
Epidemiology and Genetic Variability of HHV-8/KSHV among Rural Populations and Kaposi’s Sarcoma Patients in Gabon, Central Africa. Review of the Geographical Distribution of HHV-8 K1 Genotypes in Africa


Abstract
:1. Introduction
2. Materials and Methods
2.1. Rural Population Survey
2.2. Ethical Statement
2.3. HHV-8 Serological Tests
2.4. Molecular and Phylogenetic Analyses of HHV-8
2.5. Nucleotide Sequence Accession Numbers
2.6. Statistical Analyses
2.7. HHV-8 Genotype Distribution: Search Strategy and Data Extraction
3. Results
3.1. Rural Populations
3.2. Sero-Epidemiology of HHV-8 in Rural Populations
3.3. HHV-8 Molecular Results
3.4. HHV-8 Genetic Variability
3.5. HHV-8 Phylogenetic Analyses
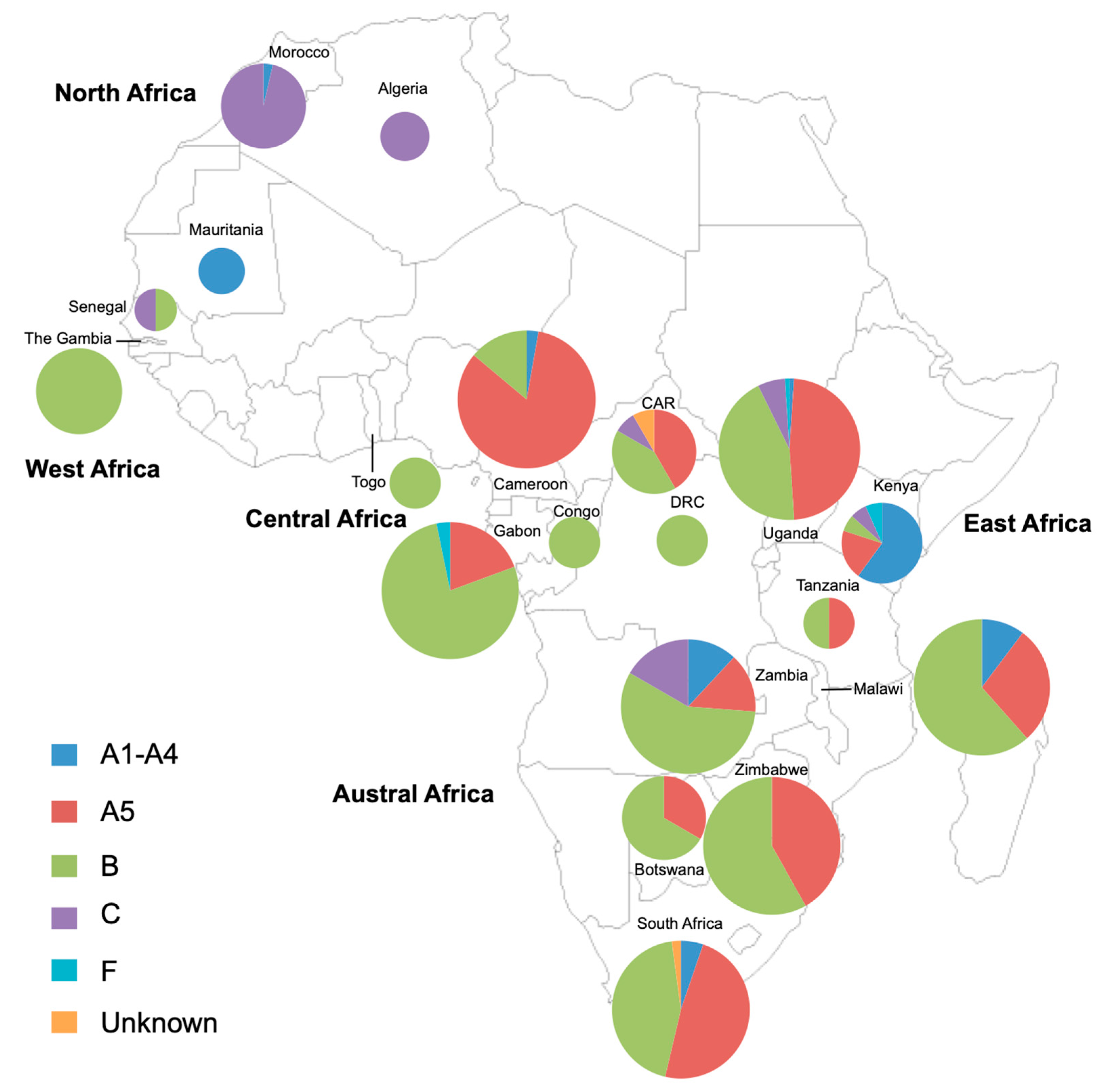
3.6. Review of the Diversity of HHV-8 Strains in Africa Based on the K1 Gene Study
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chang, Y.; Cesarman, E.; Pessin, M.S.; Lee, F.; Culpepper, J.; Knowles, D.M.; Moore, P.S. Identification of herpesvirus-like DNA sequences in AIDS-associated Kaposi’s sarcoma. Science 1994, 266, 1865–1869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boshoff, C.; Schulz, T.F.; Kennedy, M.M.; Graham, A.K.; Fisher, C.; Thomas, A.; McGee, J.O.; Weiss, R.A.; O’Leary, J.J. Kaposi’s sarcoma-associated herpesvirus infects endothelial and spindle cells. Nat. Med. 1995, 1, 1274–1278. [Google Scholar] [CrossRef] [PubMed]
- Uldrick, T.S.; Whitby, D. Update on KSHV epidemiology, Kaposi Sarcoma pathogenesis, and treatment of Kaposi Sarcoma. Cancer Lett. 2011, 305, 150–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitby, D.; Howard, M.R.; Tenant-Flowers, M.; Brink, N.S.; Copas, A.; Boshoff, C.; Hatzioannou, T.; Suggett, F.E.; Aldam, D.M.; Denton, A.S.; et al. Detection of Kaposi sarcoma associated herpesvirus in peripheral blood of HIV-infected individuals and progression to Kaposi’s sarcoma. Lancet 1995, 346, 799–802. [Google Scholar] [CrossRef]
- Nador, R.G.; Cesarman, E.; Chadburn, A.; Dawson, D.B.; Ansari, M.Q.; Sald, J.; Knowles, D.M. Primary effusion lymphoma: A distinct clinicopathologic entity associated with the Kaposi’s sarcoma-associated herpes virus. Blood 1996, 88, 645–656. [Google Scholar] [CrossRef] [Green Version]
- Gessain, A.; Sudaka, A.; Briere, J.; Fouchard, N.; Nicola, M.A.; Rio, B.; Arborio, M.; Troussard, X.; Audouin, J.; Diebold, J.; et al. Kaposi sarcoma-associated herpes-like virus (human herpesvirus type 8) DNA sequences in multicentric Castleman’s disease: Is there any relevant association in non-human immunodeficiency virus-infected patients? Blood 1996, 87, 414–416. [Google Scholar] [CrossRef]
- Soulier, J.; Grollet, L.; Oksenhendler, E.; Cacoub, P.; Cazals-Hatem, D.; Babinet, P.; d’Agay, M.F.; Clauvel, J.P.; Raphael, M.; Degos, L.; et al. Kaposi’s sarcoma-associated herpesvirus-like DNA sequences in multicentric Castleman’s disease. Blood 1995, 86, 1276–1280. [Google Scholar] [CrossRef] [Green Version]
- Chadburn, A.; Hyjek, E.; Mathew, S.; Cesarman, E.; Said, J.; Knowles, D.M. KSHV-positive solid lymphomas represent an extra-cavitary variant of primary effusion lymphoma. Am. J. Surg. Pathol. 2004, 28, 1401–1416. [Google Scholar] [CrossRef]
- Beral, V. Epidemiology of Kaposi’s sarcoma. Cancer Surv. 1991, 10, 5–22. [Google Scholar]
