
Expression of an Antiviral Gene GmRUN1 from Soybean Is Regulated via Intron-Mediated Enhancement (IME)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Growth Conditions
2.2. Construction of Recombinant Vectors
2.3. Transient Expression and Virus Inoculation
2.4. Promoter Analysis and In Situ GUS Activity Assay
2.5. Luciferase Reporter Assay and Fluorescence Quantitative Analysis
2.6. Quantitative Real-Time PCR (qRT-PCR) Analysis
3. Results
3.1. GmRUN1 cDNA Shows Partial Resistance to TMV and SMV in N. Benthamiana Transient Expression Assay
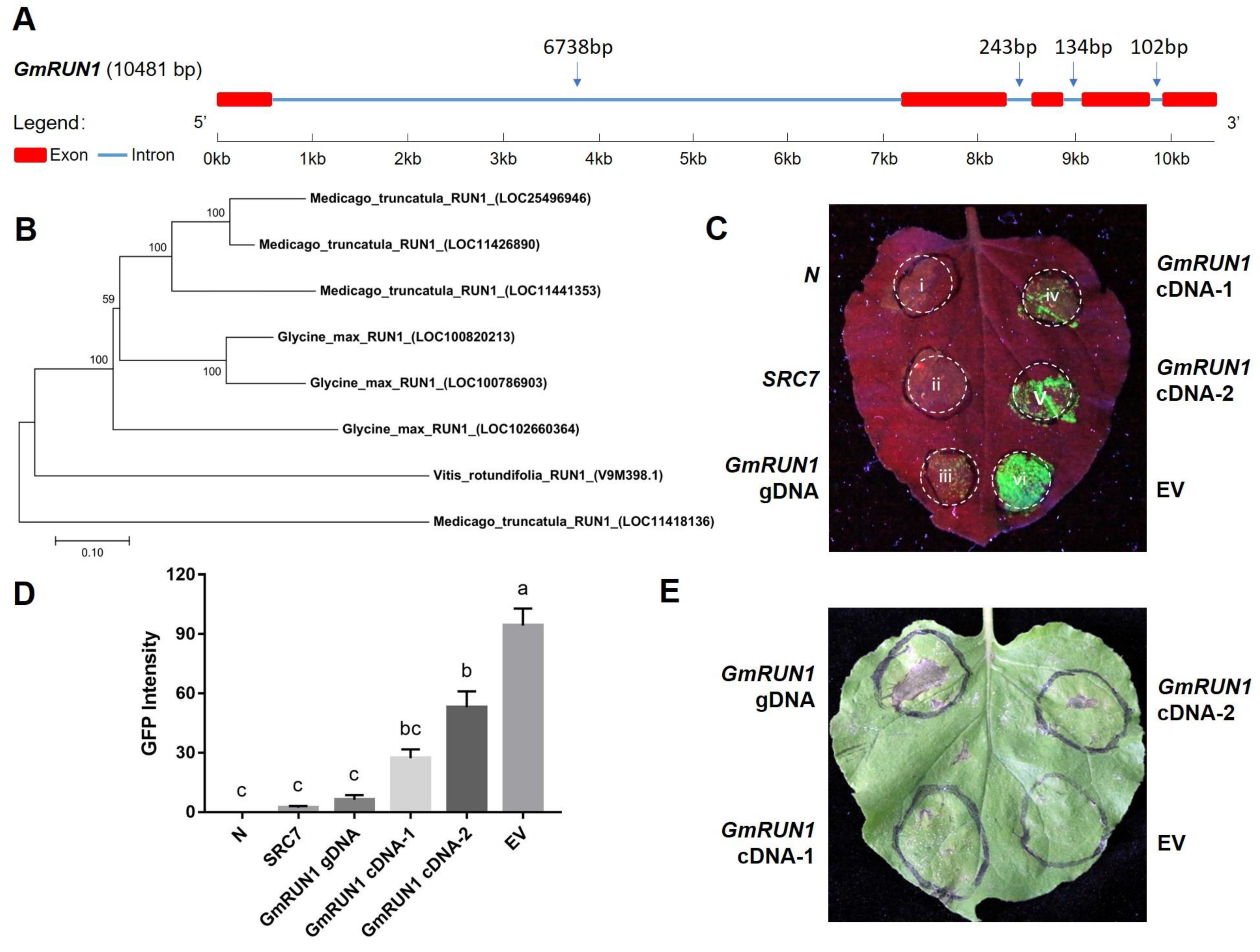
3.2. Three Canonical Domains of GmRUN1 Are Indispensable for Its Antiviral Activity
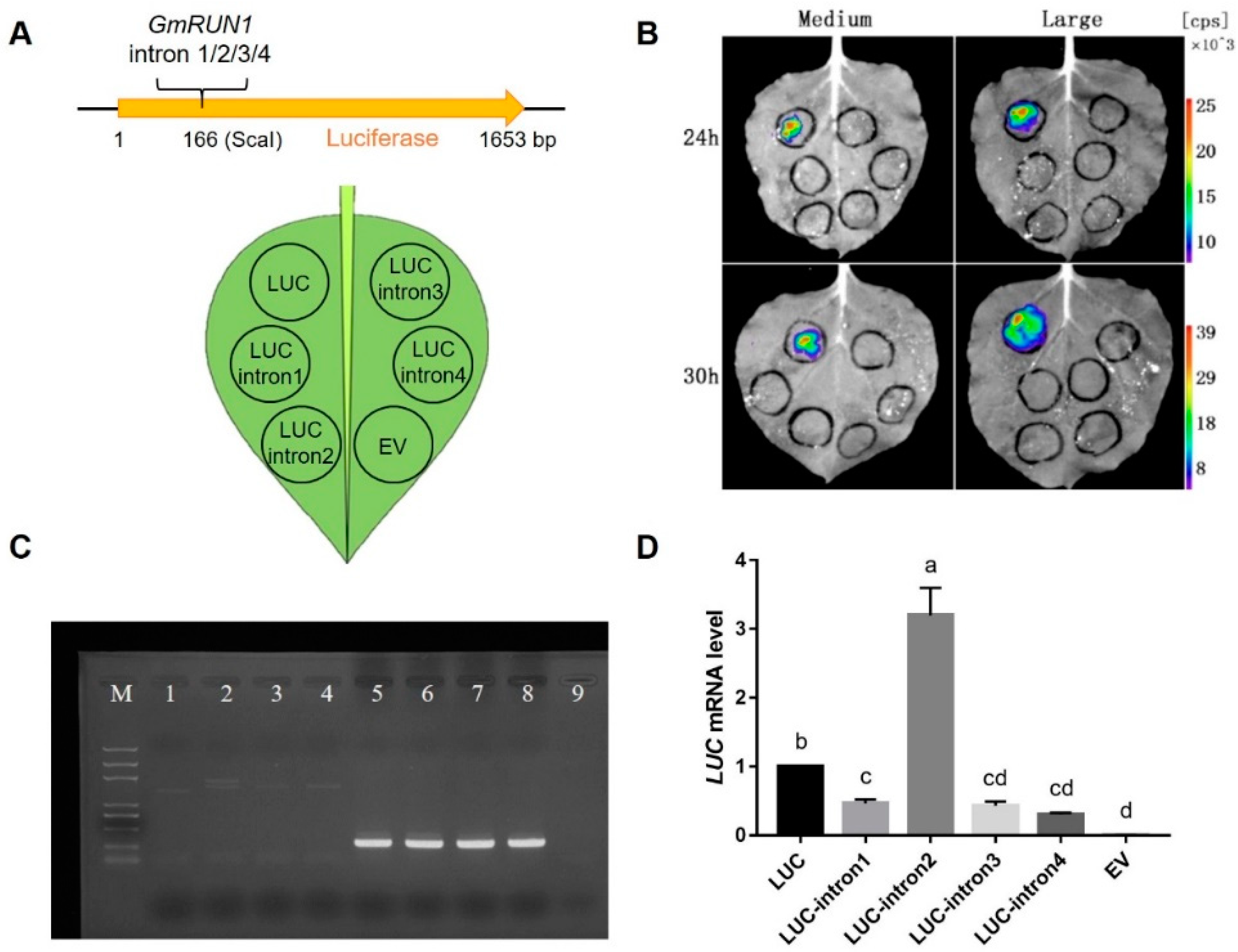
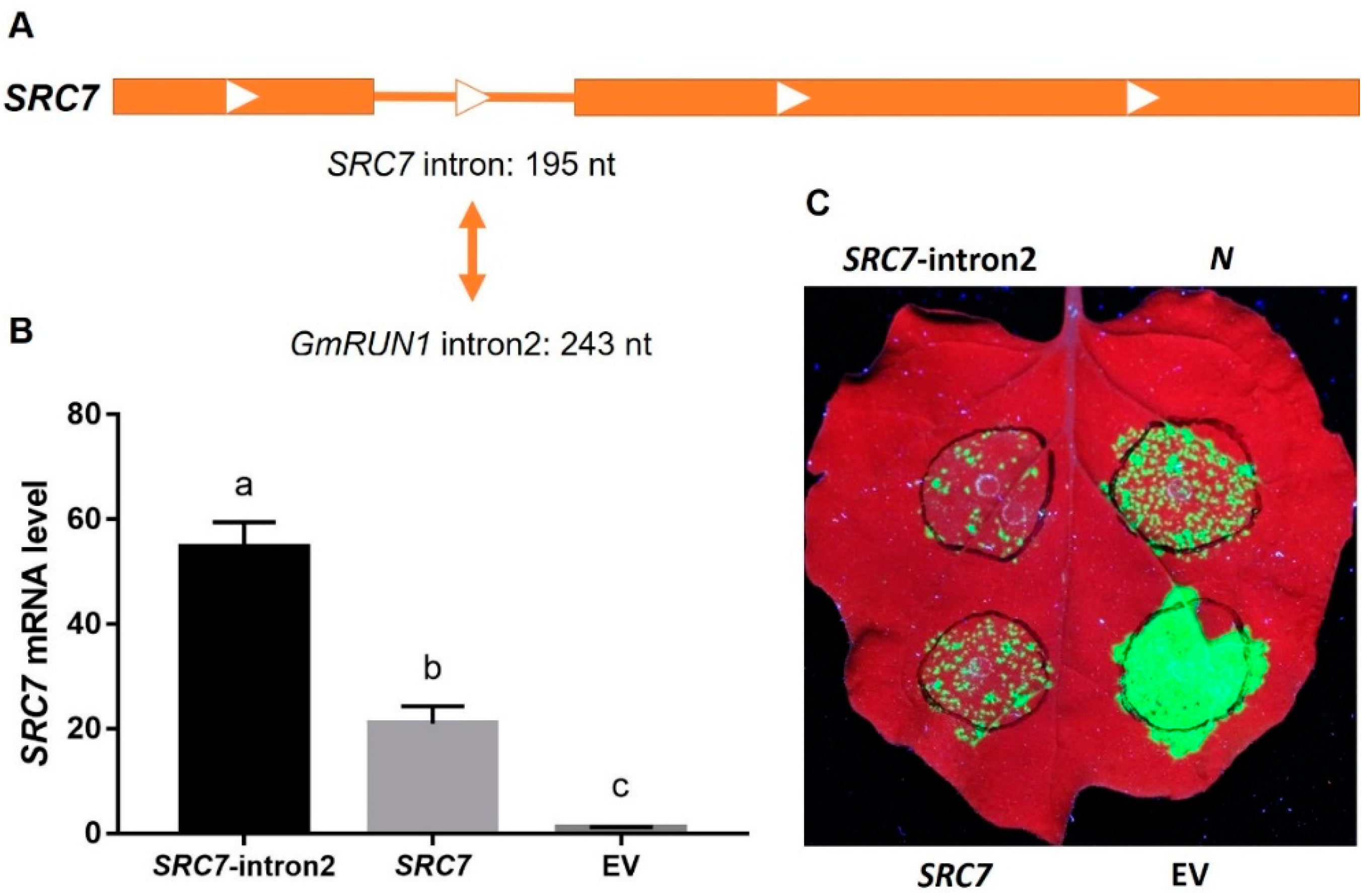
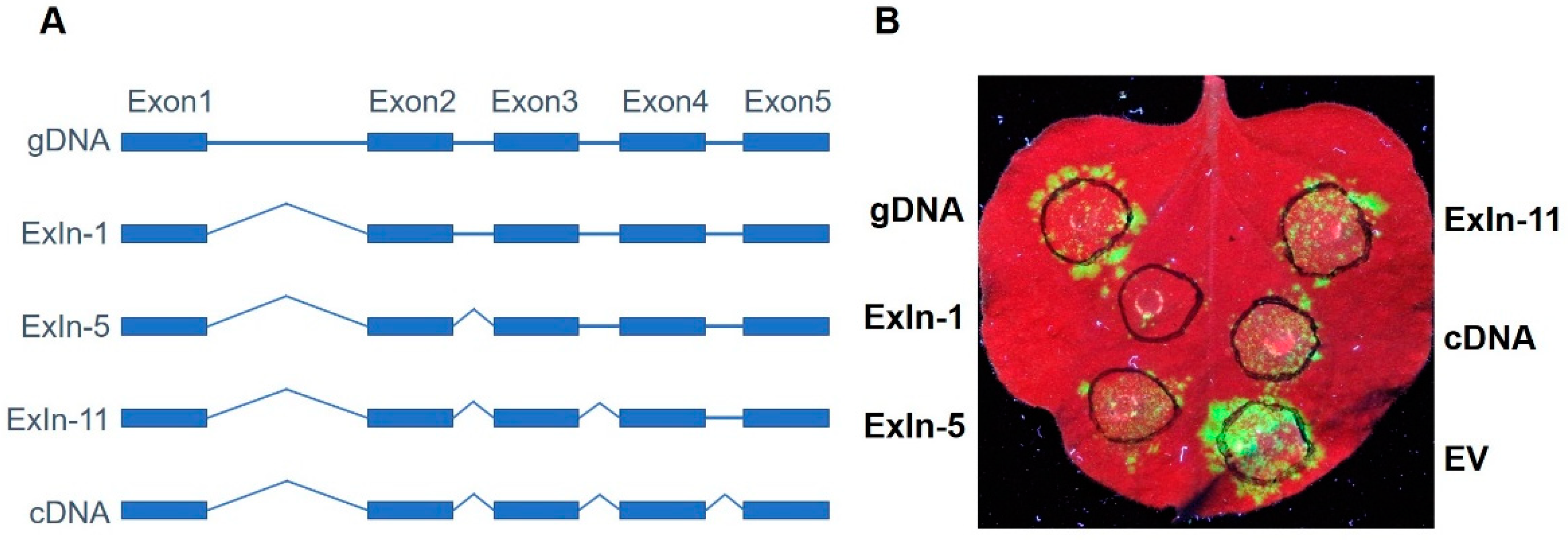
3.3. GmRUN1 Has IME Phenomena
3.4. GmRUN1 Expression Is Transcriptionally Regulated by SA
3.5. GmRUN1 Expression Is Transcriptionally Induced by Transcription Factor GmDREB3
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zipfel, C. Pattern-recognition receptors in plant innate immunity. Curr. Opin. Immunol. 2008, 20, 10–16. [Google Scholar] [CrossRef]
- Boller, T.; He, S.Y. Innate immunity in plants: An arms race between pattern recognition receptors in plants and effectors in microbial pathogens. Science 2009, 324, 742–744. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Staskawicz, B.J.; Ausubel, F.M.; Baker, B.J.; Ellis, J.G.; Jones, J. Molecular genetics of plant disease resistance. Science 1995, 268, 661–667. [Google Scholar] [CrossRef] [Green Version]
- Kourelis, J.; van der Hoorn, R.A.L. Defended to the Nines: 25 years of resistance gene cloning identifies nine mechanisms for r protein function. Plant Cell 2018, 30, 285–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, S.; Zhang, Y.; Hu, Q.; Chen, J.; Li, K.; Lu, C.; Liu, H.; Wang, W.; Kuang, H. Dynamic nucleotide-binding site and leu-cine-rich repeat-encoding genes in the grass family. Plant Physiol. 2012, 159, 197–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.; Du, C.; Covaleda, L.; Xu, Z.; Robinson, A.F.; Yu, J.Z.; Kohel, R.J.; Zhang, H.-B. Cloning, characterization, and evolution of the nbs-lrr-encoding resistance gene analogue family in polyploid cotton (Gossypium hirsutum L.). Mol. Plant-Microbe. Interact. 2004, 17, 1234–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hulbert, S.H.; Webb, C.A.; Smith, S.M.; Sun, Q. Resistance gene complexes: Evolution and utilization. Annu. Rev. Phytopathol. 2001, 39, 285–312. [Google Scholar] [CrossRef] [PubMed]
