Retroviral Antisense Transcripts and Genes: 33 Years after First Predicted, a Silent Retroviral Revolution?

 and
and 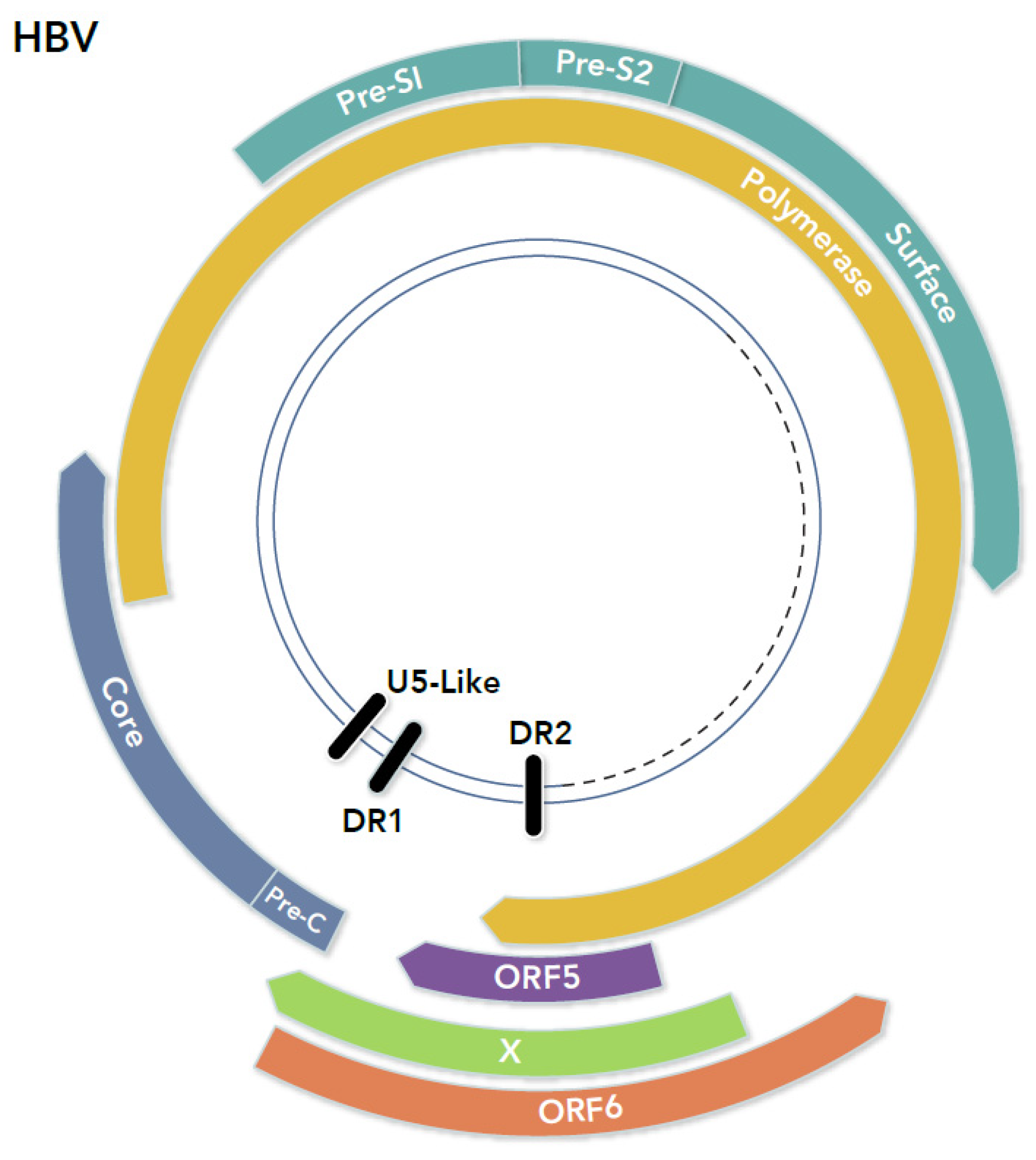{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Virology: The Birth of a Discipline
3. Retrovirology: A Cornucopia for Understanding Oncogenesis
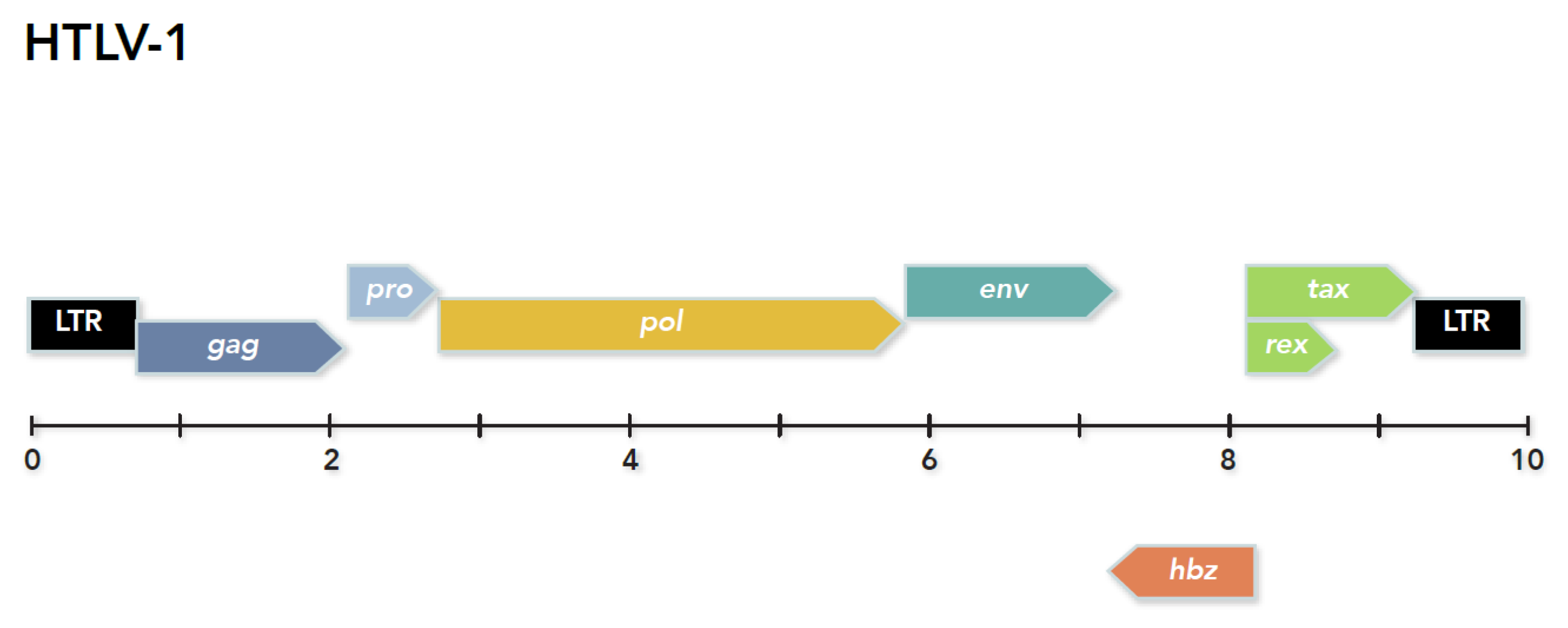
4. Discovery of the First Human Retrovirus: HTLV-1, Birth of Human Retrovirology
5. Antisense Transcription and Antisense Proteins in Retroviruses: A Significant Hypothesis
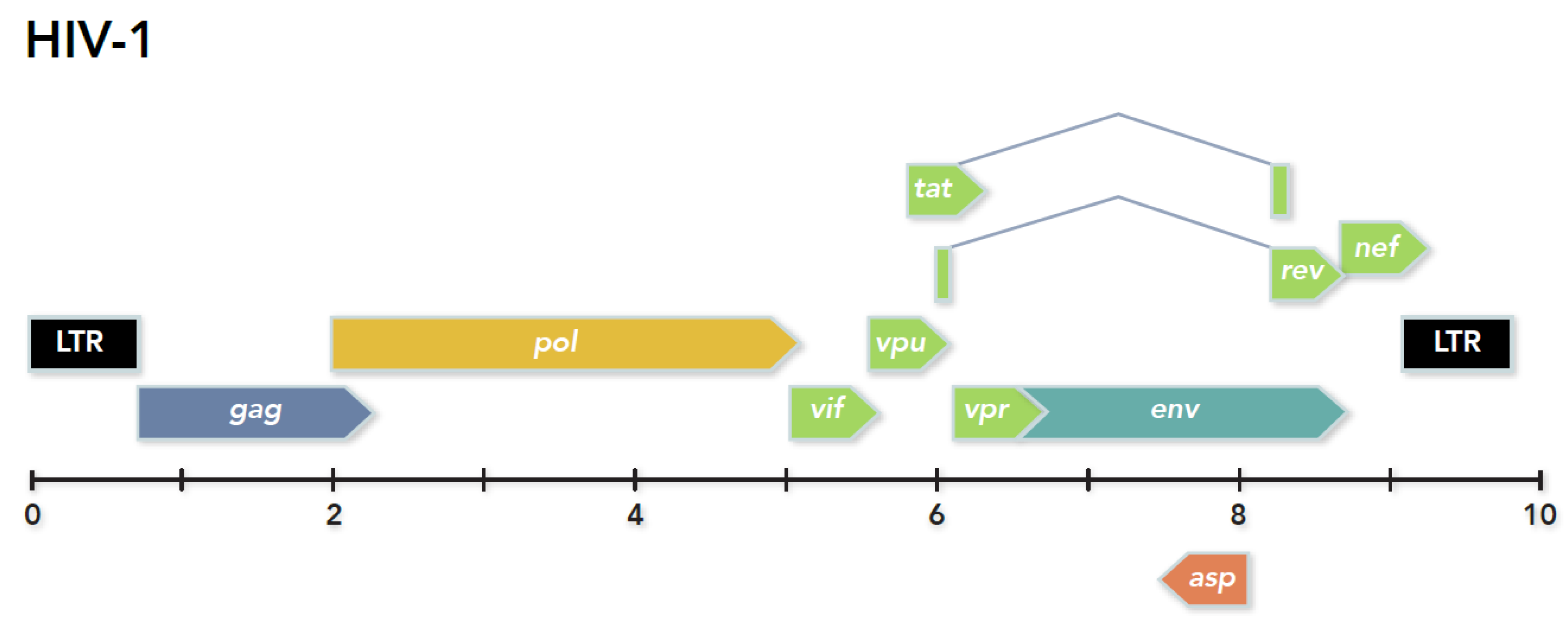
6. The 10th Gene of HIV-1 Encodes a Protein Expressed from Antisense mRNA: A Shift in Perspective
6.1. Predicting the Existence of an HIV-1 Antisense Protein
6.2. Lessons from the Hepadnavirus Genome
6.3. Formulation of the Hypothesis for an HIV-1 Antisense Protein
6.4. Additional Smaller Antisense Transcripts and Proteins in HIV-1
7. Antisense Transcripts and/or ORFs in the 3′ Genome Region of Other Exogenous Retroviruses and in Endogenous Human Retroviruses
7.1. Animal Lentiviruses
7.2. HTLV-1
7.3. Other Human and Animal Deltaretroviruses
7.4. Gammaretroviruses
7.5. Endogenous Human Retroviruses
7.6. Hepadnaviruses
8. Barriers to Overcome in the Study of HIV-1 ASP: From Technical Difficulties to Epistemological Blocks
8.1. Technical Difficulties
8.2. Epistemological Blocks
9. Summary and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pradeu, T.; Kostyrka, G.; Dupré, J. Understanding Viruses: Philosophical Investigations. Stud. Hist. Philos. Science Part C Stud. Hist. Philos. Biol. Biomed. Sci. 2016, 59, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Hendrix, R.W. Bacteriophages: Evolution of the Majority. Theor. Popul. Biol. 2002, 61, 471–480. [Google Scholar] [CrossRef]
- Wasik, B.R.; Turner, P.E. On the Biological Success of Viruses. Annu. Rev. Microbiol. 2013, 67, 519–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bar-On, Y.M.; Phillips, R.; Milo, R. The Biomass Distribution on Earth. Proc. Natl. Acad. Sci. USA 2018, 115, 6506–6511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohwer, F.; Barott, K. Viral Information. Biol. Philos. 2013, 28, 283–297. [Google Scholar] [CrossRef] [Green Version]
- Pasteur, L. La Théorie Des Germes et Ses Applications à La Médecine et à La Chirurgie, 1st ed.; Académie de médecine: Paris, France, 1878. [Google Scholar]
- Ivanovski, D.I. Über Die Mosaikkrankheit Der Tabakspflanze (Lu Le 12 Février 1892). Bull. Acad. Imp. Sci. St.-Pétersbg. 1894, 35, 67–70. [Google Scholar]
- Beijerinck, M.V. Over Een Contagium Vivum Fluidum Als Oorjaak van de Vlekziekte Der Tabaksbladen. Versl Gewone Vergad Wis En Natnurk. Afa. K. Akad. Wet. Amst. 1898, 7, 229–235. [Google Scholar]
- De Chauliac, G. Le Guidon en François, 1st ed.; B. Buyer: Lyon, France, 1478; pp. 79–423. [Google Scholar]
- Paré, A. Oeuvres Complètes d’Ambroise Paré; Librarie de l’Academie Royale de Medecine: Paris, France, 1575. [Google Scholar]
- Roux, E. Sur Les Microbes Dits «invisibles». Bull. Inst. Pasteur 1903, 1, 7–12/49–56. [Google Scholar]
- Loeffler, F.; Frosch, P. Berichte Der Komission Zur Erforschung Der Maul Und Klauenseuche Bei Dem Institut Für Infectios-Krankheiten in Berlin. Ztbl. Bakt. Parasitkde 1898, 23, 371–391. [Google Scholar]
- Reed, W.; Carroll, J.; Agramonte, A.; Lazear, W. The Etiology of Yellow Fever: A Premilary Note. Phila. Med. 1901, 6, 790–796. [Google Scholar]
- Rous, P.A. Sarcoma of the Fowl Transmissible by an Agent Separable from the Tumor Cells. J. Exp. Med. 1911, 13, 397–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Twort, F.W. An Investigation on the Nature of the Ultramicroscopic Viruses. Lancet 1915, 2, 1241–1243. [Google Scholar] [CrossRef] [Green Version]