- Boshoff, C.; Weiss, R.A. Epidemiology and pathogenesis of Kaposi’s sarcoma-associated herpesvirus. Philos. Trans. R Soc. Lond. B Biol. Sci. 2001, 356, 517–534. [Google Scholar] [CrossRef] [Green Version]
- De-The, G.; Bestetti, G.; van Beveren, M.; Gessain, A. Prevalence of human herpesvirus 8 infection before the acquired immunodeficiency disease syndrome-related epidemic of Kaposi’s sarcoma in East Africa. J. Natl. Cancer Inst. 1999, 91, 1888–1889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dukers, N.H.; Rezza, G. Human herpesvirus 8 epidemiology: What we do and do not know. Aids 2003, 17, 1717–1730. [Google Scholar] [CrossRef] [PubMed]
- Gessain, A.; Mauclere, P.; van Beveren, M.; Plancoulaine, S.; Ayouba, A.; Essame-Oyono, J.L.; Martin, P.M.; de The, G. Human herpesvirus 8 primary infection occurs during childhood in Cameroon, Central Africa. Int. J. Cancer 1999, 81, 189–192. [Google Scholar] [CrossRef]
- Hengge, U.R.; Ruzicka, T.; Tyring, S.K.; Stuschke, M.; Roggendorf, M.; Schwartz, R.A.; Seeber, S. Update on Kaposi’s sarcoma and other HHV8 associated diseases. Part 2: Pathogenesis, Castleman’s disease, and pleural effusion lymphoma. Lancet Infect. Dis. 2002, 2, 344–352. [Google Scholar] [CrossRef]
- Zhang, T.; Shao, X.; Chen, Y.; Zhang, T.; Minhas, V.; Wood, C.; He, N. Human herpesvirus 8 seroprevalence, China. Emerg. Infect. Dis. 2012, 18, 150–152. [Google Scholar] [CrossRef] [PubMed]
- Biggar, R.J.; Whitby, D.; Marshall, V.; Linhares, A.C.; Black, F. Human herpesvirus 8 in Brazilian Amerindians: A hyperendemic population with a new subtype. J. Infect. Dis. 2000, 181, 1562–1568. [Google Scholar] [CrossRef] [Green Version]
- De Souza, V.A.; Sumita, L.M.; Nascimento, M.C.; Oliveira, J.; Mascheretti, M.; Quiroga, M.; Freire, W.S.; Tateno, A.; Boulos, M.; Mayaud, P.; et al. Human herpesvirus-8 infection and oral shedding in Amerindian and non-Amerindian populations in the Brazilian Amazon region. J. Infect. Dis. 2007, 196, 844–852. [Google Scholar] [CrossRef] [Green Version]
- Kazanji, M.; Dussart, P.; Duprez, R.; Tortevoye, P.; Pouliquen, J.F.; Vandekerkhove, J.; Couppie, P.; Morvan, J.; Talarmin, A.; Gessain, A. Serological and Molecular Evidence That Human Herpesvirus 8 Is Endemic among Amerindians in French Guiana. J. Infect. Dis. 2005, 192, 1525–1529. [Google Scholar] [CrossRef] [Green Version]
- Whitby, D.; Marshall, V.A.; Bagni, R.K.; Wang, C.D.; Gamache, C.J.; Guzman, J.R.; Kron, M.; Ebbesen, P.; Biggar, R.J. Genotypic characterization of Kaposi’s sarcoma-associated herpesvirus in asymptomatic infected subjects from isolated populations. J. Gen. Virol. 2004, 85 (Pt 1), 155–163. [Google Scholar] [CrossRef]
- Casper, C.; Wald, A.; Pauk, J.; Tabet, S.R.; Corey, L.; Celum, C.L. Correlates of prevalent and incident Kaposi’s sarcoma-associated herpesvirus infection in men who have sex with men. J. Infect. Dis. 2002, 185, 990–993. [Google Scholar] [CrossRef] [Green Version]
