survey and Hantavirus Screening PCR
3.3. Whole-Genome Sequencing
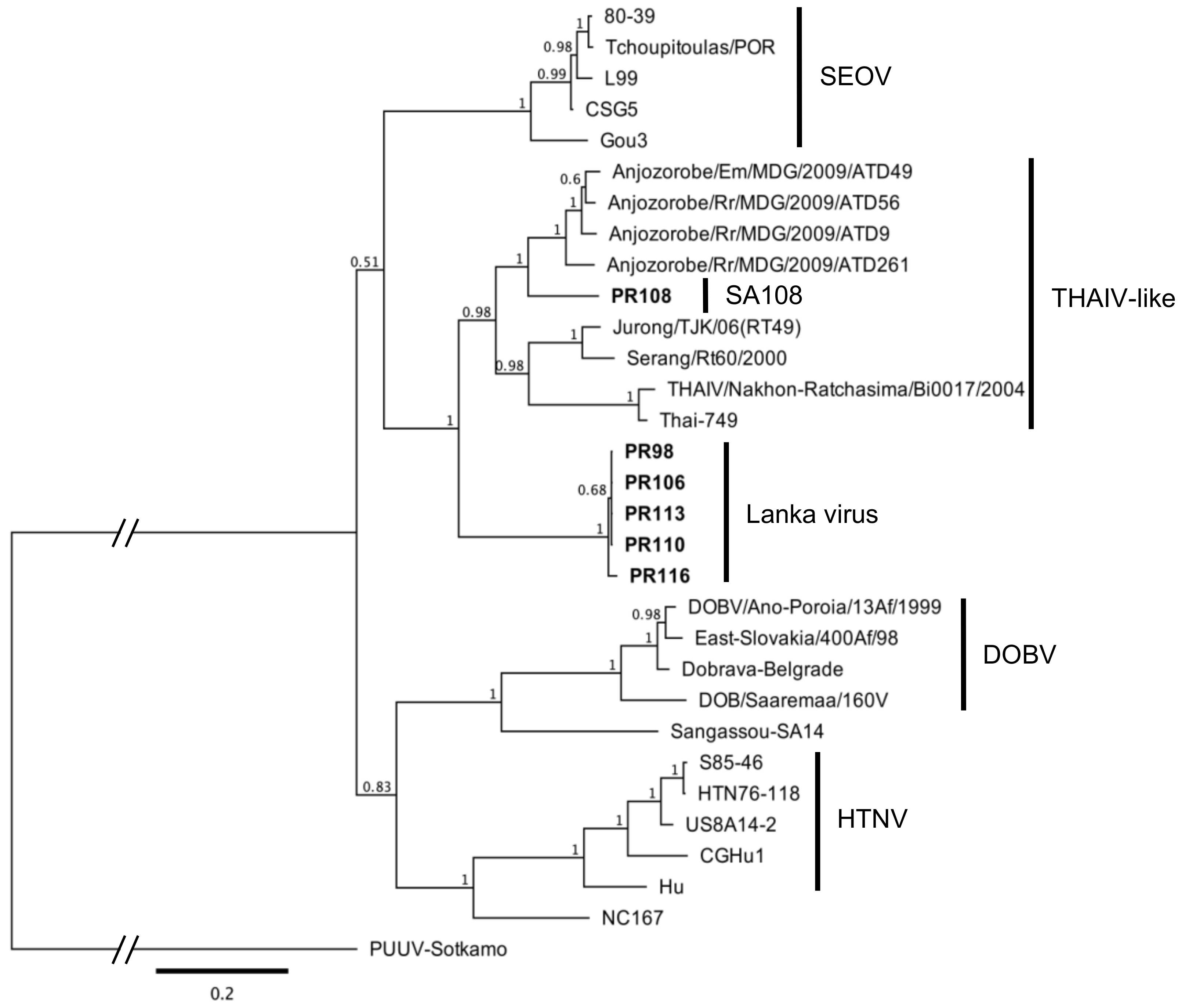
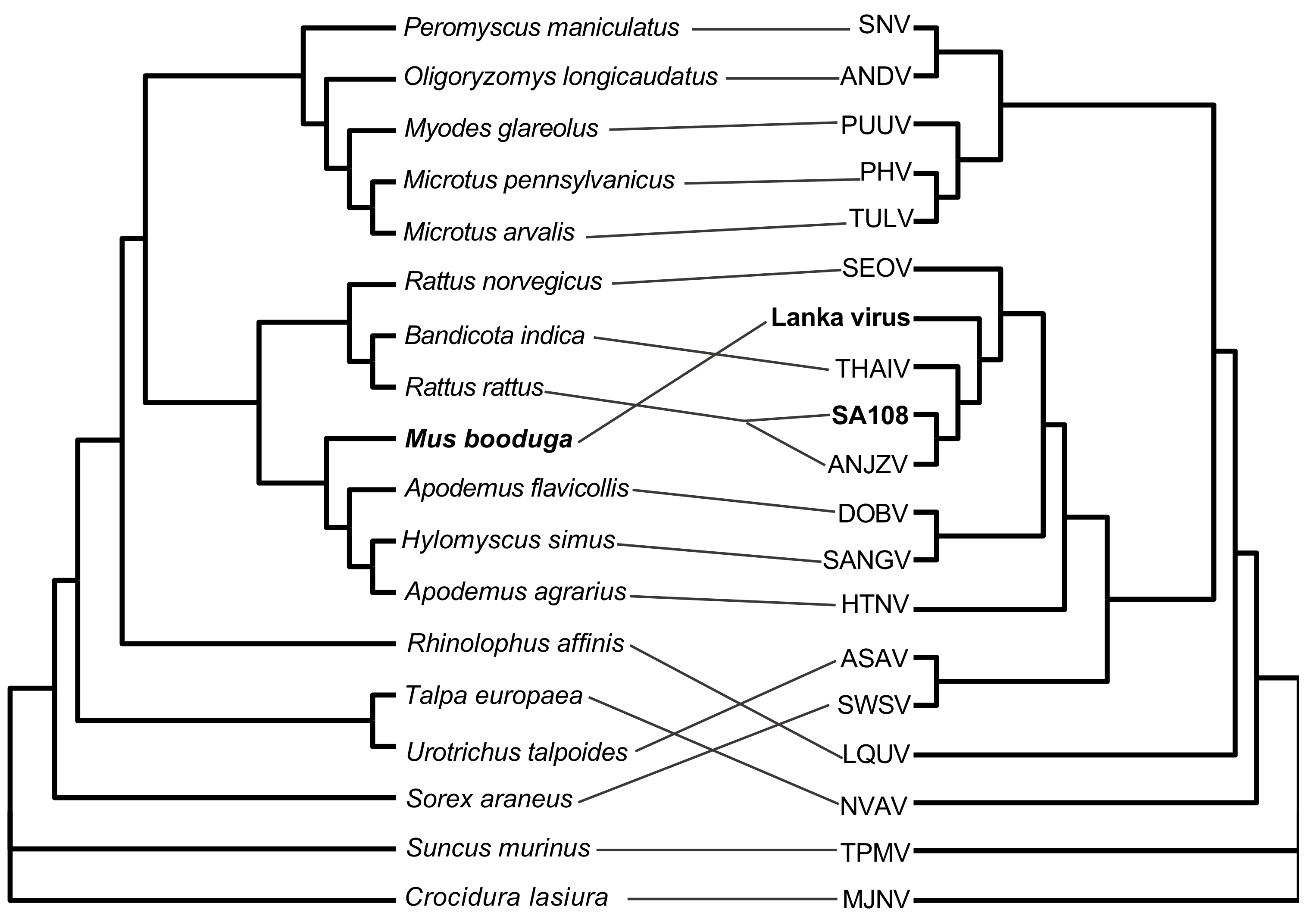
3.4. Sequence Alignment and Phylogenetic Analysis
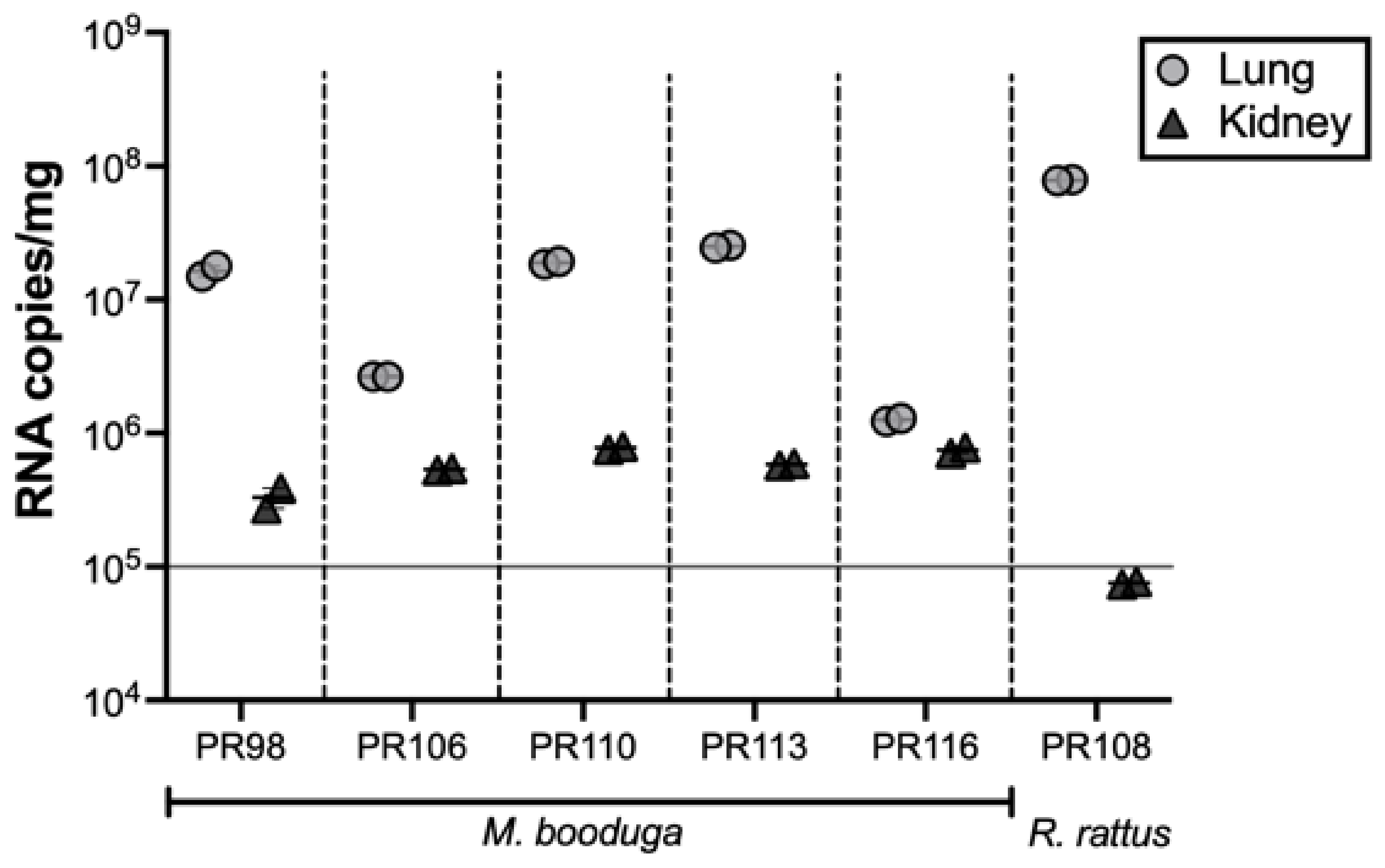
3.5. Quantification of Viral RNA
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chandrajith, R.; Nanayakkara, S.; Itai, K.; Aturaliya, T.N.; Dissanayake, C.B.; Abeysekera, T.; Harada, K.; Watanabe, T.; Koizumi, A. Chronic kidney diseases of uncertain etiology (CKDue) in Sri Lanka: Geographic distribution and environmental implications. Environ. Geochem. Health 2011, 33, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Athuraliya, N.T.; Abeysekera, T.D.; Amerasinghe, P.H.; Kumarasiri, R.; Bandara, P.; Karunaratne, U.; Milton, A.H.; Jones, A.L. Uncertain etiologies of proteinuric-chronic kidney disease in rural Sri Lanka. Kidney Int. 2011, 80, 1212–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayasekara, K.B.; Dissanayake, D.M.; Sivakanesan, R.; Ranasinghe, A.; Karunarathna, R.H.; Priyantha Kumara, G.W. Epidemiology of chronic kidney disease, with special emphasis on chronic kidney disease of uncertain etiology, in the north central region of Sri Lanka. J. Epidemiol. 2015, 25, 275–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alwis, A.A.P.; Panawala, P.V.S. A review of the national response to CKDu in Sri Lanka. Sri Lanka J. Soc. Sci. 2019, 42, 83–100. [Google Scholar] [CrossRef]
- Elliot, R.M. Molecular biology of the Bunyaviridae. J. Gen. Virol. 1990, 71, 501–522. [Google Scholar] [CrossRef] [PubMed]
- Avsic-Zupanc, T.; Saksida, A.; Korva, M. Hantavirus infections. Clin. Microbiol. Infect. 2019, 21, e6–e16. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Ma, H.; Shu, J.; Zhang, Q.; Han, M.; Liu, Z.; Jin, X.; Zhang, F.; Wu, X. Vaccines and Therapeutics Against Hantaviruses. Front. Microbiol. 2019, 10, 2989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kariwa, H.; Yoshimatsu, K.; Arikawa, J. Hantavirus infection in East Asia. Comp. Immunol. Microbiol. Infect. Dis. 2007, 30, 341–356. [Google Scholar] [CrossRef] [PubMed]
