
Extending the Coding Potential of Viral Genomes with Overlapping Antisense ORFs: A Case for the De Novo Creation of the Gene Encoding the Antisense Protein ASP of HIV-1


Abstract
:1. Introduction
2. Materials and Methods
2.1. Assembly of a Sample Set of 3725 env/asp Overlaps with ‘Intact’ asp and 1735 env/asp Overlaps with ‘Interrupted’ asp
2.2. Comparative Analysis of the Nucleotide Diversity in the Overlapping and Non-Overlapping Region of env Gene
2.3. Testing the Hypothesis of Asymmetric/Symmetric Evolution in the Antisense Overlap env/asp
2.4. Assembly of an Enlarged Sample Set of 4362 env/asp Overlaps
2.5. Detection of Critical Sites in the Antisense Protein by Comparative Analysis of the Amino Acid Composition
2.6. Assembly of a Dataset of env Sequences Covering a Wide Phylogenetic Range of Primate Lentiviruses
3. Results
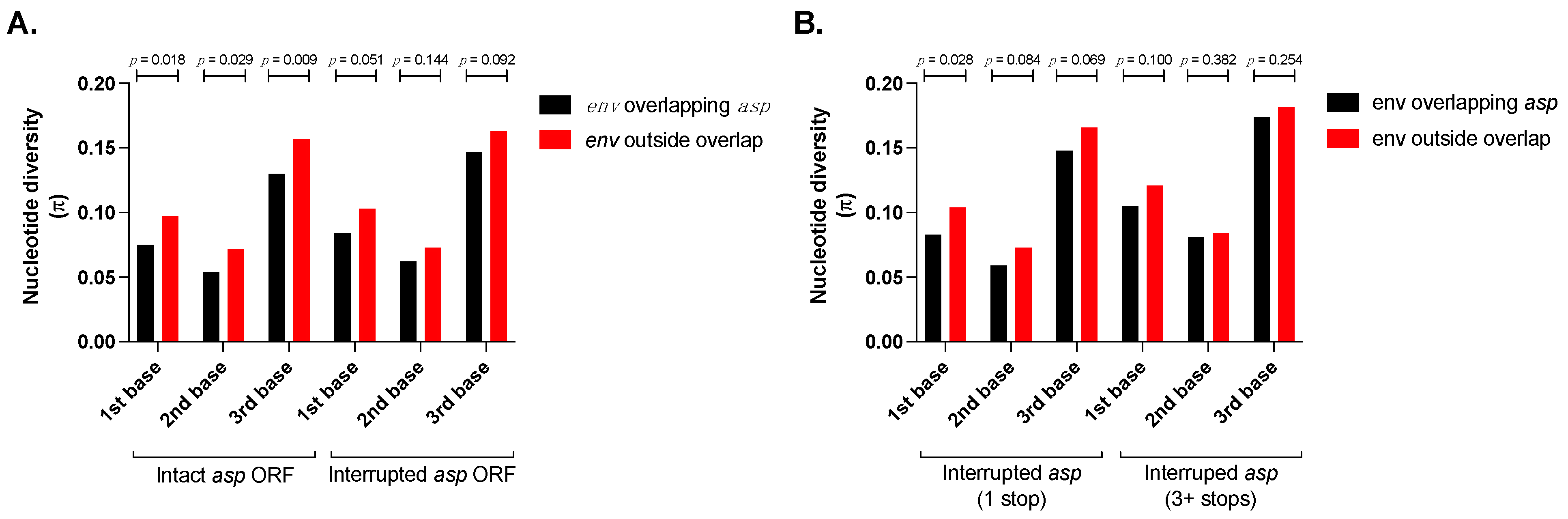
3.1. The Nucleotide Diversity of the env Region Overlapping asp Is Significantly Lower than That in the Non-Overlapping Region
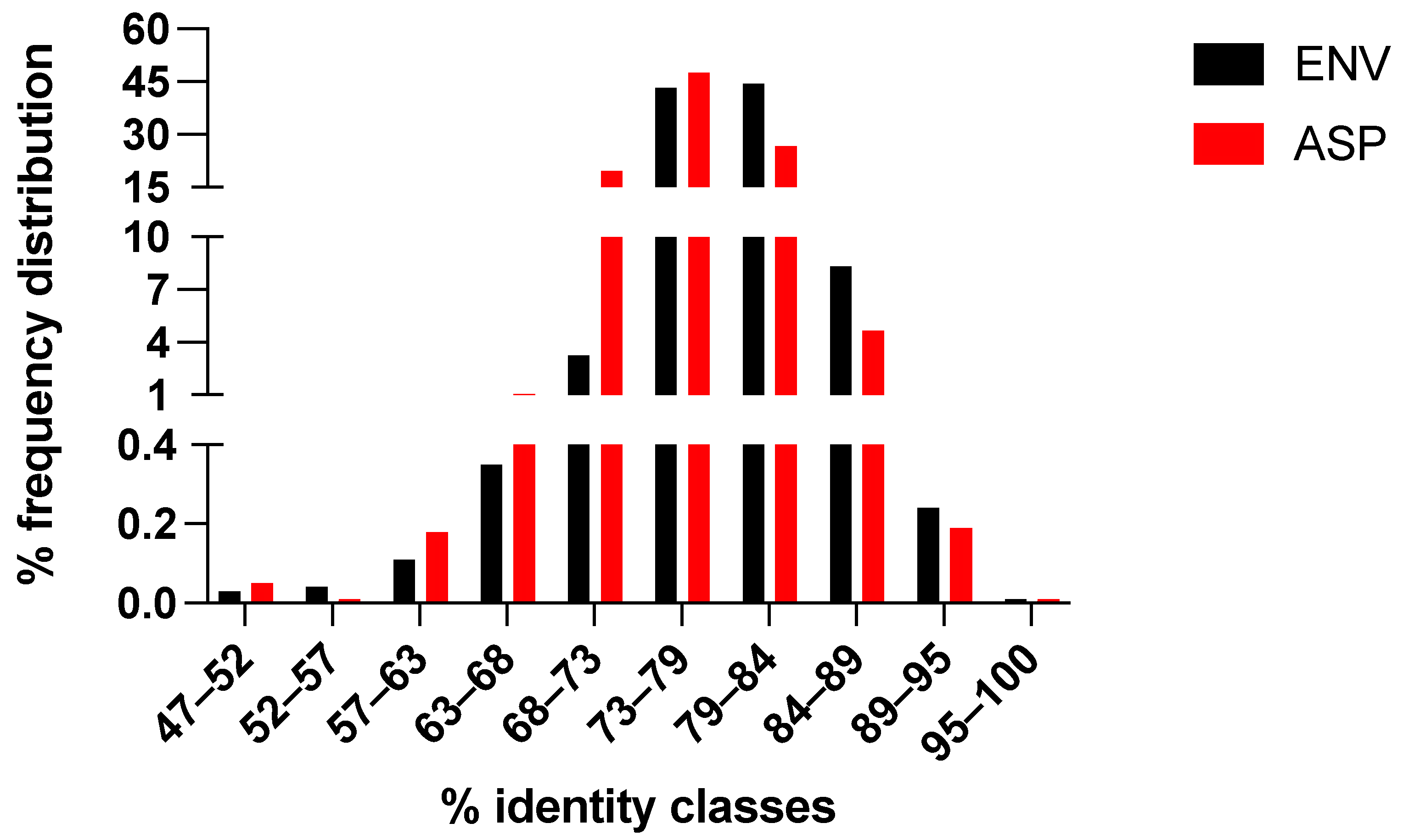
3.2. Symmetric Evolution of the env/asp Overlap
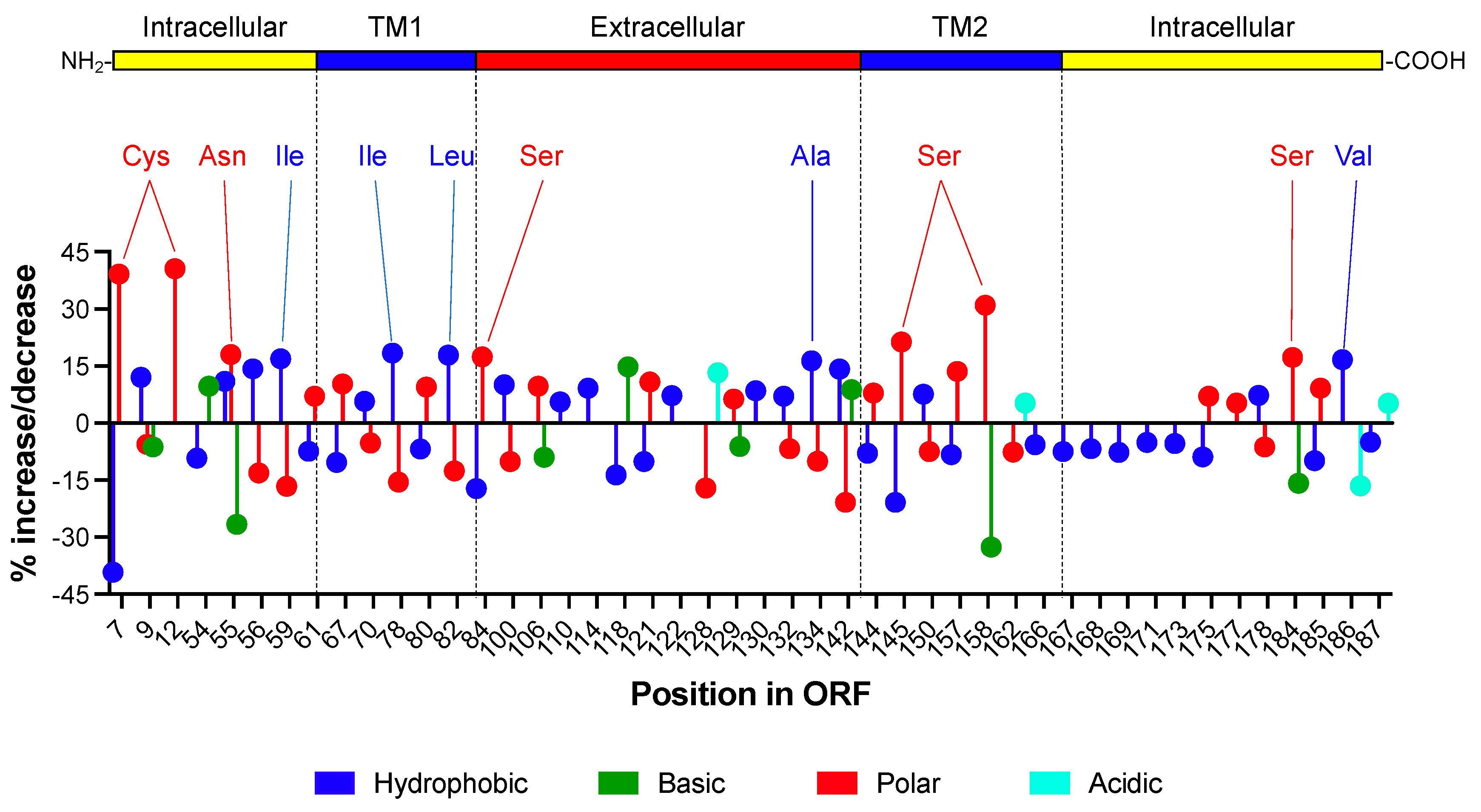
3.3. Identification of 46 Critical Amino Acid Sites in the Antisense Protein ASP
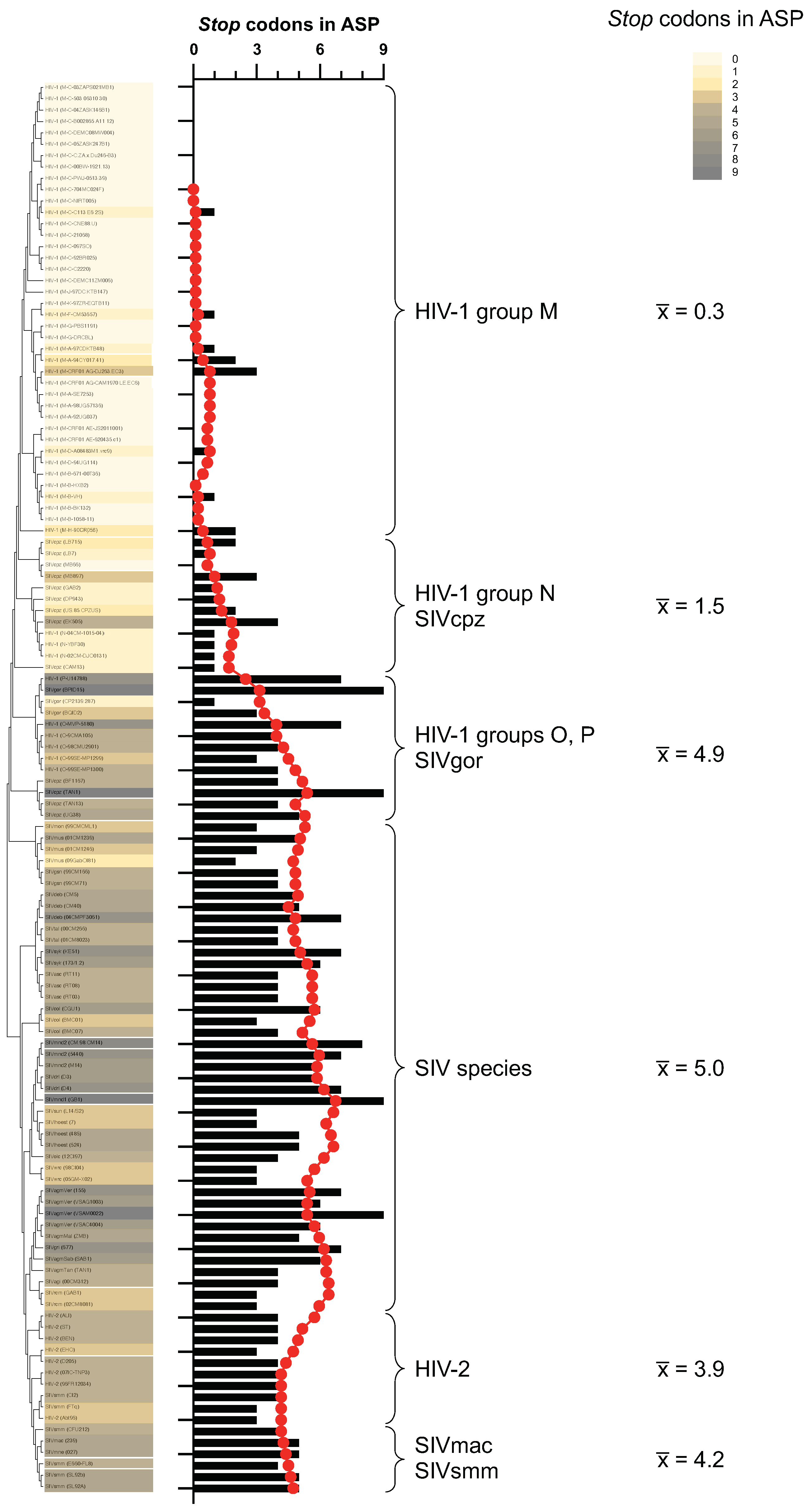
3.4. Analysis of env Sequences from a Large Dataset of Primate Lentiviruses Suggests a De Novo Origin of the Antisense ORF Encoding ASP
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Long, M.; Betran, E.; Thornton, K.; Wang, W. The origin of new genes: Glimpses from the young and old. Nat. Rev. Genet. 2003, 4, 865–875. [Google Scholar] [CrossRef]
- Jacob, F. Evolution and Tinkering. Science 1977, 196, 1161–1166. [Google Scholar] [CrossRef] [Green Version]
- Rancurel, C.; Khosravi, M.; Dunker, A.K.; Romero, P.R.; Karlin, D. Overlapping Genes Produce Proteins with Unusual Sequence Properties and Offer Insight into De Novo Protein Creation. J. Virol. 