- D’Herelle, F. Sur Un Microbe Invisible Antagoniste Des Bacilles Dysentériques. CR Acad. Sci. Paris 1917, 165, 373–375. [Google Scholar]
- Andrewes, C.H.; Laidlaw, P.P.; Smith, W. The Susceptibiliy of Mice to the Viruses of Human and Swine Influenza. Lancet 1934, 2, 859–862. [Google Scholar] [CrossRef]
- Kausche, G.A.; Pfankuch, E.; Ruska, H. Die Sichtbarmachung von Planzlichen Virus Im Ubermikroskop. Naturwissenschaften 1939, 27, 292–299. [Google Scholar] [CrossRef]
- Bernal, J.D. Fankuchen I X-ray and Crystallographic Studies of Plant Virus Preparations. J. Gen. Physiol. 1941, 25, 111–165. [Google Scholar] [CrossRef] [Green Version]
- Ellis, E.L.; Delbrück, M. The Growth of Bacteriophage. J. Gen. Physiol. 1939, 22, 365–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luria, S.E.; Anderson, T.F. The Identification and Characterization of Bacteriophages with the Electron Microscope. Proc. Natl. Acad. Sci. USA 1942, 28, 127–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luria, S.E. Bacteriophage: An Assay on Virus Reproduction. Science 1950, 111, 507–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enders, J.F.; Weller, T.H.; Robbins, F.C. Cultivation of the Lansing Strain of Poliomyelitis Virus in Cultures of Various Human Embryonic Tissues. Science 1949, 109, 85–87. [Google Scholar] [CrossRef]
- Lwoff, A. The Concept of Virus. J. Gen. Microbiol. 1957, 17, 239–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lwoff, A.; Tournier, P. The Classification of Viruses. Ann. Rev. Microbiol. 1966, 20, 45–74. [Google Scholar] [CrossRef]
- Vallée, H.; Carré, H. Sur La Nature Infectieuse de l’anémie Du Cheval. Comptes Rendus. 1904, 139, 331–333. [Google Scholar]
- Ellermann, V.u.O. Bang Experimentelle Leukämie Bei Hühnern. Zentralbl. Bakteriol. Parasitenkd. Infectionskr. Hyg. Abt. Orig. 1908, 46, 595–609. [Google Scholar]
- Claude, A.; Murphy, J.G. Transmissible Tumors of the Fowl. Physiol. Rev. 1933, 13, 246–275. [Google Scholar] [CrossRef]
- Bittner, J.J. Some Possible Effects of Nursing on the Mammary Gland Tumor Incidence in Mice. Science 1936, 84, 162. [Google Scholar] [CrossRef]
- Gross, L. Development and Serial Cell-Free Passage of a Highly Potent Strain of Mouse Leukemia Virus. Exp. Biol. Med. 1957, 94, 767–771. [Google Scholar] [CrossRef]
- Temin, H.M. The effects of actinomycin d on growth of rous sarcoma virus in vitro. Virology 1963, 20, 577–582. [Google Scholar] [CrossRef]
- Bader, J.P. Transformation of rous sarcoma virus: A requirement of dna synthesis. Science 1965, 149, 757–758. [Google Scholar] [CrossRef]
- Temin, H.M. The participation of dna in rous sarcoma virus production. Virology 1964, 23, 486–494. [Google Scholar] [CrossRef]
- Baltimore, D. RNA-Dependent DNA Polymerase in Virions of RNA Tumour Viruses. Nature 1970, 226, 1209–1211. [Google Scholar] [CrossRef]
- Mizutani, S.; Boettiger, D.; Temin, H.M. A DNA-Depenent DNA Polymerase and a DNA Endonuclease in Virions of Rous Sarcoma Virus. Nature 1970, 228, 424–427. [Google Scholar] [CrossRef] [PubMed]
- Spiegelman, S.; Burny, A.; Das, M.R.; Keydar, J.; Schlom, J.; Travnicek, M.; Watson, K. Characterization of the Products of DNA-Directed DNA Polymerases in Oncogenic RNA Viruses. Nature 1970, 227, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Martin, G.S. Rous Sarcoma Virus: A Function Required for the Maintenance of the Transformed State. Nature 1970, 227, 1021–1023. [Google Scholar] [CrossRef] [PubMed]