- Elwell, M.R.; Ward, G.S.; Tingpalapong, M.; LeDuc, J.W. Serologic evidence of Hantaan-like virus in rodents and man in Thailand. Southeast. Asian J. Trop. Med. Public Health 1985, 16, 349–354. [Google Scholar] [PubMed]
- Reynes, J.M.; Razafindralambo, N.K.; Lacoste, V.; Olive, M.M.; Barivelo, T.A.; Soarimalala, V.; Heraud, J.M.; Lavergne, A. Anjozorobe hantavirus, a new genetic variant of Thailand virus detected in rodents from Madagascar. Vector Borne Zoonotic. Dis. 2014, 14, 212–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plyusnina, A.; Ibrahim, I.N.; Plyusnin, A. A newly recognized hantavirus in the Asian house rat (Rattus tanezumi) in Indonesia. J. Gen. Virol. 2009, 90 Pt 1, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Johansson, P.; Yap, G.; Low, H.T.; Siew, C.C.; Kek, R.; Ng, L.C.; Bucht, G. Molecular characterization of two hantavirus strains from different rattus species in Singapore. Virol. J. 2010, 7, 15. [Google Scholar] [CrossRef] [Green Version]
- Filippone, C.; Castel, G.; Murri, S.; Beaulieux, F.; Ermonval, M.; Jallet, C.; Wise, E.L.; Ellis, R.J.; Marston, D.A.; McElhinney, L.M.; et al. Discovery of hantavirus circulating among Rattus rattus in French Mayotte island, Indian Ocean. J. Gen. Virol. 2016, 97, 1060–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pattamadilok, S.; Lee, B.H.; Kumperasart, S.; Yoshimatsu, K.; Okumura, M.; Nakamura, I.; Araki, K.; Khoprasert, Y.; Dangsupa, P.; Panlar, P.; et al. Geographical distribution of hantaviruses in Thailand and potential human health significance of Thailand virus. Am. J. Trop. Med. Hyg. 2006, 75, 994–1002. [Google Scholar] [CrossRef]
- Chandy, S.; Okumura, M.; Yoshimatsu, K.; Ulrich, R.G.; John, G.T.; Abraham, P.; Arikawa, J.; Sridharan, G. Hantavirus species in India: A retrospective study. Indian J. Med. Microbiol. 2009, 27, 348–350. [Google Scholar] [CrossRef]
- Gamage, C.D.; Yasuda, S.P.; Nishio, S.; Kularatne, S.A.; Weerakoon, K.; Rajapakse, J.; Nwafor-Okoli, C.; Lee, R.B.; Obayashi, Y.; Yoshimatsu, K.; et al. Serological evidence of Thailand virus-related hantavirus infection among suspected leptospirosis patients in Kandy, Sri Lanka. Jpn. J. Infect. Dis. 2011, 64, 72–75. [Google Scholar] [PubMed]