- Dukers, N.H.; Renwick, N.; Prins, M.; Geskus, R.B.; Schulz, T.F.; Weverling, G.J.; Coutinho, R.A.; Goudsmit, J. Risk factors for human herpesvirus 8 seropositivity and seroconversion in a cohort of homosexual men. Am. J. Epidemiol. 2000, 151, 213–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, J.N.; Ganem, D.E.; Osmond, D.H.; Page-Shafer, K.A.; Macrae, D.; Kedes, D.H. Sexual transmission and the natural history of human herpesvirus 8 infection. N. Engl. J. Med. 1998, 338, 948–954. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, T.R.; Kedes, D.; Ganem, D.; Macrae, D.R.; Rosenberg, P.S.; Molden, J.; Goedert, J.J. Evidence for concurrent epidemics of human herpesvirus 8 and human immunodeficiency virus type 1 in US homosexual men: Rates, risk factors, and relationship to Kaposi’s sarcoma. J. Infect. Dis. 1999, 180, 1010–1017. [Google Scholar] [CrossRef] [Green Version]
- Casper, C.; Redman, M.; Huang, M.L.; Pauk, J.; Lampinen, T.M.; Hawes, S.E.; Critchlow, C.W.; Morrow, R.A.; Corey, L.; Kiviat, N.; et al. HIV infection and human herpesvirus-8 oral shedding among men who have sex with men. J. Acquir. Immune. Defic. Syndr. 2004, 35, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Plancoulaine, S.; Abel, L.; van Beveren, M.; Tregouet, D.A.; Joubert, M.; Tortevoye, P.; de The, G.; Gessain, A. Human herpesvirus 8 transmission from mother to child and between siblings in an endemic population. Lancet 2000, 356, 1062–1065. [Google Scholar] [CrossRef]
- Butler, L.M.; Dorsey, G.; Hladik, W.; Rosenthal, P.J.; Brander, C.; Neilands, T.B.; Mbisa, G.; Whitby, D.; Kiepiela, P.; Mosam, A.; et al. Kaposi sarcoma-associated herpesvirus (KSHV) seroprevalence in population-based samples of African children: Evidence for at least 2 patterns of KSHV transmission. J. Infect. Dis. 2009, 200, 430–438. [Google Scholar] [CrossRef]
- Cook, P.M.; Whitby, D.; Calabro, M.L.; Luppi, M.; Kakoola, D.N.; Hjalgrim, H.; Ariyoshi, K.; Ensoli, B.; Davison, A.J.; Schulz, T.F. Variability and evolution of Kaposi’s sarcoma-associated herpesvirus in Europe and Africa. International Collaborative Group. Aids 1999, 13, 1165–1176. [Google Scholar] [CrossRef] [PubMed]
- Zong, J.C.; Ciufo, D.M.; Alcendor, D.J.; Wan, X.; Nicholas, J.; Browning, P.J.; Rady, P.L.; Tyring, S.K.; Orenstein, J.M.; Rabkin, C.S.; et al. High-level variability in the ORF-K1 membrane protein gene at the left end of the Kaposi’s sarcoma-associated herpesvirus genome defines four major virus subtypes and multiple variants or clades in different human populations. J. Virol. 1999, 73, 4156–4170. [Google Scholar] [CrossRef] [Green Version]
- Betsem, E.; Cassar, O.; Afonso, P.V.; Fontanet, A.; Froment, A.; Gessain, A. Epidemiology and genetic variability of HHV-8/KSHV in Pygmy and Bantu populations in Cameroon. PLoS Negl. Trop Dis. 2014, 8, e2851. [Google Scholar] [CrossRef] [Green Version]
- Cassar, O.; Afonso, P.V.; Bassot, S.; Plancoulaine, S.; Duprez, R.; Capuano, C.; Abel, M.; Martin, P.M.; Gessain, A. Novel human herpesvirus 8 subtype D strains in Vanuatu, Melanesia. Emerg. Infect. Dis. 2007, 13, 1745–1748. [Google Scholar] [CrossRef]
- Cassar, O.; Bassot, S.; Plancoulaine, S.; Quintana-Murci, L.; Harmant, C.; Gurtsevitch, V.; Senyuta, N.B.; Yakovleva, L.S.; de The, G.; Gessain, A. Human herpesvirus 8, Southern Siberia. Emerg. Infect. Dis. 2010, 16, 580–582. [Google Scholar] [CrossRef] [PubMed]