- Lelpe, D.D.; Koonin, E.V.; Aravind, L. STAND, a class of P-loop NTPases including animal and plant regulators of pro-grammed cell death: Multiple, complex domain architectures, unusual phyletic patterns, and evolution by horizontal gene transfer. J. Mol. Biol. 2004, 343, 1–28. [Google Scholar]
- Walker, J.E.; Saraste, M.; Runswick, M.J.; Gay, N.J. Distantly related sequences in the alpha subunits and beta subunits of atp synthase myosin kinases and other atp requiring enzymes and a common nucleotide binding fold. EMBO J. 1982, 1, 945–952. [Google Scholar] [CrossRef]
- Tameling, W.I.L.; Elzinga, S.D.J.; Darmin, P.S.; Vossen, J.H.; Takken, F.; Haring, M.A.; Cornelissen, B.J.C. The tomato R gene products I-2 and Mi-1 are functional atp binding proteins with atpase activity. Plant Cell 2002, 14, 2929–2939. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, V. Role of extracytoplasmic leucine rich repeat proteins in plant defence mechanisms. Microbiol. Res. 2005, 160, 83–94. [Google Scholar] [CrossRef]
- Velásquez, A.C.; Nomura, K.; Cooper, M.D.; Herrin, B.R.; He, S.Y. Leucine-rich-repeat-containing variable lymphocyte receptors as modules to target plant-expressed proteins. Plant Methods 2017, 13, 29. [Google Scholar] [CrossRef] [Green Version]
- Ellis, J.; Dodds, P.; Pryor, T. Structure, function and evolution of plant disease resistance genes. Curr. Opin. Plant Biol. 2000, 3, 278–284. [Google Scholar] [CrossRef]
- Eitas, T.K.; Dangl, J.L. NB-LRR proteins: Pairs, pieces, perception, partners, and pathways. Curr. Opin. Plant Biol. 2010, 13, 472–477. [Google Scholar] [CrossRef] [Green Version]
- Meyers, B.C.; Dickerman, A.W.; Michelmore, R.W.; Sivaramakrishnan, S.; Sobral, B.W.; Young, N.D. Plant disease resistance genes encode members of an ancient and diverse protein family within the nucleotide-binding superfamily. Plant J. 1999, 20, 317–332. [Google Scholar] [CrossRef] [PubMed]
- Meyers, B.; Kozik, A.; Griego, A.; Kuang, H.; Michelmore, R.W. Genome-wide analysis of NBS-LRR–Encoding genes in arabidopsis. Plant Cell 2003, 15, 809–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, J.-X.; Meyers, B.C.; Chen, J.-Q.; Tian, D.; Yang, S. Tracing the origin and evolutionary history of plant nucleotide-binding site-leucine-rich repeat (NBS-LRR) genes. New Phytol. 2012, 193, 1049–1063. [Google Scholar] [CrossRef]
- Mukhtar, M.S.; Carvunis, A.-R.; Dreze, M.; Epple, P.; Steinbrenner, J.; Moore, J.; Tasan, M.; Galli, M.; Hao, T.; Nishimura, M.T.; et al. Independently Evolved Virulence Effectors Converge onto Hubs in a Plant Immune System Network. Science 2011, 333, 596–601. [Google Scholar] [CrossRef] [Green Version]