- Rabemananjara, H.A.; Raharinosy, V.; Razafimahefa, R.M.; Ravalohery, J.P.; Rafisandratantsoa, J.T.; Andriamandimby, S.F.; Rajerison, M.; Rahelinirina, S.; Harimanana, A.; Irinantenaina, J.; et al. Human Exposure to Hantaviruses Associated with Rodents of the Murinae Subfamily, Madagascar. Emerg. Infect. Dis. 2020, 26, 587–590. [Google Scholar] [CrossRef] [Green Version]
- Vitarana, T.; Colombage, G.; Bandaranayake, V.; Lee, H.W. Hantavirus disease in Sri Lanka [letter]. Lancet 1988, 2, 1263. [Google Scholar]
- Dahanayaka, N.J.; Agampodi, S.B.; Bandaranayaka, A.K.; Priyankara, S.; Vinetz, J.M. Hantavirus infection mimicking leptospirosis: How long are we going to rely on clinical suspicion? J. Infect. Dev. Ctries 2014, 8, 1072–1075. [Google Scholar] [CrossRef] [Green Version]
- Sunil-Chandra, N.P.; Clement, J.; Maes, P.; de Silva, H.J.; van Esbroeck, M.; van Ranst, M. Concomitant leptospirosis-hantavirus co-infection in acute patients hospitalized in Sri Lanka: Implications for a potentially worldwide underestimated problem. Epidemiol. Infect. 2015, 143, 2081–2093. [Google Scholar] [CrossRef] [Green Version]
- Gamage, C.D.; Yoshimatsu, K.; Sarathkumara, Y.D.; Thiviyaaluxmi, K.; Nanayakkara, N.; Arikawa, J. Serological evidence of hantavirus infection in Girandurukotte, an area endemic for chronic kidney disease of unknown aetiology (CKDu) in Sri Lanka. Int. J. Infect. Dis. 2017, 57, 77–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimatsu, K.; Gamage, C.D.; Sarathkumara, Y.D.; Kulendiran, T.; Muthusinghe, D.S.; Nanayakkara, N.; Gunarathne, L.; Shimizu, K.; Tsuda, Y.; Arikawa, J. Thailand orthohantavirus infection in patients with chronic kidney disease of unknown aetiology in Sri Lanka. Arch. Virol. 2019, 164, 267–271. [Google Scholar] [CrossRef]
- Wijkstrom, J.; Jayasumana, C.; Dassanayake, R.; Priyawardane, N.; Godakanda, N.; Siribaddana, S.; Ring, A.; Hultenby, K.; Soderberg, M.; Elinder, C.G.; et al. Morphological and clinical findings in Sri Lankan patients with chronic kidney disease of unknown cause (CKDu): Similarities and differences with Mesoamerican Nephropathy. PLoS ONE 2018, 13, e0193056. [Google Scholar] [CrossRef] [PubMed]
