
Selective Interaction of Sugarcane eIF4E with VPgs from Sugarcane Mosaic Pathogens


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Viruses
2.2. RNA Extraction and Real-Time Quantitative PCR
2.3. Plasmid Construction
2.4. Protein Interaction as Determined by Y2H and BiFC Assays
2.5. Transient Expression
2.6. Confocal Microscopy
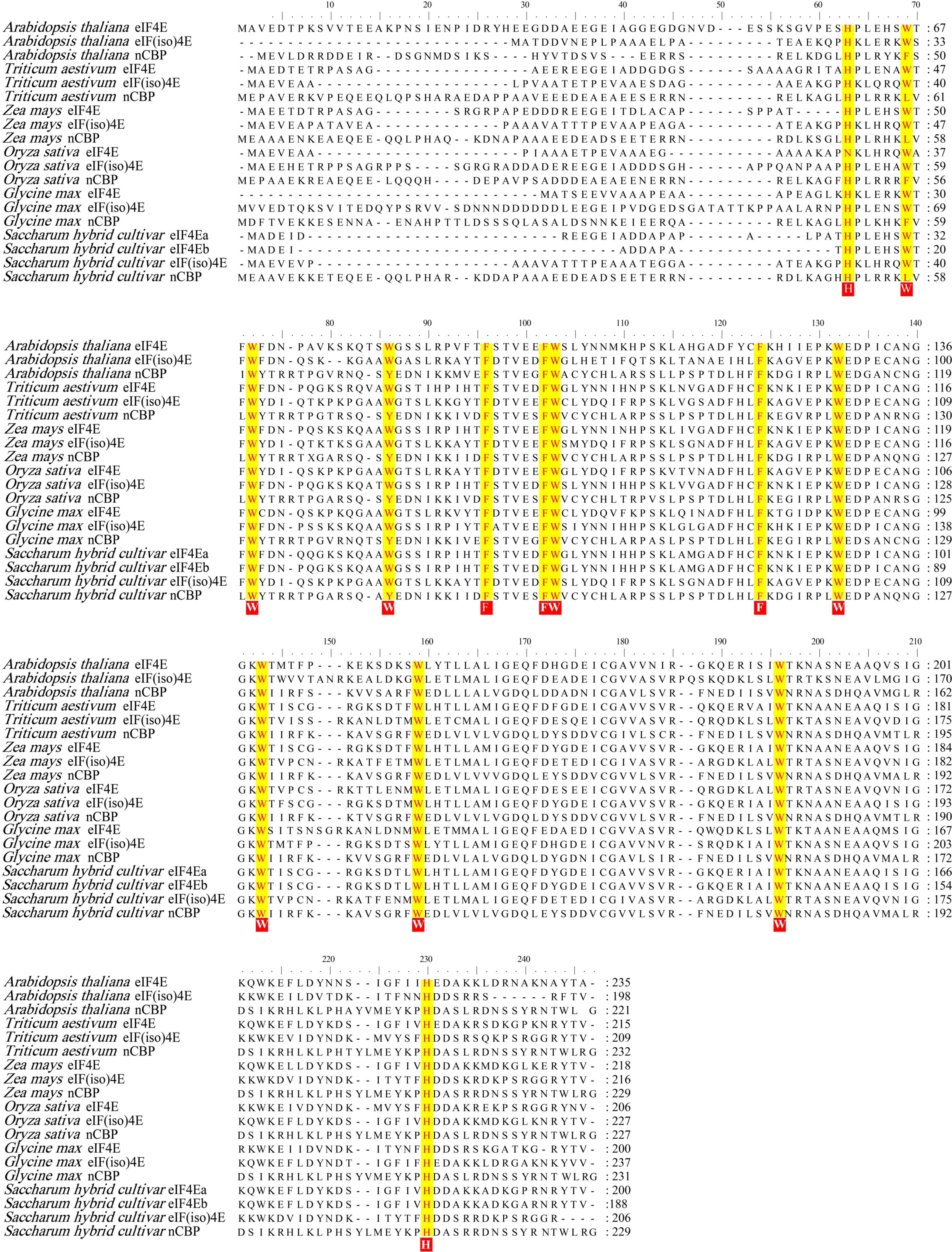
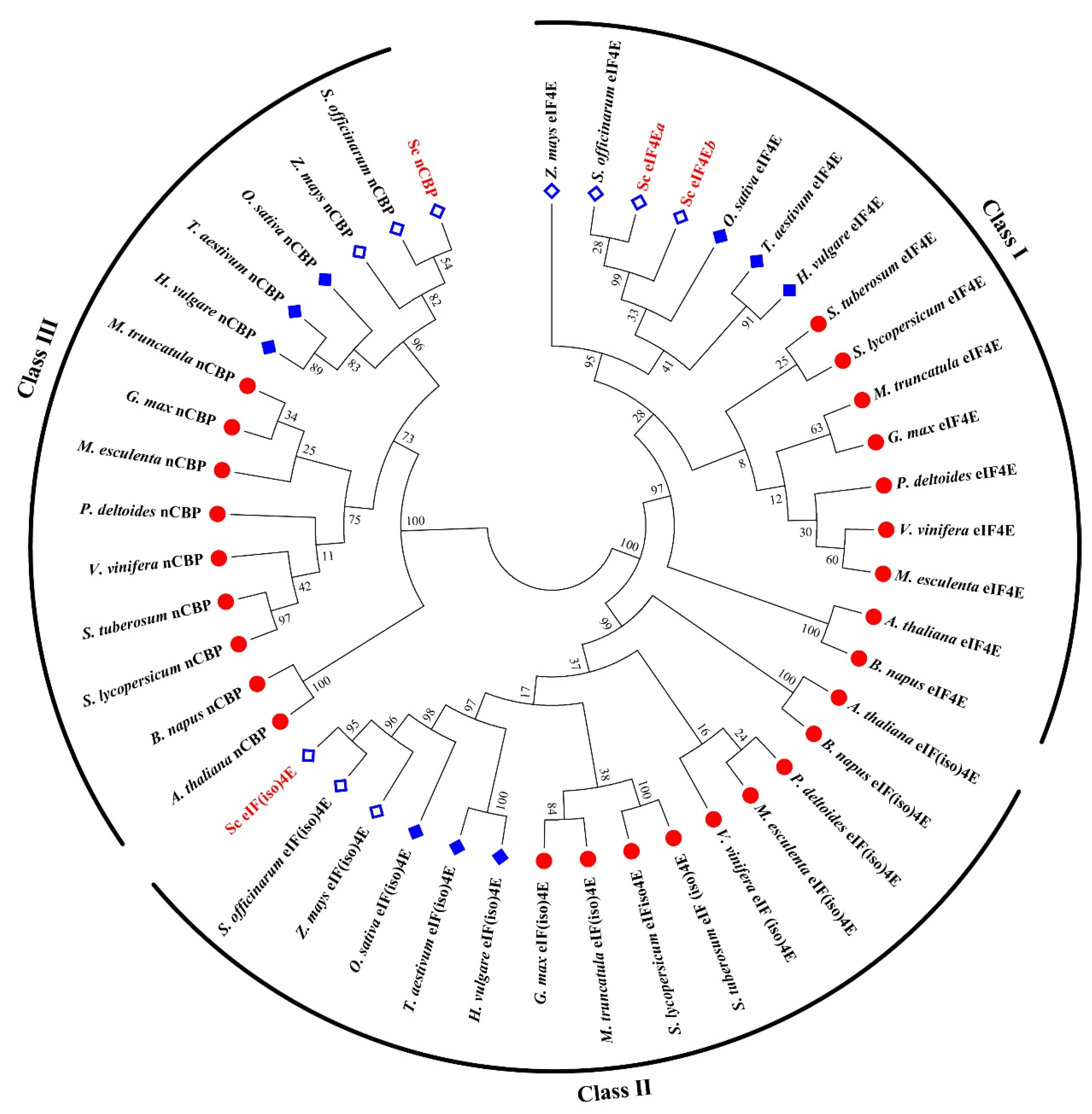
2.7. Multiple Sequence Alignment
3. Results
3.1. Cloning of SceIF4Ea, SceIF4Eb, SceIF(iso)4E, and ScnCBP from Sugarcane
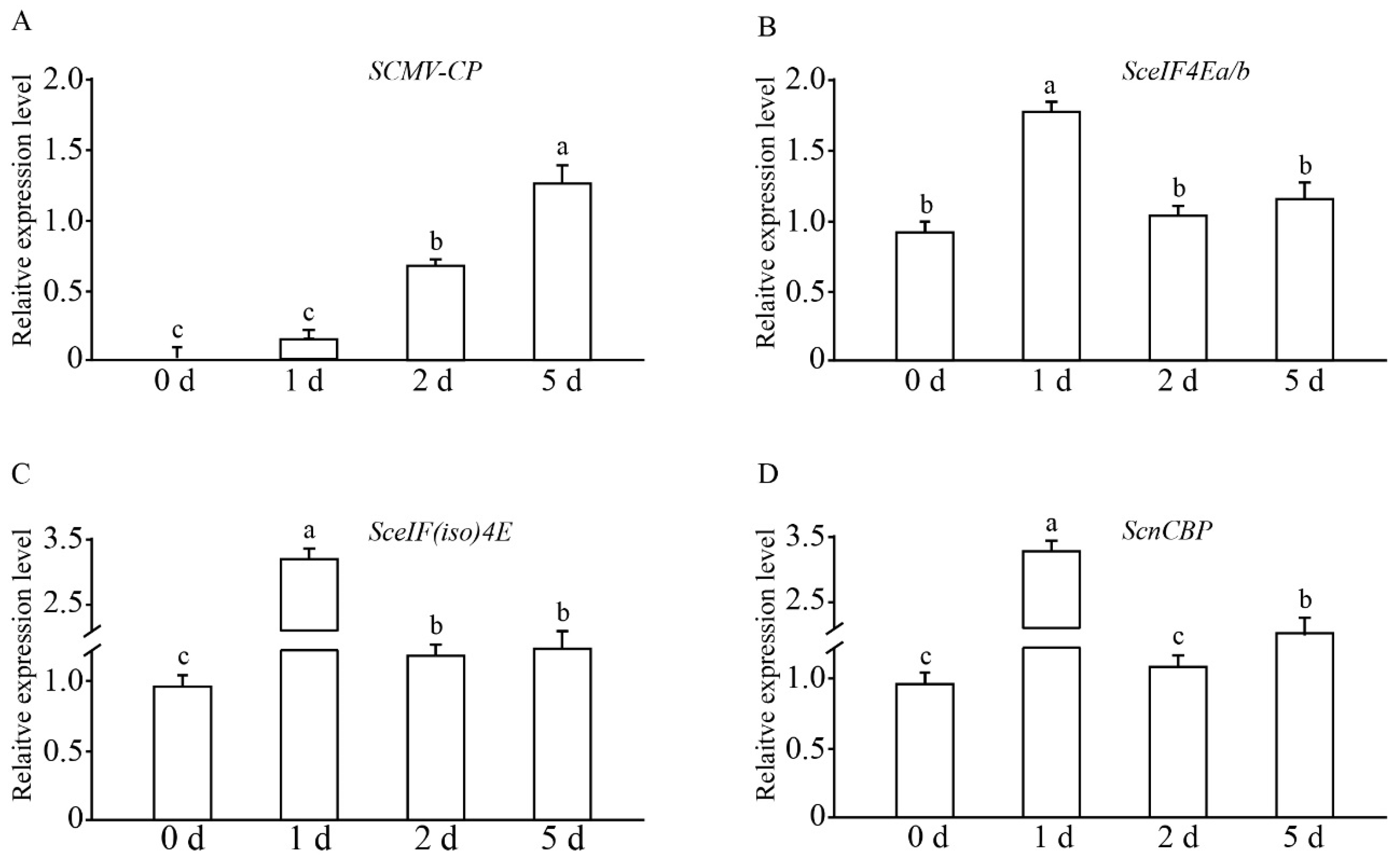
3.2. Expression Profiles of SceIF4Ea, SceIF4Eb, SceIF(iso)4E, and ScnCBP Upon the SCMV Challenge
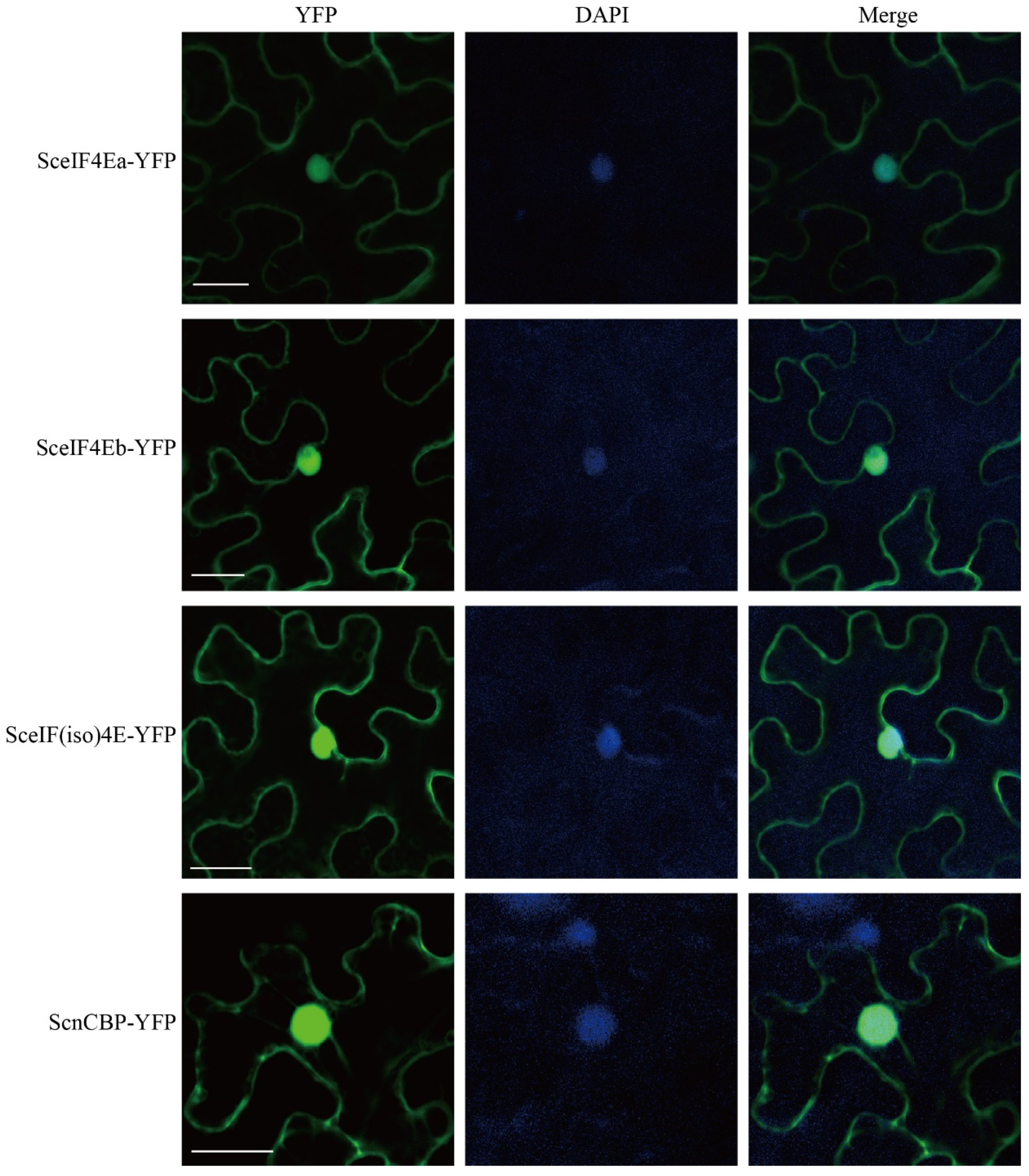
3.3. Subcellular Localization of SceIF4Ea, SceIF4Eb, SceIF(iso)4E, and ScnCBP
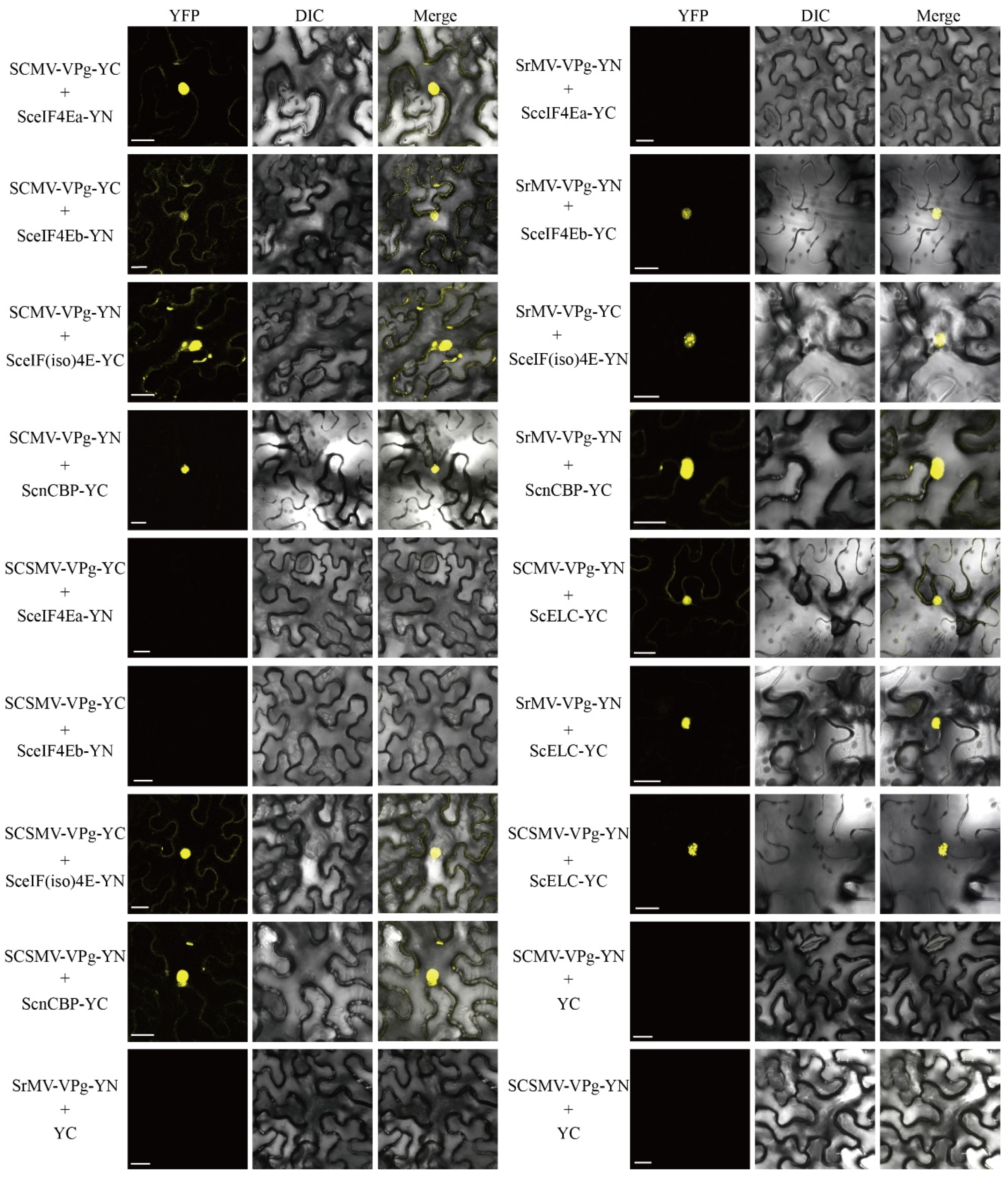
3.4. Interaction of SceIF4Ea, SceIF4Eb, SceIF(iso)4E, and ScnCBP with SCMV-/SrMV-/SCSMV-VPg
3.5. Competitive Interaction of SceIF4Ea, SceIF4Eb, SceIF(iso)4E with SCMV-/SrMV-/SCSMV-VPg
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rae, A.; Jackson, M.; Nguyen, C.H.; Bonnett, G.D. Functional Specialization of Vacuoles in Sugarcane Leaf and Stem. Trop. Plant Biol. 2008, 2, 13–22. [Google Scholar] [CrossRef]
- Lam, E.; Shine, J.; Da Silva, J.; Lawton, M.; Bonos, S.; Calvino, M.; Carrer, H.; Silva-Filho, M.C.; Glynn, N.; Helsel, Z.; et al. Improving sugarcane for biofuel: Engineering for an even better feedstock. GCB Bioenergy 2009, 1, 251–255. [Google Scholar] [CrossRef]