- Bieri, S.; Mauch, S.; Shen, Q.H.; Peart, J.; Devoto, A.; Casais, C.; Ceron, F.; Schulze, S.; Steinbiss, H.H.; Shirasu, K.; et al. RAR1 positively controls steady state levels of barley MLA resistance proteins and enables sufficient MLA6 ac-cumulation for effective resistance. Plant Cell 2004, 16, 3480–3495. [Google Scholar] [CrossRef] [Green Version]
- Mackey, D.; Belkhadir, Y.; Alonso, J.; Ecker, J.; Dangl, J.L. Arabidopsis RIN4 is a target of the type III virulence effector AvrRpt2 and modulates RPS2-mediated resistance. Cell 2003, 112, 379–389. [Google Scholar] [CrossRef] [Green Version]
- Lai, Y.; Eulgem, T. Transcript-level expression control of plant NLR genes. Mol. Plant Pathol. 2018, 19, 1267–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayo, M.A.; Pringle, C.R. Virus taxonomy-1997. J. Gen. Virol. 1998, 79, 649–657. [Google Scholar] [CrossRef]
- Yang, Y.; Lin, J.; Zheng, G.; Zhang, M.; Zhi, H. Recombinant soybean mosaic virus is prevalent in Chinese soybean fields. Arch. Virol. 2014, 159, 1793–1796. [Google Scholar] [CrossRef]
- Cho, E.K.; Goodman, R.M. Strains of soybean mosaic virus: Classification based on virulence in resistant soybean cultivars. Phytopathology 1979, 69, 467–470. [Google Scholar] [CrossRef]
- Cho, E.K.; Chung, B.J.; Lee, S.H. Studies on identification and classification of soybean virus diseases in Korea part 2 etiology of a necrotic disease of Glycine-max. Plant Dis. Rep. 1977, 61, 313–317. [Google Scholar]
- Saruta, M.; Kikuchi, A.; Okabe, A.; Sasaya, T. Molecular characterization of A2 and D strains of Soybean mosaic virus, which caused a recent virus outbreak in soybean cultivar Sachiyutaka in Chugoku and Shikoku regions of Japan. J. Gen. Plant Pathol. 2005, 71, 431–435. [Google Scholar] [CrossRef]
- Li, K.; Yang, Q.H.; Zhi, H.J.; Gai, J.Y. Identification and distribution of soybean mosaic virus strains in Southern China. Plant Dis. 2010, 94, 351–357. [Google Scholar] [CrossRef] [Green Version]
- Eggenberger, A.L.; Stark, D.M.; Beachy, R.N. The nucleotide sequence of a soybean mosaic virus coat protein-coding region and its expression in escherichia coli, agrobacterium tumefaciens and tobacco callus. J. Gen. Virol. 1989, 70, 1853–1860. [Google Scholar] [CrossRef]
- Liu, J.-Z.; Fang, Y.; Pang, H. The current status of the Soybean-Soybean Mosaic Virus (SMV) Pathosystem. Front. Microbiol. 2016, 7, 1906. [Google Scholar] [CrossRef]