- Duesberg, P.H.; Vogt, P.K. Differences between the Ribonucleic Acids of Transforming and Nontransforming Avian Tumor Viruses. Proc. Natl. Acad. Sci. USA 1970, 67, 1673–1680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.H.; Duesberg, P.H.; Kawai, S.; Hanafusa, H. Location of Envelope-Specific and Sarcoma-Specific Oligonucleotides on RNA of Schmidt-Ruppin Rous Sarcoma Virus. Proc. Natl. Acad. Sci. USA 1976, 73, 447–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duesberg, P.H.; Bister, K.; Vogt, P.K. The RNA of Avian Acute Leukemia Virus MC29. Proc. Natl. Acad. Sci. USA 1977, 74, 4320–4324. [Google Scholar] [CrossRef] [Green Version]
- Duesberg, P.H.; Vogt, P.K. Avian Acute Leukemia Viruses MC29 and MH2 Share Specific RNA Sequences: Evidence for a Second Class of Transforming Genes. Proc. Natl. Acad. Sci. USA 1979, 76, 1633–1637. [Google Scholar] [CrossRef] [Green Version]
- Karnoub, A.E.; Weinberg, R.A. Ras Oncogenes: Split Personalities. Nat. Rev. Mol. Cell Biol. 2008, 9, 517–531. [Google Scholar] [CrossRef] [Green Version]
- Uchiyama, T.; Yodoi, J.; Sagawa, K.; Takatsuki, K.; Uchino, H. Adult T-Cell Leukemia: Clinical and Hematologic Features of 16 Cases. Blood 1977, 50, 481–492. [Google Scholar] [CrossRef] [Green Version]
- Poiesz, B.J.; Ruscetti, F.W.; Gazdar, A.F.; Bunn, P.A.; Minna, J.D.; Gallo, R.C. Detection and Isolation of Type C Retrovirus Particles from Fresh and Cultured Lymphocytes of a Patient with Cutaneous T-Cell Lymphoma. Proc. Natl. Acad. Sci. USA 1980, 77, 7415–7419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gessain, A.; Barin, F.; Vernant, J.C.; Gout, O.; Maurs, L.; Calender, A.; de Thé, G. Antibodies to Human T-Lymphotropic Virus Type-I in Patients with Tropical Spastic Paraparesis. Lancet 1985, 2, 407–410. [Google Scholar] [CrossRef]
- Matsuoka, M.; Jeang, K.-T. Human T-Cell Leukaemia Virus Type 1 (HTLV-1) Infectivity and Cellular Transformation. Nat. Rev. Cancer 2007, 7, 270–280. [Google Scholar] [CrossRef]
- Marriott, S.J.; Semmes, O.J. Impact of HTLV-I Tax on Cell Cycle Progression and the Cellular DNA Damage Repair Response. Oncogene 2005, 24, 5986–5995. [Google Scholar] [CrossRef] [PubMed]
- Boxus, M.; Willems, L. Mechanisms of HTLV-1 Persistence and Transformation. Br. J. Cancer 2009, 101, 1497–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Currer, R.; Van Duyne, R.; Jaworski, E.; Guendel, I.; Sampey, G.; Das, R.; Narayanan, A.; Kashanchi, F. HTLV Tax: A Fascinating Multifunctional Co-Regulator of Viral and Cellular Pathways. Front. Microbiol. 2012, 3, 406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romanelli, M.G.; Diani, E.; Bergamo, E.; Casoli, C.; Ciminale, V.; Bex, F.; Bertazzoni, U. Highlights on Distinctive Structural and Functional Properties of HTLV Tax Proteins. Front. Microbiol. 2013, 4, 271. [Google Scholar] [CrossRef] [Green Version]
- Mohanty, S.; Harhaj, E.W. Mechanisms of Oncogenesis by HTLV-1 Tax. Pathogens 2020, 9, 543. [Google Scholar] [CrossRef]
- Akkouche, A.; Moodad, S.; Hleihel, R.; Skayneh, H.; Chambeyron, S.; El Hajj, H.; Bazarbachi, A. In Vivo Antagonistic Role of the Human T-Cell Leukemia Virus Type 1 Regulatory Proteins Tax and HBZ. PLoS Pathog. 2021, 17, e1009219. [Google Scholar] [CrossRef] [PubMed]
- Spiegelman, W.G.; Reichardt, L.F.; Yaniv, M.; Heinemann, S.F.; Kaiser, A.D.; Eisen, H. Bidirectional Transcription and the Regulation of Phage Lambda Repressor Synthesis. Proc. Natl. Acad. Sci. USA 1972, 69, 3156–3160. [Google Scholar] [CrossRef] [Green Version]
- Savoret, J.; Mesnard, J.-M.; Gross, A.; Chazal, N. Antisense Transcripts and Antisense Protein: A New Perspective on Human Immunodeficiency Virus Type 1. Front. Microbiol. 2021, 11, 625941. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.H. Human Immunodeficiency Virus May Encode a Novel Protein on the Genomic DNA plus Strand. Science 1988, 239, 1420–1422. [Google Scholar] [CrossRef]