- Sabatier, D.; Martiné, J.; Chiroleu, F.; Roussel, C.; Letourmy, P.; Antwerpen, R.V.; Gabrielle, B.; Ney, B. Optimization of sugarcane farming as a multipurpose crop for energy and food production. GCB Bioenergy 2015, 7. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.J.; Zu, X.F.; Wang, S.X.; Chen, Y.H. Sugarcane mosaic virus—Long history but still a threat to industry. Crop Prot. 2012, 42, 74–78. [Google Scholar] [CrossRef]
- Zambrano, A.Y.; Demey, J.R.; Fuchs, M.; Gonzalez, V.; Rea, R.; De Sousa, O.; Gutierrez, Z. Selection of sugarcane plants resistant to SCMV. Plant Sci. 2003, 165, 221–225. [Google Scholar] [CrossRef]
- Xie, Y.J.; Wang, M.Q.; Xu, D.L.; Li, R.H.; Zhou, G.H. Simultaneous detection and identification of four sugarcane viruses by one-step RT-PCR. J. Virol. Methods 2009, 162, 64–68. [Google Scholar] [CrossRef]
- Yang, Z.N.; Mirkov, T.E. Sequence and Relationships of Sugarcane Mosaic and Sorghum Mosaic Virus Strains and Development of RT-PCR-Based RFLPs for Strain Discrimination. Phytopathology 1997, 87, 932–939. [Google Scholar] [CrossRef] [Green Version]
- Grisham, M.P.; Pan, Y.B. A genetic shift in the virus strains that cause mosaic in Louisiana sugarcane. Plant Dis. 2007, 91, 453–458. [Google Scholar] [CrossRef] [Green Version]
- Perera, M.F.; Filippone, M.P.; Ramallo, C.J.; Cuenya, M.I.; Garcia, M.L.; Ploper, L.D.; Castagnaro, A.P. Genetic Diversity Among Viruses Associated with Sugarcane Mosaic Disease in Tucumaan, Argentina. Phytopathology 2009, 99, 38–49. [Google Scholar] [CrossRef] [PubMed]
- Ha, C.; Revill, P.; Harding, R.M.; Vu, M.; Dale, J.L. Identification and sequence analysis of potyviruses infecting crops in Vietnam. Arch. Virol. 2008, 153, 45–60. [Google Scholar] [CrossRef]