- Zhang, C.; Grosic, S.; Whitham, S.A.; Hill, J.H. The requirement of multiple defense genes in soybean Rsv1-mediated extreme resistance to soybean mosaic virus. Mol. Plant-Microbe Interact. 2012, 25, 1307–1313. [Google Scholar] [CrossRef]
- Hajimorad, M.R.; Hill, J.H. Rsv1-mediated resistance against soybean mosaic virus-N is hypersensitive re-sponse-independent at inoculation site, but has the potential to initiate a hypersensitive response-like mechanism. Mol. Plant-Microbe Interact. 2001, 14, 587–598. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Buss, G.R.; Tolin, S.A. Resistance to soybean mosaic virus conferred by two independent dominant genes in PI 486355. J. Hered. 1993, 84, 25–28. [Google Scholar] [CrossRef]
- Ishibashi, K.; Saruta, M.; Shimizu, T.; Shu, M.; Anai, T.; Komatsu, K.; Yamada, N.; Katayose, Y.; Ishikawa, M.; Ishimoto, M.; et al. Soybean antiviral immunity conferred by dsRNase targets the viral replication complex. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilut, D.C.; Lipka, A.E.; Jeong, N.; Bae, D.N.; Kim, D.H.; Kim, J.H.; Redekar, N.; Yang, K.; Park, W.; Kang, S.-T.; et al. Identification of haplotypes at the Rsv4 genomic region in soybean associated with durable resistance to soybean mosaic virus. Theor. Appl. Genet. 2016, 129, 453–468. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Li, H.; Sun, H.; Li, A.; Liu, S.; Yu, R.; Cui, X.; Zhang, D.; Wuriyanghan, H.; Su, H. Salicylic acid and broad spectrum of NBS-LRR family genes are involved in SMV-soybean interactions. Plant Physiol. Biochem. 2018, 123, 132–140. [Google Scholar] [CrossRef]
- Goodin, M.M.; Zaitlin, D.; Naidu, R.A.; Lommel, S.A. Nicotiana benthamiana: Its history and future as a model for plant–pathogen interactions. Mol. Plant-Microbe Interact. 2014, 21, 1015–1026. [Google Scholar] [CrossRef] [Green Version]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3908. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T) (-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Feechan, A.; Kocsis, M.; Riaz, S.; Zhang, W.; Gadoury, D.M.; Walker, M.A.; Dry, I.B.; Reisch, B.; Cadle-Davidson, L. Strategies for RUN1 deployment using RUN2 and REN2 to manage grapevine powdery mildew informed by studies of race specificity. Phytopathology 2015, 105, 1104–1113. [Google Scholar] [CrossRef] [Green Version]
- Pauquet, J.; Bouquet, A.; This, P.; Adam-Blondon, A.-F. Establishment of a local map of AFLP markers around the powdery mildew resistance gene Run1 in grapevine and assessment of their usefulness for marker assisted selection. Theor. Appl. Genet. 2001, 103, 1201–1210. [Google Scholar] [CrossRef]