- Beadle, G.W.; Tatum, E.L. Genetic Control of Biochemical Reactions in Neurospora. Proc. Natl. Acad. Sci. USA 1941, 27, 499–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horowitz, N.H. The One Gene-One Enzyme Hypothesis. Genetics 1948, 33, 612. [Google Scholar] [PubMed]
- Miller, R.H.; Kaneko, S.; Chung, C.T.; Girones, R.; Purcell, R.H. Compact Organization of the Hepatitis B Virus Genome. Hepatology 1989, 9, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Galibert, F.; Mandart, E.; Fitoussi, F.; Tiollais, P.; Charnay, P. Nucleotide Sequence of the Hepatitis B Virus Genome (Subtype Ayw) Cloned in E. Coli. Nature 1979, 281, 646–650. [Google Scholar] [CrossRef]
- Miller, R.H. Organization of the X Gene Region of the Hepatitis B Virus Genome. Gastroenterol. Jpn. 1990, 25, 1–5. [Google Scholar] [CrossRef]
- Summers, J.; Mason, W.S. Replication of the Genome of a Hepatitis B-like Virus by Reverse Transcription of an RNA Intermediate. Cell 1982, 29, 403–415. [Google Scholar] [CrossRef]
- Miller, R.H.; Marion, P.L.; Robinson, W.S. Hepatitis B Viral DNA-RNA Hybrid Molecules in Particles from Infected Liver Are Converted to Viral DNA Molecules during an Endogenous Dna Polymerase Reaction. Virology 1984, 139, 64–72. [Google Scholar] [CrossRef]
- Miller, R.H.; Robinson, W.S. Common Evolutionary Origin of Hepatitis B Virus and Retroviruses. Proc. Natl. Acad. Sci. USA 1986, 83, 2531–2535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, R.H. Evolutionary Relationship between Hepadna Viruses and Retroviruses; CRC Press: Boca Raton, FL, USA, 1991; pp. 227–244. [Google Scholar]
- Cassan, E.; Arigon-Chifolleau, A.-M.; Mesnard, J.-M.; Gross, A.; Gascuel, O. Concomitant Emergence of the Antisense Protein Gene of HIV-1 and of the Pandemic. Proc. Natl. Acad. Sci. USA 2016, 113, 11537–11542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimonte, S. Different HIV-1 Env Frames: Gp120 and ASP (Antisense Protein) Biosynthesis, and Theirs Co-Variation Tropic Amino Acid Signatures in X4- and R5-Viruses: HIV-1 Antisense Protein and Tropic Co-Variation with Gp120 V3 Signatures. J. Med. Virol. 2017, 89, 112–122. [Google Scholar] [CrossRef]
- Nelson, C.W.; Ardern, Z.; Wei, X. OLGenie: Estimating Natural Selection to Predict Functional Overlapping Genes. Mol. Biol. Evol. 2020, 27, 2440–2449. [Google Scholar] [CrossRef]
- Bukrinsky, M.I.; Etkin, A.F. Plus Strand of the HIV Provirus DNA Is Expressed at Early Stages of Infection. AIDS Res. Hum. Retrovir. 1990, 6, 425–426. [Google Scholar] [CrossRef] [PubMed]
- Michael, N.L.; Vahey, M.T.; d’Arcy, L.; Ehrenberg, P.K.; Mosca, J.D.; Rappaport, J.; Redfield, R.R. Negative-Strand RNA Transcripts Are Produced in Human Immunodeficiency Virus Type 1-Infected Cells and Patients by a Novel Promoter Downregulated by Tat. J. Virol. 1994, 68, 979–987. [Google Scholar] [CrossRef] [Green Version]
- Peeters, A.; Lambert, P.F.; Deacon, N.J. A Fourth Sp1 Site in the Human Immunodeficiency Virus Type 1 Long Terminal Repeat Is Essential for Negative-Sense Transcription. J. Virol. 1996, 70, 6665–6672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig, L.B.; Ambrus, J.L.; Krawczyk, K.A.; Sharma, S.; Brooks, S.; Hsiao, C.-B.; Schwartz, S.A. Human Immunodeficiency Virus-Type 1 LTR DNA Contains an Intrinsic Gene Producing Antisense RNA and Protein Products. Retrovirology 2006, 3, 80. [Google Scholar] [CrossRef] [Green Version]