- Li, W.F.; Wang, X.Y.; Huang, Y.K.; Shan, H.L.; Luo, Z.M.; Ying, X.M.; Zhang, R.Y.; Shen, K.; Yin, J. Screening sugarcane germplasm resistant to Sorghum mosaic virus. Crop Prot. 2013, 43, 27–30. [Google Scholar] [CrossRef]
- Li, W.F.; Shan, H.L.; Zhang, R.Y.; Wang, X.Y.; Yang, K.; Luo, Z.M.; Yin, J.; Cang, X.Y.; Li, J.; Huang, Y.K. Identification of resistance to Sugarcane streak mosaic virus (SCSMV) and Sorghum mosaic virus (SrMV) in new elite sugarcane varieties/clones in China. Crop Prot. 2018, 110, 77–82. [Google Scholar] [CrossRef]
- Li, W.F.; Shan, H.L.; Cang, X.Y.; Lu, X.; Zhang, R.Y.; Wang, X.Y.; Yin, J.; Luo, Z.M.; Huang, Y.K. Identification and Evaluation of Resistance to Sugarcane Streak Mosaic Virus (SCSMV) and Sorghum Mosaic Virus (SrMV) in Excellent Sugarcane Innovation Germplasms in China. Sugar Tech 2019, 21, 481–485. [Google Scholar] [CrossRef]
- Filloux, D.; Fernandez, E.; Comstock, J.C.; Mollov, D.; Roumagnac, P.; Rott, P. Viral Metagenomic-Based Screening of Sugarcane from Florida Reveals Occurrence of Six Sugarcane-Infecting Viruses and High Prevalence of Sugarcane yellow leaf virus. Plant Dis. 2018, 102, 2317–2323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akbar, S.; Yao, W.; Yu, K.; Qin, L.F.; Ruan, M.H.; Powell, C.A.; Chen, B.S.; Zhang, M.Q. Photosynthetic characterization and expression profiles of sugarcane infected by Sugarcane mosaic virus (SCMV). Photosynth. Res. 2020. [Google Scholar] [CrossRef]
- Putra, L.K.; Kristini, A.; Achadian, E.M.; Damayanti, T.A. Sugarcane streak mosaic virus in Indonesia: Distribution, Characterisation, Yield Losses and Management Approaches. Sugar Tech 2014, 16, 392–399. [Google Scholar] [CrossRef]