- Duprez, R.; Cassar, O.; Hbid, O.; Rougier, Y.; Morisse, L.; Bassot, S.; Huerre, M.; Gessain, A. Cutaneous disseminated endemic Kaposi’s sarcoma in a Polynesian man infected with a new divergent Human Herpesvirus 8 subtype D. J. Clin. Virol. 2006, 37, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Cassar, O.; Blondot, M.L.; Mohanna, S.; Jouvion, G.; Bravo, F.; Maco, V.; Duprez, R.; Huerre, M.; Gotuzzo, E.; Gessain, A. Human herpesvirus 8 genotype E in patients with Kaposi sarcoma, Peru. Emerg. Infect. Dis. 2010, 16, 1459–1462. [Google Scholar] [CrossRef] [PubMed]
- Isaacs, T.; Abera, A.B.; Muloiwa, R.; Katz, A.A.; Todd, G. Genetic diversity of HHV8 subtypes in South Africa: A5 subtype is associated with extensive disease in AIDS-KS. J. Med. Virol. 2016, 88, 292–303. [Google Scholar] [CrossRef]
- Kajumbula, H.; Wallace, R.G.; Zong, J.C.; Hokello, J.; Sussman, N.; Simms, S.; Rockwell, R.F.; Pozos, R.; Hayward, G.S.; Boto, W. Ugandan Kaposi’s sarcoma-associated herpesvirus phylogeny: Evidence for cross-ethnic transmission of viral subtypes. Intervirology 2006, 49, 133–143. [Google Scholar] [CrossRef]
- Kakoola, D.N.; Sheldon, J.; Byabazaire, N.; Bowden, R.J.; Katongole-Mbidde, E.; Schulz, T.F.; Davison, A.J. Recombination in human herpesvirus-8 strains from Uganda and evolution of the K15 gene. J. Gen. Virol. 2001, 82 (Pt 10), 2393–2404. [Google Scholar] [CrossRef]
- Kasolo, F.C.; Monze, M.; Obel, N.; Anderson, R.A.; French, C.; Gompels, U.A. Sequence analyses of human herpesvirus-8 strains from both African human immunodeficiency virus-negative and -positive childhood endemic Kaposi’s sarcoma show a close relationship with strains identified in febrile children and high variation in the K1 glycoprotein. J. Gen. Virol. 1998, 79 (Pt 12), 3055–3065. [Google Scholar]
- Lacoste, V.; Judde, J.G.; Briere, J.; Tulliez, M.; Garin, B.; Kassa-Kelembho, E.; Morvan, J.; Couppie, P.; Clyti, E.; Forteza Vila, J.; et al. Molecular epidemiology of human herpesvirus 8 in africa: Both B and A5 K1 genotypes, as well as the M and P genotypes of K14.1/K15 loci, are frequent and widespread. Virology 2000, 278, 60–74. [Google Scholar] [CrossRef] [Green Version]
- Meng, Y.X.; Sata, T.; Stamey, F.R.; Voevodin, A.; Katano, H.; Koizumi, H.; Deleon, M.; De Cristofano, M.A.; Galimberti, R.; Pellett, P.E. Molecular characterization of strains of Human herpesvirus 8 from Japan, Argentina and Kuwait. J. Gen. Virol. 2001, 82 (Pt 3), 499–506. [Google Scholar] [CrossRef]
- Olp, L.N.; Jeanniard, A.; Marimo, C.; West, J.T.; Wood, C. Whole-Genome Sequencing of Kaposi’s Sarcoma-Associated Herpesvirus from Zambian Kaposi’s Sarcoma Biopsy Specimens Reveals Unique Viral Diversity. J. Virol. 2015, 89, 12299–12308. [Google Scholar] [CrossRef] [Green Version]
- Tornesello, M.L.; Biryahwaho, B.; Downing, R.; Hatzakis, A.; Alessi, E.; Cusini, M.; Ruocco, V.; Katongole-Mbidde, E.; Loquercio, G.; Buonaguro, L.; et al. Human herpesvirus type 8 variants circulating in Europe, Africa and North America in classic, endemic and epidemic Kaposi’s sarcoma lesions during pre-AIDS and AIDS era. Virology 2010, 398, 280–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duprez, R.; Hbid, O.; Afonso, P.; Quach, H.; Belloul, L.; Fajali, N.; Ismaili, N.; Benomar, H.; Tahri, E.H.; Huerre, M.; et al. Molecular epidemiology of the HHV-8 K1 gene from Moroccan patients with Kaposi’s sarcoma. Virology 2006, 353, 