- Landry, S.; Halin, M.; Lefort, S.; Audet, B.; Vaquero, C.; Mesnard, J.-M.; Barbeau, B. Detection, Characterization and Regulation of Antisense Transcripts in HIV-1. Retrovirology 2007, 4, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laverdure, S.; Gross, A.; Arpin-André, C.; Clerc, I.; Beaumelle, B.; Barbeau, B.; Mesnard, J.-M. HIV-1 Antisense Transcription Is Preferentially Activated in Primary Monocyte-Derived Cells. J. Virol. 2012, 86, 13785–13789. [Google Scholar] [CrossRef] [Green Version]
- Mancarella, A.; Procopio, F.A.; Achsel, T.; De Crignis, E.; Foley, B.T.; Corradin, G.; Bagni, C.; Pantaleo, G.; Graziosi, C. Detection of Antisense Protein (ASP) RNA Transcripts in Individuals Infected with Human Immunodeficiency Virus Type 1 (HIV-1). J. Gen. Virol. 2019, 100, 863–876. [Google Scholar] [CrossRef] [PubMed]
- Barbagallo, M.S.; Birch, K.E.; Deacon, N.J.; Mosse, J.A. Potential Control of Human Immunodeficiency Virus Type 1 Asp. Expression by Alternative Splicing in the Upstream Untranslated Region. DNA Cell Biol. 2012, 31, 1303–1313. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi-Ishihara, M.; Yamagishi, M.; Hara, T.; Matsuda, Y.; Takahashi, R.; Miyake, A.; Nakano, K.; Yamochi, T.; Ishida, T.; Watanabe, T. HIV-1-Encoded Antisense RNA Suppresses Viral Replication for a Prolonged Period. Retrovirology 2012, 9, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zapata, J.C.; Campilongo, F.; Barclay, R.A.; DeMarino, C.; Iglesias-Ussel, M.D.; Kashanchi, F.; Romerio, F. The Human Immunodeficiency Virus 1 ASP RNA Promotes Viral Latency by Recruiting the Polycomb Repressor Complex 2 and Promoting Nucleosome Assembly. Virology 2017, 506, 34–44. [Google Scholar] [CrossRef]
- Saayman, S.; Ackley, A.; Turner, A.-M.W.; Famiglietti, M.; Bosque, A.; Clemson, M.; Planelles, V.; Morris, K.V. An HIV-Encoded Antisense Long Noncoding RNA Epigenetically Regulates Viral Transcription. Mol. Ther. 2014, 22, 1164–1175. [Google Scholar] [CrossRef] [Green Version]
- Ma, G.; Yasunaga, J.-I.; Shimura, K.; Takemoto, K.; Watanabe, M.; Amano, M.; Nakata, H.; Liu, B.; Zuo, X.; Matsuoka, M. Human Retroviral Antisense MRNAs Are Retained in the Nuclei of Infected Cells for Viral Persistence. Proc. Natl. Acad. Sci. USA 2021, 118, e2014783118. [Google Scholar] [CrossRef]
- Kobayashi-Ishihara, M.; Terahara, K.; Martinez, J.P.; Yamagishi, M.; Iwabuchi, R.; Brander, C.; Ato, M.; Watanabe, T.; Meyerhans, A.; Tsunetsugu-Yokota, Y. HIV LTR-Driven Antisense RNA by Itself Has Regulatory Function and May Curtail Virus Reactivation from Latency. Front. Microbiol. 2018, 9, 1066. [Google Scholar] [CrossRef] [Green Version]
- Tagieva, N.E.; Vaquero, C. Expression of Naturally Occurring Antisense RNA Inhibits Human Immunodeficiency Virus Type 1 Heterologous Strain Replication. J. Gen. Virol. 1997, 78, 2503–2511. [Google Scholar] [CrossRef] [Green Version]
- Torresilla, C.; Mesnard, J.-M.; Barbeau, B. Reviving an Old HIV-1 Gene: The HIV-1 Antisense Protein. CHR 2015, 13, 117–124. [Google Scholar] [CrossRef]
- Li, R.; Sklutuis, R.; Groebner, J.L.; Romerio, F. HIV-1 Natural Antisense Transcription and Its Role in Viral Persistence. Viruses 2021, 13, 795. [Google Scholar] [CrossRef] [PubMed]
- Vanhée-Brossollet, C.; Thoreau, H.; Serpente, N.; D’Auriol, L.; Lévy, J.P.; Vaquero, C. A Natural Antisense RNA Derived from the HIV-1 Env Gene Encodes a Protein Which Is Recognized by Circulating Antibodies of HIV+ Individuals. Virology 1995, 206, 196–202. [Google Scholar] [CrossRef] [Green Version]
- Savoret, J.; Chazal, N.; Moles, J.-P.; Tuaillon, E.; Boufassa, F.; Meyer, L.; Lecuroux, C.; Lambotte, O.; Van De Perre, P.; Mesnard, J.-M.; et al. A Pilot Study of the Humoral Response Against the AntiSense Protein (ASP) in HIV-1-Infected Patients. Front. Microbiol. 2020, 11, 20. [Google Scholar] [CrossRef]