2009, 83, 10719–10736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabath, N.; Wagner, A.; Karlin, D. Evolution of viral proteins originated de novo by overprinting. Mol. Biol. Evol. 2012, 29, 3767–3780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, D.; Eisenberg, D. Finding families for genomic ORFans. Bioinformatics 1999, 15, 759–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, G.A.; Bertrand, N.; Patel, Y.; Hughes, J.B.; Feil, E.J.; Field, D. Orphans as taxonomically restricted and ecologically important genes. Microbiology 2005, 151, 2499–2501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belshaw, R.; Pybus, O.G.; Rambaut, A. The evolution of genome compression and genomic novelty in RNA viruses. Genome Res. 2007, 17, 1496–1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chirico, N.; Vianelli, A.; Belshaw, R. Why genes overlap in viruses. Proc. R. Soc. B Boil. Sci. 2010, 277, 3809–3817. [Google Scholar] [CrossRef] [Green Version]
- Pavesi, A. New insights into the evolutionary features of viral overlapping genes by discriminant analysis. Virology 2020, 546, 51–66. [Google Scholar] [CrossRef]
- Pavesi, A.; De Iaco, B.; Granero, M.I.; Porati, A. On the Informational Content of Overlapping Genes in Prokaryotic and Eukaryotic Viruses. J. Mol. Evol. 1997, 44, 625–631. [Google Scholar] [CrossRef]
- Pavesi, A.; Magiorkinis, G.; Karlin, D.G. Viral Proteins Originated De Novo by Overprinting Can Be Identified by Codon Usage: Application to the “Gene Nursery” of Deltaretroviruses. PLoS Comput. Biol. 2013, 9, e1003162. [Google Scholar] [CrossRef] [Green Version]
- Pavesi, A.; Vianelli, A.; Chirico, N.; Bao, Y.; Blinkova, O.; Belshaw, R.; Firth, A.; Karlin, D. Overlapping genes and the proteins they encode differ significantly in their sequence composition from non-overlapping genes. PLoS ONE 2018, 13, e0202513. [Google Scholar] [CrossRef] [Green Version]
- Pavesi, A. Asymmetric evolution in viral overlapping genes is a source of selective protein adaptation. Virology 2019, 532, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.W., Jr. Simultaneous codon usage, the origin of the proteome, and the emergence of de-novo proteins. Curr. Opin. Struct. Biol. 2021, 68, 142–148. [Google Scholar] [CrossRef]