- Lindbo, J.A. TRBO: A high-efficiency tobacco mosaic virus RNA-based overexpression vector. Plant Physiol. 2007, 145, 1232–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitham, S.; Dinesh-Kumar, S.P.; Choi, D.; Hehl, R.; Corr, C.; Baker, B. The product of the tobacco mosaic virus resistance gene N: Similarity to toll and the interleukin-1 receptor. Cell 1994, 78, 1101–1115. [Google Scholar] [CrossRef]
- Callis, J.; Fromm, M.; Walbot, V. Introns increase gene expression in cultured maize cells. Genes Dev. 1987, 1, 1183–1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, A.B. Requirements for intron-mediated enhancement of gene expression in Arabidopsis. RNA 2002, 8, 1444–1453. [Google Scholar] [CrossRef] [Green Version]
- Zhao, T.; Rui, L.; Li, J.; Nishimura, M.; Vogel, J.; Liu, N.; Liu, S.; Zhao, Y.; Dangl, J.L.; Tang, D. A Truncated NLR protein, TIR-NBS2, is required for activated defense responses in the exo70B1 Mutant. PLoS Genet. 2015, 11, e1004945. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.; Liu, J.; Ding, Y.; Wang, S.; Zhang, X.; Liu, Y.; Yang, S. Temperature-dependent autoimmunity mediated by chs1 requires its neighboring TNL gene SOC 3. New Phytol. 2017, 213, 1330–1345. [Google Scholar] [CrossRef]
- Nandety, R.S.; Caplan, J.L.; Cavanaugh, K.; Perroud, B.; Wroblewski, T.; Michelmore, R.W.; Meyers, B.C. The role of TIR-NBS and TIR-X proteins in plant basal defense responses. Plant Physiol. 2013, 162, 1459–1472. [Google Scholar] [CrossRef] [Green Version]
- Neupane, S.; Ma, Q.; Mathew, F.M.; Varenhorst, A.J.; Andersen, E.J.; Nepal, M.P. Evolutionary divergence of TNL disease-resistant proteins in soybean (Glycine max) and common bean (Phaseolus vulgaris). Biochem. Genet. 2018, 56, 397–422. [Google Scholar] [CrossRef]
- Karasov, T.L.; Chae, E.; Herman, J.J.; Bergelson, J. Mechanisms to mitigate the trade-off between growth and defense. Plant Cell 2017, 29, 666–680. [Google Scholar] [CrossRef] [Green Version]
- Garreton, V.; Carpinelli, J.; Jordana, X.; Holuigue, L. The as-1 promoter element is an oxidative stress-responsive element and salicylic acid activates it via oxidative species. Plant Physiol. 2002, 130, 1516–1526. [Google Scholar] [CrossRef] [Green Version]
- Grüner, R.; Strompen, G.; Pfitzner, A.J.P.; Pfitzner, U.M. Salicylic acid and the hypersensitive response initiate distinct signal transduction pathways in tobacco that converge on the as-1-like element of the PR-1a promoter. JBIC J. Biol. Inorg. Chem. 2003, 270, 4876–4886. [Google Scholar]