- Bansal, A.; Carlson, J.; Yan, J.; Akinsiku, O.T.; Schaefer, M.; Sabbaj, S.; Bet, A.; Levy, D.N.; Heath, S.; Tang, J.; et al. CD8 T Cell Response and Evolutionary Pressure to HIV-1 Cryptic Epitopes Derived from Antisense Transcription. J. Exp. Med. 2010, 207, 51–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bet, A.; Maze, E.A.; Bansal, A.; Sterrett, S.; Gross, A.; Graff-Dubois, S.; Samri, A.; Guihot, A.; Katlama, C.; Theodorou, I.; et al. The HIV-1 Antisense Protein (ASP) Induces CD8 T Cell Responses during Chronic Infection. Retrovirology 2015, 12, 15. [Google Scholar] [CrossRef] [PubMed]
- Berger, C.T.; Llano, A.; Carlson, J.M.; Brumme, Z.L.; Brockman, M.A.; Cedeño, S.; Harrigan, P.R.; Kaufmann, D.E.; Heckerman, D.; Meyerhans, A.; et al. Immune Screening Identifies Novel T Cell Targets Encoded by Antisense Reading Frames of HIV-1. J. Virol. 2015, 89, 4015–4019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briquet, S.; Vaquero, C. Immunolocalization Studies of an Antisense Protein in HIV-1-Infected Cells and Viral Particles. Virology 2002, 292, 177–184. [Google Scholar] [CrossRef] [Green Version]
- Clerc, I.; Laverdure, S.; Torresilla, C.; Landry, S.; Borel, S.; Vargas, A.; Arpin-André, C.; Gay, B.; Briant, L.; Gross, A.; et al. Polarized Expression of the Membrane ASP Protein Derived from HIV-1 Antisense Transcription in T Cells. Retrovirology 2011, 8, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torresilla, C.; Larocque, É.; Landry, S.; Halin, M.; Coulombe, Y.; Masson, J.-Y.; Mesnard, J.-M.; Barbeau, B. Detection of the HIV-1 Minus-Strand-Encoded Antisense Protein and Its Association with Autophagy. J. Virol. 2013, 87, 5089–5105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Torresilla, C.; Xiao, Y.; Nguyen, P.T.; Caté, C.; Barbosa, K.; Rassart, É.; Cen, S.; Bourgault, S.; Barbeau, B. HIV-1 Antisense Protein of Different Clades Induces Autophagy and Associates with the Autophagy Factor P62. J. Virol. 2018, 93, e01757-18. [Google Scholar] [CrossRef] [Green Version]
- Affram, Y.; Zapata, J.C.; Gholizadeh, Z.; Tolbert, W.D.; Zhou, W.; Iglesias-Ussel, M.D.; Pazgier, M.; Ray, K.; Latinovic, O.S.; Romerio, F. The HIV-1 Antisense Protein ASP Is a Transmembrane Protein of the Cell Surface and an Integral Protein of the Viral Envelope. J. Virol. 2019, 93, e00574-19. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, L.; Albert, M. MORT, a Locus for Apoptosis in the Human Immunodeficiency Virus-Type 1 Antisense Gene: Implications for AIDS, Cancer, and Covid-19. bioRixv 2020, in press. [Google Scholar]
- Ludwig, L.B. RNA Silencing and HIV: A Hypothesis for the Etiology of the Severe Combined Immunodeficiency Induced by the Virus. Retrovirology 2008, 5, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Zhao, X.; Shen, W.; Kong, X. Evidence for the Antisense Transcription in the Proviral R29-127 Strain of Bovine Immunodeficiency Virus. Virol. Sin. 2015, 30, 224–227. [Google Scholar] [CrossRef] [PubMed]
- Briquet, S.; Richardson, J.; Vanhée-Brossollet, C.; Vaquero, C. Natural Antisense Transcripts Are Detected in Different Cell Lines and Tissues of Cats Infected with Feline Immunodeficiency Virus. Gene 2001, 267, 157–164. [Google Scholar] [CrossRef]
- Miller, C.; Abdo, Z.; Ericsson, A.; Elder, J.; VandeWoude, S. Applications of the FIV Model to Study HIV Pathogenesis. Viruses 2018, 10, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larocca, D.; Chao, L.A.; Seto, M.H.; Brunck, T.K. Human T-Cell Leukemia Virus Minus Strand Transcription in Infected T-Cells. Biochem. Biophys. Res. Commun. 1989, 163, 1006–1013. [Google Scholar] [CrossRef]