- Karlin, D.; Ferron, F.; Canard, B.; Longhi, S. Structural disorder and modular organization in Paramyxovirinae N and P. J. Gen. Virol. 2003, 84, 3239–3252. [Google Scholar] [CrossRef]
- Willis, S.; Masel, J. Gene Birth Contributes to Structural Disorder Encoded by Overlapping Genes. Genetics 2018, 210, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Wensman, J.J.; Munir, M.; Thaduri, S.; Hörnaeus, K.; Rizwan, M.; Blomström, A.-L.; Briese, T.; Lipkin, W.I.; Berg, M. The X proteins of bornaviruses interfere with type I interferon signalling. J. Gen. Virol. 2013, 94, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Van Knippenberg, I.C.; Carlton-Smith, C.; Elliott, R.M. The N-terminus of Bunyamwera orthobunyavirus NSs protein is essential for interferon antagonism. J. Gen. Virol. 2010, 91, 2002–2006. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Ding, S.-W. Virus Counterdefense: Diverse Strategies for Evading the RNA-Silencing Immunity. Annu. Rev. Microbiol. 2006, 60, 503–531. [Google Scholar] [CrossRef] [Green Version]
- Chellappan, P.; Vanitharani, R.; Fauquet, C.M. MicroRNA-binding viral protein interferes with Arabidopsis development. Proc. Natl. Acad. Sci. USA 2005, 102, 10381–10386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Calvo, P.A.; Malide, D.; Gibbs, J.; Schubert, U.; Bacik, I.; Basta, S.; O’Neill, R.; Schickli, J.; Palese, P.; et al. A novel influenza A virus mitochondrial protein that induces cell death. Nat. Med. 2001, 7, 1306–1312. [Google Scholar] [CrossRef]
- Boehme, K.W.; Hammer, K.; Tollefson, W.C.; Konopka-Anstadt, J.L.; Kobayashi, T.; Dermody, T.S. Nonstructural protein sigma1s mediates reovirus-induced cell cycle arrest and apoptosis. J. Virol. 2013, 87, 12967–12979. [Google Scholar] [CrossRef] [Green Version]
- Taliansky, M.; Roberts, I.M.; Kalinina, N.; Ryabov, E.; Raj, S.K.; Robinson, D.J.; Oparka, K.J. An Umbraviral Protein, Involved in Long-Distance RNA Movement, Binds Viral RNA and Forms Unique, Protective Ribonucleoprotein Complexes. J. Virol. 2003, 77, 3031–3040. [Google Scholar] [CrossRef] [Green Version]