121–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.J.; Davis, T.L.; Wang, X.P.; Deng, J.H.; Baillargeon, J.; Yeh, I.T.; Jenson, H.B.; Gao, S.J. Distinct distribution of rare US genotypes of Kaposi’s sarcoma-associated herpesvirus (KSHV) in South Texas: Implications for KSHV epidemiology. J. Infect. Dis. 2001, 183, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Kamiyama, K.; Kinjo, T.; Chinen, K.; Iwamasa, T.; Uezato, H.; Miyagi, J.I.; Mori, N.; Yamane, N. Human herpesvirus 8 (HHV8) sequence variations in HHV8 related tumours in Okinawa, a subtropical island in southern Japan. J. Clin. Pathol. 2004, 57, 529–535. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Pu, X.; Wu, W.; Jin, Y.; Juhear, M.; Wu, X. Genotypic analysis on the ORF-K1 gene of human herpesvirus 8 from patients with Kaposi’s sarcoma in Xinjiang, China. J. Genet. Genom. 2008, 35, 657–663. [Google Scholar] [CrossRef]
- Fouchard, N.; Lacoste, V.; Couppie, P.; Develoux, M.; Mauclere, P.; Michel, P.; Herve, V.; Pradinaud, R.; Bestetti, G.; Huerre, M.; et al. Detection and genetic polymorphism of human herpes virus type 8 in endemic or epidemic Kaposi’s sarcoma from West and Central Africa, and South America. Int. J. Cancer 2000, 85, 166–170. [Google Scholar] [CrossRef]
- Kasolo, F.C.; Spinks, J.; Bima, H.; Bates, M.; Gompels, U.A. Diverse genotypes of Kaposi’s sarcoma associated herpesvirus (KSHV) identified in infant blood infections in African childhood-KS and HIV/AIDS endemic region. J. Med. Virol. 2007, 79, 1555–1561. [Google Scholar] [CrossRef] [Green Version]
- White, T.; Hagen, M.; Gudza, I.; White, I.E.; Ndemera, B.; Gwanzura, L.; Borok, M.; Campbell, T.B. Genetic diversity of the Kaposi’s sarcoma herpesvirus K1 protein in AIDS-KS in Zimbabwe. J. Clin. Virol. 2008, 42, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Cassar, O.; Charavay, F.; Bassot, S.; Plancoulaine, S.; Grangeon, J.P.; Laumond-Barny, S.; Martin, P.M.; Chanteau, S.; Gessain, A. Divergent KSHV/HHV-8 subtype D strains in New Caledonia and Solomon Islands, Melanesia. J. Clin. Virol. 2012, 53, 214–218. [Google Scholar] [CrossRef]
- Capan-Melser, M.; Mombo-Ngoma, G.; Akerey-Diop, D.; Basra, A.; Manego-Zoleko, R.; Wurbel, H.; Lotsch, F.; Groger, M.; Skoll, M.; Schwing, J.; et al. Epidemiology of Human Herpes Virus 8 in Pregnant Women and their Newborns--A cross-sectional delivery survey in Central Gabon. Int. J. Infect. Dis. 2015, 39, 16–19. [Google Scholar] [CrossRef] [Green Version]
- Etta, E.M.; Alayande, D.P.; Mavhandu-Ramarumo, L.G.; Gachara, G.; Bessong, P.O. HHV-8 Seroprevalence and Genotype Distribution in Africa, 1998–2017: A Systematic Review. Viruses 2018, 10, 458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djuicy, D.D.; Mouinga-Ondeme, A.; Cassar, O.; Ramassamy, J.L.; Idam Mamimandjiami, A.; Bikangui, R.; Fontanet, A.; Gessain, A. Risk factors for HTLV-1 infection in Central Africa: A rural population-based survey in Gabon. PLoS Negl. Trop Dis. 2018, 12, e0006832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arvanitakis, L.; Mesri, E.A.; Nador, R.G.; Said, J.W.; Asch, A.S.; Knowles, D.M.; Cesarman, E. Establishment and characterization of a primary effusion (body cavity-based) lymphoma cell line (BC-3) harboring kaposi’s sarcoma-associated herpesvirus (KSHV/HHV-8) in the absence of Epstein-Barr virus. Blood 1996, 88, 2648–2654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadyrova, E.; Lacoste, V.; Duprez, R.; Pozharissky, K.; Molochkov, V.; Huerre, M.; Gurtsevitch, V.; Gessain, A. Molecular epidemiology of Kaposi’s sarcoma-associated herpesvirus/human herpesvirus 8 strains from Russian patients with classic, posttransplant, and AIDS-associated Kaposi’s sarcoma. J. Med. Virol. 