- Sarathkumara, Y.D.; Gamage, C.D.; Lokupathirage, S.; Muthusinghe, D.S.; Nanayakkara, N.; Gunarathne, L.; Shimizu, K.; Tsuda, Y.; Arikawa, J.; Yoshimatsu, K. Exposure to Hantavirus is a Risk Factor Associated with Kidney Diseases in Sri Lanka: A Cross Sectional Study. Viruses 2019, 11, 700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lokupathirage, S.M.W.; Muthusinghe, D.S.; Shimizu, K.; Nishigami, K.; Noda, K.; Tsuda, Y.; Sarathkumara, Y.D.; Gunawardana, S.; Arikawa, J.; Gamage, C.D.; et al. Serological Evidence of Thailand Orthohantavirus or Antigenically Related Virus Infection Among Rodents in a Chronic Kidney Disease of Unknown Etiology Endemic Area, Girandurukotte, Sri Lanka. Vector Borne Zoonotic Dis. 2019, 19, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Gamage, C.D.; Sarathkumara, Y.D. Chronic kidney disease of uncertain etiology in Sri Lanka: Are leptospirosis and Hantaviral infection likely causes? Med. Hypotheses 2016, 91, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, S.P.; Vogel, P.; Tsuchiya, K.; Han, S.H.; Lin, L.K.; Suzuki, H. Phylogeographic patterning of mtDNA in the widely distributed harvest mouse (Micromys minutus) suggests dramatic cycles of range contraction and expansion during the mid- to late Pleistocene. Can. J. Zool 2005, 83, 1411–1420. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, S.P.; Gamage, C.D.; Koizumi, N.; Nishio, S.; Isozumi, R.; Shimizu, K.; Koma, T.; Amada, T.; Suzuki, H.; Yoshimatsu, K.; et al. Distinct genetic characteristics of Sri Lankan Rattus and Bandicota (Murinae, Rodentia) inferred from mitochondrial and nuclear markers. Genes Genet. Syst. 2014, 89, 71–80. [Google Scholar] [CrossRef] [Green Version]
- Yoshimatsu, K.; Arikawa, J.; Kariwa, H. Application of a recombinant baculovirus expressing hantavirus nucleocapsid protein as a diagnostic antigen in IFA test: Cross reactivities among 3 serotypes of hantavirus which causes hemorrhagic fever with renal syndrome (HFRS). J. Vet. Med. Sci. 1993, 55, 1047–1050. [Google Scholar] [CrossRef] [Green Version]