- Yao, W.; Ruan, M.; Qin, L.; Yang, C.; Chen, R.; Chen, B.; Zhang, M. Field Performance of Transgenic Sugarcane Lines Resistant to Sugarcane Mosaic Virus. Front. Plant Sci. 2017, 8, 104. [Google Scholar] [CrossRef] [Green Version]
- Ricaud, C.; Egan, B.; Gillaspie, A.G.; Hughes, C.G. Diseases of Sugarcane: Major Diseases; Elsevier: New York, NY, USA, 1989; pp. 301–322. [Google Scholar]
- Charron, C.; Nicolai, M.; Gallois, J.L.; Robaglia, C.; Moury, B.T.; Palloix, A.; Caranta, C. Natural variation and functional analyses provide evidence for co-evolution between plant eIF4E and potyviral VPg. Plant J. 2008, 54, 56–68. [Google Scholar] [CrossRef]
- Ward, C.; Shukla, D.J.I. Taxonomy of potyviruses: Current problems and some solutions. Intervirology 1991, 32 5, 269–296. [Google Scholar] [CrossRef]
- Hema, M.; Sreenivasulu, P.; Savithri, H.S. Taxonomic position of Sugarcane streak mosaic virus in the family Potyviridae. Arch Virol. 2002, 147, 1997–2007. [Google Scholar] [CrossRef]
- Xu, D.L.; Zhou, G.H.; Xie, Y.J.; Mock, R.; Li, R. Complete nucleotide sequence and taxonomy of Sugarcane streak mosaic virus, member of a novel genus in the family Potyviridae. Virus Genes 2010, 40, 432–439. [Google Scholar] [CrossRef]
- Li, W.; He, Z.; Li, S.; Huang, Y.; Zhang, Z.; Jiang, D.; Wang, X.; Luo, Z. Molecular characterization of a new strain of Sugarcane streak mosaic virus (SCSMV). Arch Virol. 2011, 156, 2101–2104. [Google Scholar] [CrossRef] [PubMed]
- Urcuqui-Inchima, S.; Haenni, A.L.; Bernardi, F. Potyvirus proteins: A wealth of functions. Virus Res. 2001, 74, 157–175. [Google Scholar] [CrossRef]