- Singh, M. No vaccine against HIV yet-Are we not perfectly equipped? Virol. J. 2006, 3, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pluta, A.; Jaworski, J.P.; Douville, R.N. Regulation of Expression and Latency in BLV and HTLV. Viruses 2020, 12, 1079. [Google Scholar] [CrossRef]
- Matsuoka, M.; Mesnard, J.-M. HTLV-1 bZIP factor: The key viral gene for pathogenesis. Retrovirology 2020, 17, 2. [Google Scholar] [CrossRef]
- Satou, Y.; Yasunaga, J.; Yoshida, M.; Matsuoka, M. HTLV-I basic leucine zipper factor gene mRNA supports proliferation of adult T cell leukemia cells. Proc. Natl. Acad. Sci. USA 2006, 103, 720–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, M.; Satou, Y.; Yasunaga, J.-I.; Fujisawa, J.-I.; Matsuoka, M. Transcriptional Control of Spliced and Unspliced Human T-Cell Leukemia Virus Type 1 bZIP Factor (HBZ) Gene. J. Virol. 2008, 82, 9359–9368. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, M.; Yasunaga, J.-I.; Taniguchi, Y.; Tamiya, S.; Nakahata, T.; Matsuoka, M. Preferential Selection of Human T-Cell Leukemia Virus Type 1 Provirus Lacking the 5′ Long Terminal Repeat during Oncogenesis. J. Virol. 2007, 81, 5714–5723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taniguchi, Y.; Nosaka, K.; Yasunaga, J.-I.; Maeda, M.; Mueller, N.; Okayama, A.; Matsuoka, M. Silencing of human T-cell leukemia virus type I gene transcription by epigenetic mechanisms. Retrovirology 2005, 2, 64. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.; Ma, G.; Nosaka, K.; Tanabe, J.; Satou, Y.; Koito, A.; Wain-Hobson, S.; Vartanian, J.-P.; Matsuoka, M. APOBEC3G Generates Nonsense Mutations in Human T-Cell Leukemia Virus Type 1 Proviral Genomes In Vivo. J. Virol. 2010, 84, 7278. [Google Scholar] [CrossRef] [Green Version]
- Savoret, J.; Mesnard, J.-M.; Gross, A.; Chazal, N. Antisense Transcripts and Antisense Protein: A New Perspective on Human Immunodeficiency Virus Type 1. Front. Microbiol. 2021, 11, 625941. [Google Scholar] [CrossRef]
- Miura, M.; Yasunaga, J.-I.; Tanabe, J.; Sugata, K.; Zhao, T.; Ma, G.; Miyazato, P.; Ohshima, K.; Kaneko, A.; Watanabe, A.; et al. Characterization of simian T-cell leukemia virus type 1 in naturally infected Japanese macaques as a model of HTLV-1 infection. Retrovirology 2013, 10, 118. [Google Scholar] [CrossRef] [Green Version]