- Klempa, B.; Fichet-Calvet, E.; Lecompte, E.; Auste, B.; Aniskin, V.; Meisel, H.; Denys, C.; Koivogui, L.; ter Meulen, J.; Kruger, D.H. Hantavirus in African wood mouse, Guinea. Emerg. Infect. Dis. 2006, 12, 838–840. [Google Scholar] [CrossRef]
- Ito, N.; Kakemizu, M.; Ito, K.A.; Yamamoto, A.; Yoshida, Y.; Sugiyama, M.; Minamoto, N. A comparison of complete genome sequences of the attenuated RC-HL strain of rabies virus used for production of animal vaccine in Japan, and the parental Nishigahara strain. Microbiol. Immunol. 2001, 45, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Huson, D.H.; Scornavacca, C. Dendroscope 3: An interactive tool for rooted phylogenetic trees and networks. Syst. Biol. 2012, 61, 1061–1067. [Google Scholar] [CrossRef] [Green Version]
- Laenen, L.; Vergote, V.; Calisher, C.H.; Klempa, B.; Klingstrom, J.; Kuhn, J.H.; Maes, P. Hantaviridae: Current Classification and Future Perspectives. Viruses 2019, 11, 788. [Google Scholar] [CrossRef] [Green Version]
- Baek, L.J.; Yanagihara, R.; Gibbs, C.J.; Miyazaki, M.; Gajdusek, D.C. Leakey virus: A new hantavirus isolated from Mus musculus in the United States. J. Gen. Virol. 1988, 69 Pt 12, 3129–3132. [Google Scholar] [CrossRef]
- Arthur, R.R.; Lofts, R.S.; Gomez, J.; Glass, G.E.; Leduc, J.W.; Childs, J.E. Grouping of hantaviruses by small (S) genome segment polymerase chain reaction and amplification of viral RNA from wild-caught rats. Am. J. Trop. Med. Hyg. 1992, 47, 210–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Childs, J.E.; Ksiazek, T.G.; Spiropoulou, C.F.; Krebs, J.W.; Morzunov, S.; Maupin, G.O.; Gage, K.L.; Rollin, P.E.; Sarisky, J.; Enscore, R.E.; et al. Serologic and genetic identification of Peromyscus maniculatus as the primary rodent reservoir for a new hantavirus in the southwestern United States. J. Infect. Dis. 1994, 169, 1271–1280. [Google Scholar] [CrossRef] [Green Version]