- Wang, A.M. Dissecting the Molecular Network of Virus-Plant Interactions: The Complex Roles of Host Factors. Annu. Rev. Phytopathol. 2015, 53, 45–66. [Google Scholar] [CrossRef] [PubMed]
- Olspert, A.; Chung, B.Y.; Atkins, J.F.; Carr, J.P.; Firth, A.E. Transcriptional slippage in the positive-sense RNA virus family Potyviridae. EMBO Rep. 2015, 16, 995–1004. [Google Scholar] [CrossRef] [PubMed]
- Olspert, A.; Carr, J.P.; Firth, A.E. Mutational analysis of the Potyviridae transcriptional slippage site utilized for expression of the P3N-PIPO and P1N-PISPO proteins. Nucleic Acids Res. 2016, 44, 7618–7629. [Google Scholar] [CrossRef] [Green Version]
- Riechmann, J.L.; Laín, S.; García, J.A. Highlights and prospects of potyvirus molecular biology. J. Gen. Virol. 1992, 73 Pt 1, 1–16. [Google Scholar] [CrossRef]
- Cui, X.; Yaghmaiean, H.; Wu, G.; Wu, X.; Chen, X.; Thorn, G.; Wang, A. The C-terminal region of the Turnip mosaic virus P3 protein is essential for viral infection via targeting P3 to the viral replication complex. Virology 2017, 510, 147–155. [Google Scholar] [CrossRef]
- Cheng, G.; Dong, M.; Xu, Q.; Peng, L.; Yang, Z.; Wei, T.; Xu, J. Dissecting the Molecular Mechanism of the Subcellular Localization and Cell-to-cell Movement of the Sugarcane mosaic virus P3N-PIPO. Sci. Rep. 2017, 7, 9868. [Google Scholar] [CrossRef]
- Cheng, G.; Yang, Z.; Zhang, H.; Zhang, J.; Xu, J. Remorin interacting with PCaP1 impairs Turnip mosaic virus intercellular movement but is antagonised by VPg. New Phytol. 2020, 225, 2122–2139. [Google Scholar] [CrossRef]