- Gaudray, G.; Gachon, F.; Basbous, J.; Biard-Piechaczyk, M.; Devaux, C.; Mesnard, J.-M. The Complementary Strand of the Human T-Cell Leukemia Virus Type 1 RNA Genome Encodes a BZIP Transcription Factor That Down-Regulates Viral Transcription. J. Virol. 2002, 76, 12813–12822. [Google Scholar] [CrossRef] [Green Version]
- Barbeau, B.; Mesnard, J.-M. Does Chronic Infection in Retroviruses Have a Sense? Trends Microbiol. 2015, 23, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Satou, Y.; Yasunaga, J.-I.; Zhao, T.; Yoshida, M.; Miyazato, P.; Takai, K.; Shimizu, K.; Ohshima, K.; Green, P.L.; Ohkura, N.; et al. HTLV-1 BZIP Factor Induces T-Cell Lymphoma and Systemic Inflammation in Vivo. PLoS Pathog. 2011, 7, e1001274. [Google Scholar] [CrossRef]
- Ma, G.; Yasunaga, J.-I.; Matsuoka, M. Multifaceted Functions and Roles of HBZ in HTLV-1 Pathogenesis. Retrovirology 2016, 13, 16. [Google Scholar] [CrossRef] [Green Version]
- Miyazato, P.; Matsuo, M.; Katsuya, H.; Satou, Y. Transcriptional and Epigenetic Regulatory Mechanisms Affecting HTLV-1 Provirus. Viruses 2016, 8, 171. [Google Scholar] [CrossRef] [Green Version]
- Matsuoka, M.; Mesnard, J.-M. HTLV-1 BZIP Factor: The Key Viral Gene for Pathogenesis. Retrovirology 2020, 17, 2. [Google Scholar] [CrossRef]
- Arnold, J.; Zimmerman, B.; Li, M.; Lairmore, M.D.; Green, P.L. Human T-Cell Leukemia Virus Type-1 Antisense-Encoded Gene, Hbz, Promotes T-Lymphocyte Proliferation. Blood 2008, 112, 3788–3797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sintasath, D.M.; Wolfe, N.D.; Zheng, H.; LeBreton, M.; Peeters, M.; Tamoufe, U.; Djoko, C.F.; Diffo, J.L.; Mpoudi-Ngole, E.; Heneine, W.; et al. Genetic Characterization of the Complete Genome of a Highly Divergent Simian T-Lymphotropic Virus (STLV) Type 3 from a Wild Cercopithecus Mona Monkey. Retrovirology 2009, 6, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durkin, K.; Rosewick, N.; Artesi, M.; Hahaut, V.; Griebel, P.; Arsic, N.; Burny, A.; Georges, M.; Van den Broeke, A. Characterization of Novel Bovine Leukemia Virus (BLV) Antisense Transcripts by Deep Sequencing Reveals Constitutive Expression in Tumors and Transcriptional Interaction with Viral MicroRNAs. Retrovirology 2016, 13, 33. [Google Scholar] [CrossRef] [Green Version]
- Halin, M.; Douceron, E.; Clerc, I.; Journo, C.; Ko, N.L.; Landry, S.; Murphy, E.L.; Gessain, A.; Lemasson, I.; Mesnard, J.-M.; et al. Human T-Cell Leukemia Virus Type 2 Produces a Spliced Antisense Transcript Encoding a Protein That Lacks a Classic BZIP Domain but Still Inhibits Tax2-Mediated Transcription. Blood 2009, 114, 2427–2438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbeau, B.; Peloponese, J.-M.; Mesnard, J.-M. Functional Comparison of Antisense Proteins of HTLV-1 and HTLV-2 in Viral Pathogenesis. Front. Microbiol. 2013, 4, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panfil, A.R.; Dissinger, N.J.; Howard, C.M.; Murphy, B.M.; Landes, K.; Fernandez, S.A.; Green, P.L. Functional Comparison of HBZ and the Related APH-2 Protein Provides Insight into Human T-Cell Leukemia Virus Type 1 Pathogenesis. J. Virol. 2016, 90, 3760–3772. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, M.H.; Ballarín-González, B.; Liu, J.; Lassen, L.B.; Füchtbauer, A.; Füchtbauer, E.-M.; Nielsen, A.L.; Pedersen, F.S. Antisense Transcription in Gammaretroviruses as a Mechanism of Insertional Activation of Host Genes. J. Virol. 2010, 84, 3780–3788. [Google Scholar] [CrossRef] [Green Version]
- Robinson, W.S.; Miller, R.H.; Marion, P.L. Hepadnaviruses and Retroviruses Share Genome Homology and Features of Replication. Hepatology 1987, 7, 64S–73S. [Google Scholar] [CrossRef]