- Zhou, J.-M.; Zhang, Y. Plant Immunity: Danger Perception and Signaling. Cell 2020, 181, 978–989. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Xu, Z.P.; Lv, R.; Zhu, L.S.; Peng, Q.D.; Qiu, L.; Xi, D.H. N gene enhances resistance to Chili veinal mottle virus and hypersensitivity to salt stress in tobacco. J. Plant Physiol. 2018, 230, 92–100. [Google Scholar] [CrossRef]
- Gutterson, N.; Reuber, T.L. Regulation of disease resistance pathways by AP2/ERF transcription factors. Curr. Opin. Plant Biol. 2004, 7, 465–471. [Google Scholar] [CrossRef]
- Huang, P.-Y.; Catinot, J.; Zimmerli, L. Ethylene response factors in Arabidopsis immunity. J. Exp. Bot. 2015, 67, 1231–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Deng, Y.; Hu, J.; Jin, C.; Zhu, X.; Li, D. Genome-wide analyses of direct target genes of an ERF11 transcription factor involved in plant defense against bacterial pathogens. Biochem. Biophys. Res. Commun. 2020, 532, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Hormuzdi, S.G.; Penttinen, R.; Jaenisch, R.; Bornstein, P. A Gene-targeting approach identifies a function for the first intron in expression of the alpha 1 (I) collagen gene. Mol. Cell. Biol. 1998, 18, 3368–3375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furger, A.; O’Sullivan, J.M.; Binnie, A.; Lee, B.A.; Proudfoot, N.J. Promoter proximal splice sites enhance transcription. Genes Dev. 2002, 16, 2792–2799. [Google Scholar] [CrossRef] [Green Version]
- Okkema, P.G.; Harrison, S.W.; Plunger, V.; Aryana, A.; Fire, A. Sequence requirements for myosin gene expression and regulation in Caenorhabditis elegans. Genetics 1993, 135, 385–404. [Google Scholar] [CrossRef]
- Moabbi, A.M.; Agarwal, N.; El Kaderi, B.; Ansari, A. Role for gene looping in intron-mediated enhancement of transcription. Proc. Natl. Acad. Sci. USA 2012, 109, 8505–8510. [Google Scholar] [CrossRef] [Green Version]
- Morello, L.; Giani, S.; Troina, F.; Breviario, D. Testing the IMEter on rice introns and other aspects of intron-mediated enhancement of gene expression. J. Exp. Bot. 2010, 62, 533–544. [Google Scholar] [CrossRef] [PubMed]
- Parra, G.; Bradnam, K.; Rose, A.B.; Korf, I. Comparative and functional analysis of intron-mediated enhancement signals reveals conserved features among plants. Nucleic Acids Res. 2011, 39, 5328–5337. [Google Scholar] [CrossRef] [Green Version]
- Liao, L.; Ning, G.; Liu, C.; Zhang, W.; Bao, M. The intron from the 5′-UTR of the FBP11 gene in petunia displays promoter- and enhancer-like functions. Sci. Hortic. 2013, 154, 96–101. [Google Scholar] [CrossRef]