- Weidmann, M.; Schmidt, P.; Vackova, M.; Krivanec, K.; Munclinger, P.; Hufert, F.T. Identification of genetic evidence for dobrava virus spillover in rodents by nested reverse transcription (RT)-PCR and TaqMan RT-PCR. J. Clin. Microbiol. 2005, 43, 808–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.F.; Zuo, S.Q.; Zhang, W.Y.; Wu, X.M.; Tang, F.; De Vlas, S.J.; Zhao, W.J.; Zhang, P.H.; Dun, Z.; Wang, R.M.; et al. Prevalence and genetic diversities of hantaviruses in rodents in Beijing, China. Am. J. Trop. Med. Hyg. 2008, 78, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Aplin, K.; Molur, S.; Nameer, P.O. Mus booduga (errata version published in 2017). IUCN Red List Threat. Species 2016. [Google Scholar] [CrossRef]
- Plyusnin, A.; Vapalahti, O.; Lundkvist, A. Hantaviruses: Genome structure, expression and evolution. J. Gen. Virol. 1996, 77, 2677–2687. [Google Scholar] [CrossRef] [PubMed]
- Schmaljohn, C.; Hjelle, B. Hantaviruses—A Global Disease Problem. Emerg. Infect. Dis. 1997, 3, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, S.P.; Shimizu, K.; Koma, T.; Hoa, N.T.; Le, M.Q.; Wei, Z.; Muthusinghe, D.S.; Lokupathirage, S.M.W.; Hasebe, F.; Yamashiro, T.; et al. Immunological Responses to Seoul Orthohantavirus in Experimentally and Naturally Infected Brown Rats (Rattus norvegicus). Viruses 2021, 13, 665. [Google Scholar] [CrossRef] [PubMed]
- Correa-Rotter, R.; Wesseling, C.; Johnson, R.J. CKD of unknown origin in Central America: The case for a Mesoamerican nephropathy. Am. J. Kidney Dis. 2014, 63, 506–520. [Google Scholar] [CrossRef]
- Sunil-Chandra, N.P.; Jayaweera, J.; Kumbukgolla, W.; Jayasundara, M. Association of Hantavirus Infections and Leptospirosis With the Occurrence of Chronic Kidney Disease of Uncertain Etiology in the North Central Province of Sri Lanka: A Prospective Study With Patients and Healthy Persons. Front. Cell Infect. Microbiol. 2020, 10, 556737. [Google Scholar] [CrossRef] [PubMed]
- Yih, W.K.; Kulldorff, M.; Friedman, D.J.; Leibler, J.H.; Amador, J.J.; Lopez-Pilarte, D.; Galloway, R.L.; Ramirez-Rubio, O.; Riefkohl, A.; Brooks, D.R. Investigating Possible Infectious Causes of Chronic Kidney Disease of Unknown Etiology in a Nicaraguan Mining Community. Am. J. Trop. Med. Hyg. 2019, 101, 676–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | No. of Captured Animals | IFA Antibody (% Positive) | PCR (% Positive) |