2003, 71, 548–556. [Google Scholar] [CrossRef] [PubMed]
- Stebbing, J.; Bourboulia, D.; Johnson, M.; Henderson, S.; Williams, I.; Wilder, N.; Tyrer, M.; Youle, M.; Imami, N.; Kobu, T.; et al. Kaposi’s sarcoma-associated herpesvirus cytotoxic T lymphocytes recognize and target Darwinian positively selected autologous K1 epitopes. J. Virol. 2003, 77, 4306–4314. [Google Scholar] [CrossRef] [Green Version]
- Dedicoat, M.; Newton, R. Review of the distribution of Kaposi’s sarcoma-associated herpesvirus (KSHV) in Africa in relation to the incidence of Kaposi’s sarcoma. Br. J. Cancer 2003, 88, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Bagni, R.; Whitby, D. Kaposi’s sarcoma-associated herpesvirus transmission and primary infection. Curr. Opin. HIV AIDS 2009, 4, 22–26. [Google Scholar] [CrossRef]
- Crabtree, K.L.; Wojcicki, J.M.; Minhas, V.; Kankasa, C.; Mitchell, C.; Wood, C. Association of Household Food- and Drink-Sharing Practices with Human Herpesvirus 8 Seroconversion in a Cohort of Zambian Children. J. Infect. Dis. 2017, 216, 842–849. [Google Scholar] [CrossRef]
- Minhas, V.; Wood, C. Epidemiology and transmission of Kaposi’s sarcoma-associated herpesvirus. Viruses 2014, 6, 4178–4194. [Google Scholar] [CrossRef] [Green Version]
- Plancoulaine, S.; Abel, L.; Tregouet, D.; Duprez, R.; van Beveren, M.; Tortevoye, P.; Froment, A.; Gessain, A. Respective roles of serological status and blood specific antihuman herpesvirus 8 antibody levels in human herpesvirus 8 intrafamilial transmission in a highly endemic area. Cancer Res. 2004, 64, 8782–8787. [Google Scholar] [CrossRef] [Green Version]
- Newton, R.; Labo, N.; Wakeham, K.; Marshall, V.; Roshan, R.; Nalwoga, A.; Sebina, I.; Muhangi, L.; Webb, E.L.; Miley, W.; et al. Determinants of Gammaherpesvirus Shedding in Saliva Among Ugandan Children and Their Mothers. J. Infect. Dis. 2018, 218, 892–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezza, G.; Tchangmena, O.B.; Andreoni, M.; Bugarini, R.; Toma, L.; Bakary, D.K.; Glikoutou, M.; Sarmati, L.; Monini, P.; Pezzotti, P.; et al. Prevalence and risk factors for human herpesvirus 8 infection in northern Cameroon. Sex. Transm. Dis. 2000, 27, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Corchero, J.L.; Mar, E.C.; Spira, T.J.; Pellett, P.E.; Inoue, N. Comparison of serologic assays for detection of antibodies against human herpesvirus 8. Clin. Diagn. Lab. Immunol. 2001, 8, 913–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudnall, S.D.; Chen, T.; Rady, P.; Tyring, S.; Allison, P. Human herpesvirus 8 seroprevalence and viral load in healthy adult blood donors. Transfusion 2003, 43, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Duprez, R.; Kassa-Kelembho, E.; Plancoulaine, S.; Briere, J.; Fossi, M.; Kobangue, L.; Minsart, P.; Huerre, M.; Gessain, A. Human herpesvirus 8 serological markers and viral load in patients with AIDS-associated Kaposi’s sarcoma in Central African Republic. J. Clin. Microbiol. 