- Nott, A.; Le Hir, H.; Moore, M.J. Splicing enhances translation in mammalian cells: An additional function of the exon junction complex. Genes Dev. 2004, 18, 210–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samadder, P.; Sivamani, E.; Lu, J.; Li, X.; Qu, R. Transcriptional and post-transcriptional enhancement of gene ex-pression by the 5′ UTR intron of rice rubi3 gene in transgenic rice cells. Mol. Genet. Genom. 2008, 279, 429–439. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Full Resistance | No Resistance | Partial Resistance |
|---|---|---|---|
| pBI121-N | 30/30 (100%) | 0/30 (0%) | 0/30 (0%) |
| pBI121-SRC7 | 30/30 (100%) | 0/30 (0%) | 0/30 (0%) |
| pBI121-GmRUN1 gDNA | 24/30 (80%) | 0/30 (0%) | 6/30 (20%) |
| pCB301-GmRUN1 cDNA | 0/30 (0%) | 13/30 (43%) | 17/30 (57%) |
| pBI121-GmRUN1 cDNA | 0/30 (0%) | 24/30 (80%) | 6/30 (20%) |
| pBI121 | 0/30 (0%) | 30/30 (100%) | 0/30 (0%) |
| Gene Name | HR/Total Leaves |
|---|---|
| pBI121-GmRUN1 gDNA | 14/30 (47%) |
| pCB301-GmRUN1 cDNA | 9/30 (30%) |
| pBI121-GmRUN1 cDNA | 7/30 (23%) |
| pCB301 | 0/30 (0%) |
| Gene Name | Full Resistance | No Resistance | Partial Resistance |
|---|---|---|---|
| pBI121-GmRUN1 gDNA | 17/25 (68%) | 0/25 (0%) | 8/25 (32%) |
| pCB301-GmRUN1TIR | 0/25 (0%) | 25/25 (100%) | 0/25 (0%) |
| pCB301-GmRUN1NBS | 0/25 (0%) | 25/25 (100%) | 0/25 (0%) |
| pCB301- GmRUN1LRR | 0/25 (0%) | 25/25 (100%) | 0/25 (0%) |
| pCB301-GmRUN1TN | 0/25 (0%) | 25/25 (100%) | 0/25 (0%) |
| pCB301-GmRUN1NL | 0/25 (0%) | 25/25 (100%) | 0/25 (0%) |
| pCB301-GmRUN1TNL | 0/25 (0%) | 6/25 (24%) | 19/25 (76%) |
| pCB301 | 0/25 (0%) | 25/25 (100%) | 0/25 (0%) |
| Gene Name | HR/Total Leaves |
|---|---|
| pBI121-GmRUN1 gDNA | 10/25 (40%) |
| pCB301-GmRUN1TIR | 1/25 (4%) |
| pCB301-GmRUN1NBS | 0/25 (0%) |
| pCB301-GmRUN1LRR | 0/25 (0%) |
| pCB301-GmRUN1TN | 1/25 (4%) |
| pCB301-GmRUN1NL | 0/25 (0%) |
| pCB301-GmRUN1TNL | 6/25 (24%) |
| pCB301 | 0/25 (0%) |
| Gene Name | Full Resistance | No Resistance | Partial Resistance |
|---|---|---|---|
| pCambia1300-SRC7-intron2 | 15/20 (75%) | 0/20 (0%) | 5/20 (25%) |
| pCambia1300-SRC7 | 7/20 (35%) | 0/20 (0%) | 13/20 (65%) |
| pCambia1300-N | 8/20 (40%) | 0/20 (0%) | 12/20 (60%) |
| pCambia1300 | 0/20 (0%) | 20/20 (100%) | 0/20 (0%) |
| Gene Name | Full Resistance | No Resistance | Partial Resistance |
|---|---|---|---|
| pCB301-gDNA | 9/35 (26%) | 0/35 (0%) | 26/35 (74%) |
| pCB301-GmRUN1-ExIn-1 | 15/35 (43%) | 0/35 (0%) | 20/35 (57%) |
| pCB301-GmRUN1-ExIn-5 | 2/35 (6%) | 0/35 (0%) | 33/35 (94%) |