- Miller, R.H. Human Immunodeficiency Virus May Encode a Novel Protein on the Genomic DNA Plus Strand. Science 1988, 239, 1420–1422. [Google Scholar] [CrossRef] [PubMed]
- Affram, Y.; Zapata, J.C.; Gholizadeh, Z.; Tolbert, W.D.; Zhou, W.; Iglesias-Ussel, M.D.; Pazgier, M.; Ray, K.; Latinovic, O.S.; Romerio, F. The HIV-1 Antisense Protein ASP Is a Transmembrane Protein of the Cell Surface and an Integral Protein of the Viral Envelope. J. Virol. 2019, 93, e00574-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clerc, I.; Laverdure, S.; Torresilla, C.; Landry, S.; Borel, S.; Vargas, A.; Arpin-André, C.; Gay, B.; Briant, L.; Gross, A.; et al. Polarized expression of the membrane ASP protein derived from HIV-1 antisense transcription in T cells. Retrovirology 2011, 8, 74. [Google Scholar] [CrossRef] [Green Version]
- Gholizadeh, Z.; Iqbal, M.; Li, R.; Romerio, F. The HIV-1 Antisense Gene ASP: The New Kid on the Block. Vaccines 2021, 9, 513. [Google Scholar] [CrossRef]
- Cassan, E.; Arigon-Chifolleau, A.-M.; Mesnard, J.-M.; Gross, A.; Gascuel, O. Concomitant emergence of the antisense protein gene of HIV-1 and of the pandemic. Proc. Natl. Acad. Sci. USA 2016, 113, 11537–11542. [Google Scholar] [CrossRef] [Green Version]
- Dimonte, S. Different HIV-1 env frames: gp120 and ASP (antisense protein) biosynthesis, and their co-variation tropic amino acid signatures in X4- and R5-viruses. J. Med. Virol. 2017, 89, 112–122. [Google Scholar] [CrossRef]
- Nelson, C.W.; Ardern, Z.; Wei, X. OLGenie: Estimating Natural Selection to Predict Functional Overlapping Genes. Mol. Biol. Evol. 2020, 37, 2440–2449. [Google Scholar] [CrossRef]
- Nei, M.; Li, W.-H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef] [Green Version]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods; Iowa State University Press: Ames, IA, USA, 1967; p. 593. [Google Scholar]
- Olsen, H.S.; Nelbock, P.; Cochrane, A.W.; Rosen, C.A. Secondary Structure is the Major Determinant for Interaction of HIV rev Protein with RNA. Science 1990, 247, 845–848. [Google Scholar] [CrossRef]