|---|---|---|---|
| Rattus rattus complex | 99 | 34 (34.3%) | 2 (2%) |
| Mus booduga | 11 | 5 (45.5%) | 5 (45.5%) |
| Tatera indica | 3 | 0 | 0 |
| Bandicota bengalensis | 1 | 1 | 0 |
| Bandicota indica | 1 | 0 | 0 |
| Crocidura horsfieldii | 1 | 0 | 0 |
| Total | 116 | 40 | 7 |
| Lanka Virus Strain-PR 98 | ||||||
|---|---|---|---|---|---|---|
| Nucleotide Identity, % | Amino Acid Identity, % | |||||
| ORF_S | ORF_M | ORF_L | N | GPC | L-protein | |
| THAIV | 78.1 | 76.6 | 79.2 | 92.1 | 85.4 | 94.0 |
| ANJZV | 79.4 | 76.9 | 79.5 | 93.2 | 86.7 | 94.3 |
| SA108 | 80.1 | 76.9 | 79.7 | 93.0 | 87.2 | 94.5 |
| SEOV | 75.0 | 72.3 | 76.7 | 85.8 | 79.7 | 88.3 |
| HTNV | 74.0 | 71.2 | 74.0 | 84.1 | 76.8 | 84.8 |
| DOBV | 72.6 | 70.7 | 74.2 | 83.2 | 75.7 | 85.6 |
| PUUV | 62.6 | 59.4 | 66.9 | 61.4 | 53.6 | 68.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muthusinghe, D.S.; Shimizu, K.; Lokupathirage, S.M.W.; Wei, Z.; Sarathkumara, Y.D.; Fonseka, G.R.A.; Senarathne, P.; Koizumi, N.; Kawakami, T.; Koizumi, A.; et al. Identification of Novel Rodent-Borne Orthohantaviruses in an Endemic Area of Chronic Kidney Disease of Unknown Etiology (CKDu) in Sri Lanka. Viruses 2021, 13, 1984. https://0-doi-org.brum.beds.ac.uk/10.3390/v13101984
Muthusinghe DS, Shimizu K, Lokupathirage SMW, Wei Z, Sarathkumara YD, Fonseka GRA, Senarathne P, Koizumi N, Kawakami T, Koizumi A, et al. Identification of Novel Rodent-Borne Orthohantaviruses in an Endemic Area of Chronic Kidney Disease of Unknown Etiology (CKDu) in Sri Lanka. Viruses. 2021; 13(10):1984. https://0-doi-org.brum.beds.ac.uk/10.3390/v13101984
Chicago/Turabian StyleMuthusinghe, Devinda S., Kenta Shimizu, Sithumini M. W. Lokupathirage, Zhouoxing Wei, Yomani D. Sarathkumara, G. R. Amanda Fonseka, Pavani Senarathne, Nobuo Koizumi, Tomonori Kawakami, Akio Koizumi, and et al. 2021. "Identification of Novel Rodent-Borne Orthohantaviruses in an Endemic Area of Chronic Kidney Disease of Unknown Etiology (CKDu) in Sri Lanka" Viruses 13, no. 10: 1984. https://0-doi-org.brum.beds.ac.uk/10.3390/v13101984