| pCB301-GmRUN1-ExIn-11 | 2/35 (6%) | 5/35 (14%) | 28/35 (80%) |
| pCB301-cDNA | 0/35 (0%) | 17/35 (49%) | 18/35 (51%) |
| pCB301 | 0/35 (0%) | 35/35 (100%) | 0/35 (0%) |
| Promoter Element | Sequence | Function |
|---|---|---|
| TATA-box | TATAA | Core promoter element around –30 of transcription start |
| CAAT-box | CCAAT | Common cis-acting element in promoter and enhancer regions |
| CGTCA-motif | CGTCA | Cis-acting regulatory element involved in the MeJA-responsiveness |
| TGACG-motif | TGACG | Cis-acting regulatory element involved in the MeJA-responsiveness |
| as-1 | TGACG | Cis-acting element related to salicylic acid induction |
| TCA-element | TCAGAAGAGG | Cis-acting element involved in salicylic acid responsiveness |
| TCA | TCATCTTCAT | Unknown functional element |
| ABRE | ACGTG | Cis-acting element involved in the abscisic acid responsiveness |
| ERE | ATTTTAAA | Cis-acting element involved in ethylene response |
| ARE | AAACCA | Cis-acting regulatory element essential for the anaerobic induction |
| LTR | CCGAAA | Cis-acting element involved in low-temperature responsiveness |
| MBS | CAACTG | MYB binding site involved in drought inducibility |
| W-box | TTGACC | Cis-acting element involved in disease resistance |
| CAT-box | GCCACT | Cis-acting regulatory element related to meristem expression |
| AT1-motif | AATTATTTTTTATT | Part of a light-responsive module |
| GT1-motif | GGTTAAT | Light-responsive element |
| TCT-motif | TCTTAC | Part of a light-responsive element |
| G-Box | CACGTG | Cis-acting regulatory element involved in light responsiveness |
| MRE | AACCTAA | MYB binding site involved in light responsiveness |
| Box 4 | ATTAAT | Part of a conserved DNA module involved in light responsiveness |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diao, P.; Sun, H.; Bao, Z.; Li, W.; Niu, N.; Li, W.; Wuriyanghan, H. Expression of an Antiviral Gene GmRUN1 from Soybean Is Regulated via Intron-Mediated Enhancement (IME). Viruses 2021, 13, 2032. https://0-doi-org.brum.beds.ac.uk/10.3390/v13102032
Diao P, Sun H, Bao Z, Li W, Niu N, Li W, Wuriyanghan H. Expression of an Antiviral Gene GmRUN1 from Soybean Is Regulated via Intron-Mediated Enhancement (IME). Viruses. 2021; 13(10):2032. https://0-doi-org.brum.beds.ac.uk/10.3390/v13102032
Chicago/Turabian StyleDiao, Pengfei, Hongyu Sun, Zhuo Bao, Wenxia Li, Niu Niu, Weimin Li, and Hada Wuriyanghan. 2021. "Expression of an Antiviral Gene GmRUN1 from Soybean Is Regulated via Intron-Mediated Enhancement (IME)" Viruses 13, no. 10: 2032. https://0-doi-org.brum.beds.ac.uk/10.3390/v13102032