- Chai, M.; Wu, X.; Liu, J.; Fang, Y.; Luan, Y.; Cui, X.; Zhou, X.; Wang, A.; Cheng, X. P3N-PIPO Interacts with P3 via the Shared N-Terminal Domain to Recruit Viral Replication Vesicles for Cell-to-Cell Movement. J. Virol. 2020, 94. [Google Scholar] [CrossRef] [PubMed]
- Gingras, A.C.; Raught, B.; Sonenberg, N. eIF4 initiation factors: Effectors of mRNA recruitment to ribosomes and regulators of translation. Annu. Rev. Biochem. 1999, 68, 913–963. [Google Scholar] [CrossRef] [PubMed]
- Sonenberg, N.; Hershey, J.; Mathews, M. Translational Control of Gene Expression; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2000; pp. 1–31. [Google Scholar]
- Wittmann, S.; Chatel, H.; Fortin, M.G.; Laliberté, J.F. Interaction of the viral protein genome linked of turnip mosaic potyvirus with the translational eukaryotic initiation factor (iso) 4E of Arabidopsis thaliana using the yeast two-hybrid system. Virology 1997, 234, 84–92. [Google Scholar] [CrossRef] [Green Version]
- Leonard, S.; Plante, D.; Wittmann, S.; Daigneault, N.; Fortin, M.; Laliberté, J.F. Complex Formation between Potyvirus VPg and Translation Eukaryotic Initiation Factor 4E Correlates with Virus Infectivity. J. Virol. 2000, 74, 7730–7737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, A.M.; Krishnaswamy, S. Eukaryotic translation initiation factor 4E-mediated recessive resistance to plant viruses and its utility in crop improvement. Mol. Plant Pathol. 2012, 13, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Anindya, R.; Chittori, S.; Savithri, H.S. Tyrosine 66 of Pepper vein banding virus genome-linked protein is uridylylated by RNA-dependent RNA polymerase. Virology 2005, 336, 154–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belliot, G.; Sosnovtsev, S.V.; Chang, K.