- Villesen, P.; Aagaard, L.; Wiuf, C.; Pedersen, F.S. Identification of Endogenous Retroviral Reading Frames in the Human Genome. Retrovirology 2004, 1, 32. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Elkahloun, A.G.; Candotti, F.; Grajkowski, A.; Beaucage, S.L.; Petricoin, E.F.; Calvert, V.; Juhl, H.; Mills, F.; Mason, K.; et al. A Novel Function of RNAs Arising from the Long Terminal Repeat of Human Endogenous Retrovirus 9 in Cell Cycle Arrest. J. Virol. 2013, 87, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Domansky, A.N.; Kopantzev, E.P.; Snezhkov, E.V.; Lebedev, Y.B.; Leib-Mosch, C.; Sverdlov, E.D. Solitary HERV-K LTRs Possess Bi-Directional Promoter Activity and Contain a Negative Regulatory Element in the U5 Region. FEBS Lett. 2000, 472, 191–195. [Google Scholar] [CrossRef] [Green Version]
- Manghera, M.; Magnusson, A.; Douville, R.N. The Sense behind Retroviral Anti-Sense Transcription. Virol. J. 2017, 14, 9. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.S.; Kaneko, S.; Girones, R.; Anderson, R.W.; Hornbuckle, W.E.; Tennant, B.C.; Cote, P.J.; Gerin, J.L.; Purcell, R.H.; Miller, R.H. The Woodchuck Hepatitis Virus X Gene Is Important for Establishment of Virus Infection in Woodchucks. J. Virol. 1993, 67, 1218–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimoda, A.; Sugata, F.; Chen, H.; Miller, R.H.; Purcell, R.H. Evidence for a Bidirectional Promoter Complex within the X Gene of Woodchuck Hepatitis Virus. Virus Res. 1998, 56, 25–39. [Google Scholar] [CrossRef]
- Velhagen, I.; Hilger, C.; Lamberts, C.; Zentgraf, H.; Schroder, C.H. An Antisense Promoter within the Hepatitis B Virus X Gene. Intervirology 1995, 38, 127–133. [Google Scholar] [CrossRef]
- Moriyama, K.; Hayashida, K.; Shimada, M.; Nakano, S.; Nakashima, Y.; Fukumaki, Y. Antisense RNAs Transcribed from the Upstream Region of the Precore/Core Promoter of Hepatitis B Virus. J. Gen. Virol. 2003, 84, 1907–1913. [Google Scholar] [CrossRef] [PubMed]
- Gunnery, S.; Mathews, M.B. Functional MRNA Can Be Generated by RNA Polymerase III. Mol. Cell. Biol. 1995, 15, 3597–3607. [Google Scholar] [CrossRef] [Green Version]
- Niwa, H.; Yamamura, K.; Miyazaki, J. Efficient Selection for High-Expression Transfectants with a Novel Eukaryotic Vector. Gene 1991, 108, 193–199. [Google Scholar] [CrossRef]
- Claverie, J.-M.; Abergel, C. Giant Viruses: The Difficult Breaking of Multiple Epistemological Barriers. Stud. Hist. Philos. Sci. Part. C Stud. Hist. Philos. Biol. Biomed. Sci. 2016, 59, 89–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gholizadeh, Z.; Iqbal, M.S.; Li, R.; Romerio, F. The HIV-1 Antisense Gene ASP: The New Kid on the Block. Vaccines 2021, 9, 513. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, T.S. The Structure of Scientific Revolutions, 1st ed.; University of Chicago Press: Chigaco, IL, USA, 1962. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miller, R.H.; Zimmer, A.; Moutot, G.; Mesnard, J.-M.; Chazal, N. Retroviral Antisense Transcripts and Genes: 33 Years after First Predicted, a Silent Retroviral Revolution? Viruses 2021, 13, 2221. https://0-doi-org.brum.beds.ac.uk/10.3390/v13112221
Miller RH, Zimmer A, Moutot G, Mesnard J-M, Chazal N. Retroviral Antisense Transcripts and Genes: 33 Years after First Predicted, a Silent Retroviral Revolution? Viruses. 2021; 13(11):2221. https://0-doi-org.brum.beds.ac.uk/10.3390/v13112221
Chicago/Turabian StyleMiller, Roger H., Alexis Zimmer, Gilles Moutot, Jean-Michel Mesnard, and Nathalie Chazal. 2021. "Retroviral Antisense Transcripts and Genes: 33 Years after First Predicted, a Silent Retroviral Revolution?" Viruses 13, no. 11: 2221. https://0-doi-org.brum.beds.ac.uk/10.3390/v13112221