- Hall, B.G. Building Phylogenetic Trees from Molecular Data with MEGA. Mol. Biol. Evol. 2013, 30, 1229–1235. [Google Scholar] [CrossRef] [Green Version]
- Keele, B.F.; Van Heuverswyn, F.; Li, Y.; Bailes, E.; Takehisa, J.; Santiago, M.L.; Bibollet-Ruche, F.; Chen, Y.; Wain, L.V.; Liegeois, F.; et al. Chimpanzee Reservoirs of Pandemic and Nonpandemic HIV-1. Science 2006, 313, 523–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Chen, J.; Deng, L.; Mao, Q.; Zheng, J.; Wu, J.; Zeng, C.; Li, Y. Evolutionary selection associated with the multi-function of overlapping genes in the hepatitis B virus. Infect. Genet. Evol. 2010, 10, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Torresilla, C.; Xiao, Y.; Nguyen, P.T.; Caté, C.; Barbosa, K.; Rassart, É.; Cen, S.; Bourgault, S.; Barbeau, B. HIV-1 Antisense Protein of Different Clades Induces Autophagy and Associates with the Autophagy Factor p62. J. Virol. 2019, 93, e01757-18. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus Species (Group-Clade-Isolate) | NCBI acc. # | Codons from ATG to First Stop | Stops between Canonical ATG and Stop | Mean Number (sd) of Internal Stops in the 10,000× Permutated Antisense ORF | Percent of 10,000× Permutated Antisense ORF Not Interrupted by Stops |
|---|---|---|---|---|---|
| SIVgor (BPID15) | KP004990 | 67 | 9 | 7.5 (1.6) | 0 |
| SIVmnd1 (GB1) | M27470 | 29 | 9 | 7.3 (1.2) | 0 |
| SIVagmVer (VSAM0022) | KR862356 | 19 | 9 | 7.0 (1.7) | 0 |
| SIVcpz (TAN1) | AF447763 | 17 | 9 | 6.2 (1.5) | 0 |
| SIVagmVer (155) | M29975 | 18 | 7 | 7.0 (1.5) | 0 |
| SIVgri (677) | M58410 | 85 | 7 | 6.0 (1.3) | 0 |
| HIV-1 (P-U14788) | HQ179987 | 70 | 7 | 6.0 (1.1) | 0 |
| SIVagmVer (VSAC4004) | KR862336 | 97 | 6 | 6.3 (1.4) | 0 |
| SIVagmVer (VSAG1003) | KR862363 | 18 | 6 | 5.7 (1.5) | 0 |
| SIVmac (239) | M33262 | 34 | 5 | 5.0 (1.0) | 0 |
| SIVagmMal (ZMB) | LC114462 | 29 | 5 | 5.0 (1.3) | 0 |
| SIVmne (027) | U79412 | 34 | 5 | 4.9 (1.1) | 0 |
| SIVcpz (UG38) | JN91690 | 54 | 5 | 3.9 (1.3) | 0 |