2005, 43, 4840–4843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dedicoat, M.; Newton, R.; Alkharsah, K.R.; Sheldon, J.; Szabados, I.; Ndlovu, B.; Page, T.; Casabonne, D.; Gilks, C.F.; Cassol, S.A.; et al. Mother-to-child transmission of human herpesvirus-8 in South Africa. J. Infect. Dis. 2004, 190, 1068–1075. [Google Scholar] [CrossRef] [Green Version]
- Dow, D.E.; Cunningham, C.K.; Buchanan, A.M. A Review of Human Herpesvirus 8, the Kaposi’s Sarcoma-Associated Herpesvirus, in the Pediatric Population. J. Pediatric Infect. Dis. Soc. 2014, 3, 66–76. [Google Scholar] [CrossRef] [Green Version]
- Mayama, S.; Cuevas, L.E.; Sheldon, J.; Omar, O.H.; Smith, D.H.; Okong, P.; Silvel, B.; Hart, C.A.; Schulz, T.F. Prevalence and transmission of Kaposi’s sarcoma-associated herpesvirus (human herpesvirus 8) in Ugandan children and adolescents. Int. J. Cancer 1998, 77, 817–820. [Google Scholar] [CrossRef]
- Mbulaiteye, S.M.; Pfeiffer, R.M.; Whitby, D.; Brubaker, G.R.; Shao, J.; Biggar, R.J. Human herpesvirus 8 infection within families in rural Tanzania. J. Infect. Dis. 2003, 187, 1780–1785. [Google Scholar] [CrossRef]
- Sarmati, L. HHV-8 infection in African children. Herpes 2004, 11, 50–53. [Google Scholar]
- Mbulaiteye, S.M.; Pfeiffer, R.M.; Engels, E.A.; Marshall, V.; Bakaki, P.M.; Owor, A.M.; Ndugwa, C.M.; Katongole-Mbidde, E.; Goedert, J.J.; Biggar, R.J.; et al. Detection of kaposi sarcoma-associated herpesvirus DNA in saliva and buffy-coat samples from children with sickle cell disease in Uganda. J. Infect. Dis. 2004, 190, 1382–1386. [Google Scholar] [CrossRef] [PubMed]
- Jary, A.; Leducq, V.; Desire, N.; Petit, H.; Palich, R.; Joly, V.; Canestri, A.; Gothland, A.; Lambert-Niclot, S.; Surgers, L.; et al. New Kaposi’s sarcoma-associated herpesvirus variant in men who have sex with men associated with severe pathologies. J. Infect. Dis. 2020, 222, 1320–1328. [Google Scholar] [CrossRef] [PubMed]
- Cassar, O.; Desrames, A.; Marcais, A.; Gout, O.; Taylor, G.P.; Hermine, O.; Soriano, V.; Mendoza, C.; Dehan, O.; Mener, M.L.; et al. Multiple recombinant events in human T-cell Leukemia virus Type 1: Complete sequences of recombinant African strains. Emerg. Microbes Infect. 2020, 9, 913–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zong, J.; Ciufo, D.M.; Viscidi, R.; Alagiozoglou, L.; Tyring, S.; Rady, P.; Orenstein, J.; Boto, W.; Kalumbuja, H.; Romano, N.; et al. Genotypic analysis at multiple loci across Kaposi’s sarcoma herpesvirus (KSHV) DNA molecules: Clustering patterns, novel variants and chimerism. J. Clin. Virol. 2002, 23, 119–148. [Google Scholar] [CrossRef]
- Sallah, N.; Palser, A.L.; Watson, S.J.; Labo, N.; Asiki, G.; Marshall, V.; Newton, R.; Whitby, D.; Kellam, P.; Barroso, I. Genome-Wide Sequence Analysis of Kaposi Sarcoma-Associated Herpesvirus Shows Diversification Driven by Recombination. J. Infect. Dis. 2018, 218, 1700–1710. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| n | HHV-8 Serological Results (IFA) | HHV-8 Molecular Results (PCR) | |||||
|---|---|---|---|---|---|---|---|