-O.; McPhie, P.; Green, K.Y. Nucleotidylylation of the VPg protein of a human norovirus by its proteinase-polymerase precursor protein. Virology 2008, 374, 33–49. [Google Scholar] [CrossRef] [Green Version]
- Murphy, J.F.; Rychlik, W.; Rhoads, R.E.; Hunt, A.G.; Shaw, J.G. A tyrosine residue in the small nuclear inclusion protein of tobacco vein mottling virus links the VPg to the viral RNA. J. Virol. 1991, 65, 511–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, J.; Klein, P.G.; Hunt, A.; Shaw, J.J.V. Replacement of the tyrosine residue that links a potyviral VPg to the viral RNA is lethal. Virology 1996, 220 2, 535–538. [Google Scholar] [CrossRef] [Green Version]
- Robaglia, C.; Caranta, C. Translation initiation factors: A weak link in plant RNA virus infection. Trends Plant Sci. 2006, 11, 40–45. [Google Scholar] [CrossRef]
- Goodfellow, I. The genome-linked protein VPg of vertebrate viruses—A multifaceted protein. Curr. Opin. Virol. 2011, 1, 355–362. [Google Scholar] [CrossRef] [Green Version]
- Lellis, A.D.; Kasschau, K.D.; Whitham, S.A.; Carrington, J.C. Loss-of-susceptibility mutants of Arabidopsis thaliana reveal an essential role for eIF(iso)4E during potyvirus infection. Curr. Biol. 2002, 12, 1046–1051. [Google Scholar] [CrossRef] [Green Version]
- Duprat, A.; Caranta, C.; Revers, F.; Menand, B.; Browning, K.S.; Robaglia, C. The Arabidopsis eukaryotic initiation factor (iso)4E is dispensable for plant growth but required for susceptibility to potyviruses. Plant J. 2002, 32, 927–934. [Google Scholar] [CrossRef]