| SIVgsn (99CM71) | AF468658 | 75 | 4 | 2.9 (1.3) | 2 |
| SIVsmm (CI2) | JX860430 | 16 | 4 | 2.9 (0.8) | 0 |
| SIVsmm (CFU212) | JX860407 | 50 | 4 | 4.4 (1.0) | 0 |
| SIVagmTan (TAN1) | U58991 | 90 | 4 | 4.4 (1.3) | 0 |
| SIVcpz (BF1167) | JQ866001 | 19 | 4 | 3.0 (1.0) | 0 |
| SIVcpz (EK505) | DQ373065 | 117 | 4 | 4.0 (1.3) | 0 |
| SIVtal (00CM266) | AY655744 | 116 | 4 | 5.0 (1.3) | 0 |
| SIVtal (01CM8023) | AM182197 | 72 | 4 | 5.3 (1.2) | 0 |
| SIVgsn (99CM166) | AF468659 | 94 | 4 | 3.8 (1.2) | 0 |
| SIVsmm (FTq) | JX860414 | 42 | 4 | 4.7 (1.2) | 0 |
| HIV-2 (96FR12034) | AY530889 | 38 | 4 | 3.9 (1.0) | 0 |
| HIV-1 (M-CRF01-AG-91DJ_263) | JQ715398 | 67 | 3 | 2.1 (1.0) | 4 |
| SIVmus (01CM1246) | EF070329 | 125 | 3 | 3.3 (1.5) | 2 |
| SIVgor (CP2139.287) | FJ424866 | 85 | 3 | 2.9 (1.2) | 1 |
| HIV-2 (EHO) | U27200 | 42 | 3 | 3.5 (0.9) | 0 |
| SIVcpz (MB897) | EF535994 | 127 | 3 | 2.9 (1.0) | 0 |
| SIVgor (BQID2) | KP004991 | 176 | 3 | 4.1 (1.3) | 0 |
| SIVcpz (GAB2) | AF382828 | 96 | 2 | 1.2 (0.9) | 24 |
| SIVcpz (US.85.CPZUS) | AF103818 | 151 | 2 | 1.6 (0.9) | 10 |
| HIV-1 (M-A-94CY017.41) | AF286237 | 44 | 2 | 1.5 (0.9) | 9 |
| SIVmus (09GabOI81) | KF304707 | 0 | 2 | 3.2 (1.2) | 0 |
| SIVcpz (LB715) | KP861923 | 148 | 2 | 3.1 (1.0) | 0 |
| HIV-1 (M-H-90CR056) | AF005496 | 108 | 2 | 3.1 (0.8) | 0 |
| SIVcpz (DP943) | EF535993 | 145 | 1 | 0.7 (0.7) | 41 |
| HIV-1 (M-D-A08483M1.vrc9) | HM215358 | 110 | 1 | 0.8 (0.7) | 39 |
| HIV-1 (M-F-CM53657) | AF377956 | 132 | 1 | 1.3 (0.9) | 21 |
| HIV-1 (N-02CM-DJO0131) | AY532635 | 136 | 1 | 1.2 (0.8) | 20 |
| HIV-1 (M-C-C113-E6-2S) | HM639260 | 136 | 1 | 1.5 (1.1) | 17 |
| HIV-1 (M-B-VH) | AF146728 | 113 | 1 | 1.4 (0.9) | 13 |
| HIV-1 (N-YBF30) | AJ006022 | 139 | 1 | 1.9 (1.1) | 8 |
| SIVcpz (LB7) | DQ373064 | 88 | 1 | 1.4 (0.6) | 0 |
| HIV-1 (M-A-97CDKTB48) | AF286238 | 140 | 1 † | 1.6 (0.7) | 0 |
| HIV-1 (M-B-1058-11) | AY331295 | 191 | 0 | 0.4 (0.6) | 66 |
| HIV-1 (M-C-503-06310-30) | KT183169 | 171 | 0 | 0.4 (0.6) | 64 |
| HIV-1 (M-C-03ZAPS021MB1) | DQ369978 | 182 | 0 | 0.4 (0.6) | 65 |
| HIV-1 (M-C-04ZASK146B1) | AY772699 | 187 | 0 | 0.4 (0.6) | 62 |