| nIFA 1:160 | Seroprevalence (95%CI) | p-Value | nPCR K1 | Percentage of Positive PCR (95%CI) | p-Value | ||
| Sex | |||||||
| Female | 388 | 143 | 36.9% (32.0–41.9) | 0.84 | 10 | 2.6% (1.2–4.7) | 0.96 |
| Male | 632 | 229 | 36.2% (32.5–40.1) | 16 | 2.5% (1.5–4.1) | ||
| Age category | |||||||
| 15–35 | 238 | 85 | 35.7% (29.6–42.2) | 0.65 | 4 | 1.7% (0.5–4.2) | 0.44 |
| 36–50 | 250 | 86 | 34.4% (28.5–40.6) | 8 | 3.2% (1.4–6.2) | ||
| 51–65 | 276 | 109 | 39.5% (33.7–45.5) | 5 | 1.8% (0.6–4.2) | ||
| 66–95 | 256 | 92 | 35.9% (30.1–42.1) | 9 | 3.5% (1.6–6.6) | ||
| Ethnic group | |||||||
| Bantus | 931 | 342 | 36.8% (33.6–39.9) | 0.57 | 26 | 2.8% (1.8–4.1) | - |
| Pygmies | 89 | 30 | 33.7% (24.0–44.5) | 0 | - | ||
| Province | |||||||
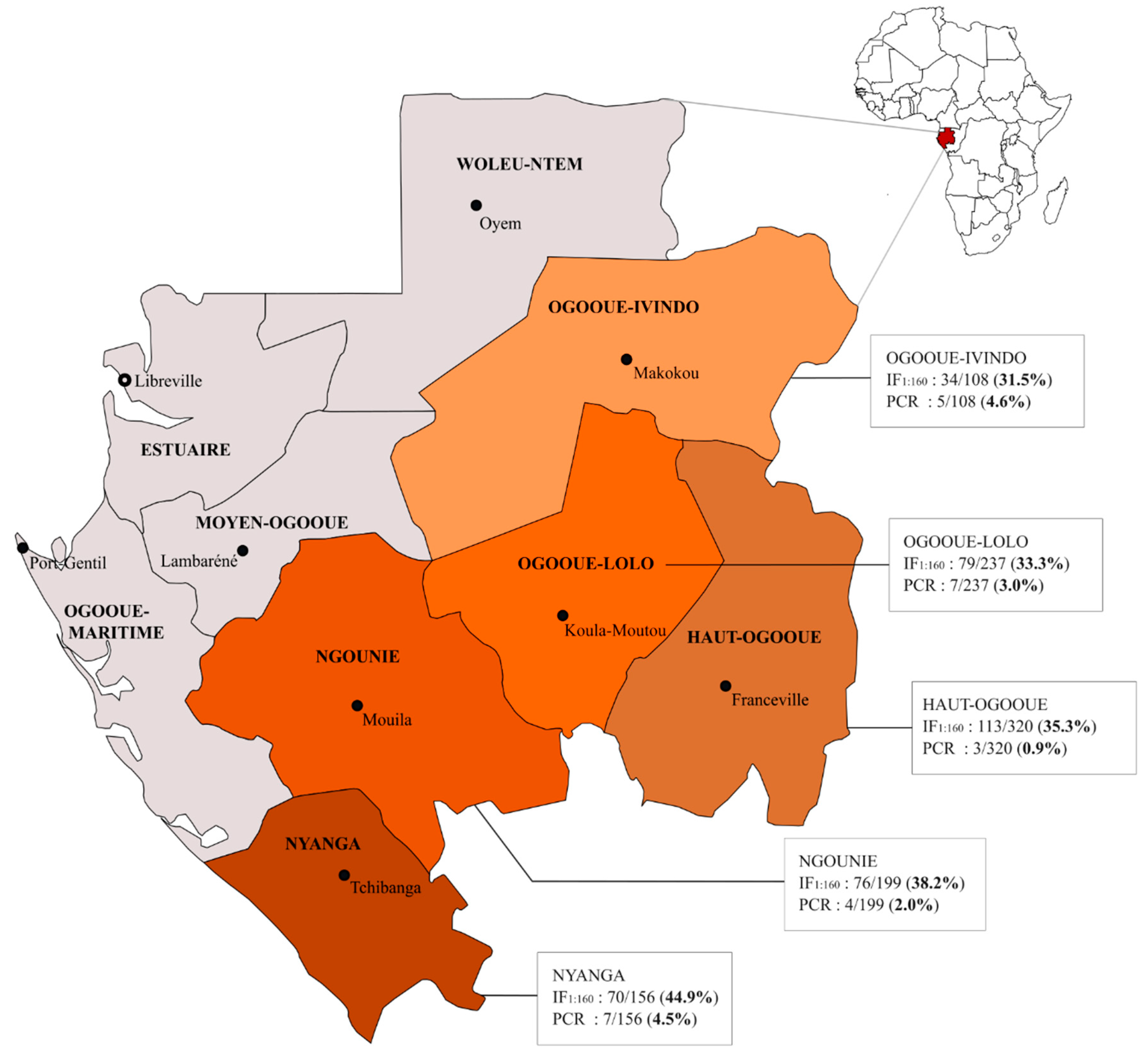
| Haut-Ogooué | 320 | 113 | 35.3% (30.1–40.8) | 0.12 | 3 | 0.9% (0.2–2.7) | 0.09 |
| Ngounié | 199 | 76 | 38.2% (31.4–45.3) | 4 | 2.0% (0.6–5.1) | ||
| Nyanga | 156 | 70 | 44.9% (36.9–53.0) | 7 | 4.5% (1.8–9.0) | ||
| Ogooué-Ivindo | 108 | 34 | 31.5% (22.9–41.1) | 5 | 4.6% (1.5–10.5) | ||
| Ogooué-Lolo | 237 | 79 | 33.3% (27.4–39.7) | 7 | 3.0% (1.2–6.0) | ||
| HTLV-1 status | |||||||
| Non infected | 896 | 333 | 37.2% (34.0–40.4) | 0.22 | 20 | 2.2% (1.4–3.4) | 0.08 |
| Infected | 124 | 39 | 31.5% (23.4–40.4) | 6 | 4.8% (1.8–10.2) | ||
| Total | 1020 | 372 | 36.5% (33.5–39.5) | 26 | 2.5% (1.7–3.7) | ||
| HHV-8 Serological Results | n | HHV-8 Sero-Prevalence (95%CI) | n PCR + K1 | Percentage of Positive PCR (95%CI) | p-Value |
|---|---|---|---|---|---|
| IFA 1:40 | |||||
| Positive | 493 | 48.3% (45.2–51.5) | 19 | 3.9% (2.3–6.0) | |
| Negative | 527 | 7 | 1.3% (0.5–2.7) | 0.01 | |
| IFA 1:80 | |||||
| Positive | 455 | 44.6% (41.5–47.7) | 17 | 3.7% (2.2–5.9) | |
| Negative | 565 | 9 | 1.6% (0.7–3.0) | 0.03 | |
| IFA 1:160 | |||||
| Positive | 372 | 36.5% (33.5–39.5) | 13 | 3.5% (1.9–5.9) | |
| Negative | 648 | 13 | 2.0% (1.1–3.4) | 0.15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mamimandjiami, A.I.; Mouinga-Ondémé, A.; Ramassamy, J.-L.; Djuicy, D.D.; Afonso, P.V.; Mahé, A.; Lekana-Douki, J.-B.; Cassar, O.; Gessain, A. Epidemiology and Genetic Variability of HHV-8/KSHV among Rural Populations and Kaposi’s Sarcoma Patients in Gabon, Central Africa. Review of the Geographical Distribution of HHV-8 K1 Genotypes in Africa. Viruses 2021, 13, 175. https://0-doi-org.brum.beds.ac.uk/10.3390/v13020175
Mamimandjiami AI, Mouinga-Ondémé A, Ramassamy J-L, Djuicy DD, Afonso PV, Mahé A, Lekana-Douki J-B, Cassar O, Gessain A. Epidemiology and Genetic Variability of HHV-8/KSHV among Rural Populations and Kaposi’s Sarcoma Patients in Gabon, Central Africa. Review of the Geographical Distribution of HHV-8 K1 Genotypes in Africa. Viruses. 2021; 13(2):175. https://0-doi-org.brum.beds.ac.uk/10.3390/v13020175
Chicago/Turabian StyleMamimandjiami, Antony Idam, Augustin Mouinga-Ondémé, Jill-Léa Ramassamy, Délia Doreen Djuicy, Philippe V. Afonso, Antoine Mahé, Jean-Bernard Lekana-Douki, Olivier Cassar, and Antoine Gessain. 2021. "Epidemiology and Genetic Variability of HHV-8/KSHV among Rural Populations and Kaposi’s Sarcoma Patients in Gabon, Central Africa. Review of the Geographical Distribution of HHV-8 K1 Genotypes in Africa" Viruses 13, no. 2: 175. https://0-doi-org.brum.beds.ac.uk/10.3390/v13020175