| HIV-1 (M-C-92BR025) | U52953 | 187 | 0 | 0.5 (0.6) | 60 |
| HIV-1 (M-CRF01-AE-620435_c01) | JX512900 | 189 | 0 | 0.5 (0.7) | 55 |
| HIV-1 (M-C-21068) | AF067155 | 193 | 0 | 0.6 (0.7) | 53 |
| HIV-1 (M-C-097SO) | MF373199 | 184 | 0 | 0.6 (0.7) | 50 |
| HIV-1 (M-G-DRCBL) | AF084936 | 183 | 0 | 0.6 (0.7) | 50 |
| SIVcpz (MB66) | DQ373063 | 182 | 0 | 0.6 (0.7) | 49 |
| HIV-1 (M-C-C2220) | U46016 | 183 | 0 | 0.7 (0.7) | 47 |
| HIV-1 (M-B-HXB2) | K03455 | 190 | 0 | 0.8 (0.8) | 41 |
| HIV-1 (M-J-97DC.KTB147) | EF614151 | 186 | 0 | 0.9 (0.8) | 38 |
| HIV-1 (M-B-671-00T36) | AY423387 | 188 | 0 | 0.9 (0.8) | 36 |
| HIV-1 (M-C-05ZASK247B1) | DQ369994 | 181 | 0 | 0.9 (0.8) | 34 |
| HIV-1 (M-G-PBS1191) | MH705134 | 181 | 0 | 1.0 (0.9) | 30 |
| HIV-1 (M-D-94UG114) | U88824 | 188 | 0 | 1.0 (0.8) | 30 |
| HIV-1 (M-K-97ZR-EQTB11) | AJ249235 | 180 | 0 | 1.2 (0.9) | 25 |
| HIV-1 (M-CRF01-AG-CAM1970 LE.EC6) | JQ715394 | 155 | 0 †,‡ | 0.5 (0.7) | 59 |
| HIV-1 (M-A-92UG037) | U51190 | 158 | 0 † | 0.6 (0.7) | 53 |
| HIV-1 (M-CRF01-AE-JS2011001) | KM111555 | 176 | 0 ‡ | 0.6 (0.7) | 49 |
| HIV-1 (M-A-98UG57136) | AF484509 | 162 | 0 † | 1.1 (0.9) | 26 |
| HIV-1 (M-A-SE7253) | AF069670 | 162 | 0 † | 1.2 (0.9) | 22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavesi, A.; Romerio, F. Extending the Coding Potential of Viral Genomes with Overlapping Antisense ORFs: A Case for the De Novo Creation of the Gene Encoding the Antisense Protein ASP of HIV-1. Viruses 2022, 14, 146. https://0-doi-org.brum.beds.ac.uk/10.3390/v14010146
Pavesi A, Romerio F. Extending the Coding Potential of Viral Genomes with Overlapping Antisense ORFs: A Case for the De Novo Creation of the Gene Encoding the Antisense Protein ASP of HIV-1. Viruses. 2022; 14(1):146. https://0-doi-org.brum.beds.ac.uk/10.3390/v14010146
Chicago/Turabian StylePavesi, Angelo, and Fabio Romerio. 2022. "Extending the Coding Potential of Viral Genomes with Overlapping Antisense ORFs: A Case for the De Novo Creation of the Gene Encoding the Antisense Protein ASP of HIV-1" Viruses 14, no. 1: 146. https://0-doi-org.brum.beds.ac.uk/10.3390/v14010146