- Sato, M.; Nakahara, K.; Yoshii, M.; Ishikawa, M.; Uyeda, I. Selective involvement of members of the eukaryotic initiation factor 4E family in the infection of Arabidopsis thaliana by potyviruses. FEBS Lett. 2005, 579, 1167–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuta, C.; Yamana, T.; Tacahashi, Y.; Uyeda, I.; Sato, M.; Ueda, S.; Matsumura, T. Development of clover yellow vein virus as an efficient, stable gene-expression system for legume species. Plant J. 2000, 23, 539–546. [Google Scholar] [CrossRef] [Green Version]
- Ruffel, S.; Dussault, M.H.; Palloix, A.; Moury, B.; Bendahmane, A.; Robaglia, C.; Caranta, C. A natural recessive resistance gene against potato virus Y in pepper corresponds to the eukaryotic initiation factor 4E (eIF4E). Plant J. 2002, 32, 1067–1075. [Google Scholar] [CrossRef]
- Nicaise, V.; German-Retana, S.; Sanjuan, R.; Dubrana, M.P.; Mazier, M.; Maisonneuve, B.; Candresse, T.; Caranta, C.; LeGall, O. The eukaryotic translation initiation factor 4E controls lettuce susceptibility to the Potyvirus Lettuce mosaic virus. Plant Physiol. 2003, 132, 1272–1282. [Google Scholar] [CrossRef] [Green Version]
- Piron, F.; Nicolai, M.; Minoia, S.; Piednoir, E.; Moretti, A.; Salgues, A.; Zamir, D.; Caranta, C.; Bendahmane, A. An Induced Mutation in Tomato eIF4E Leads to Immunity to Two Potyviruses. PLoS ONE 2010, 5, e11313. [Google Scholar] [CrossRef] [Green Version]
- Mazier, M.; Flamain, F.; Nicolai, M.; Sarnette, V.; Caranta, C. Knock-Down of Both eIF4E1 and eIF4E2 Genes Confers Broad-Spectrum Resistance against Potyviruses in Tomato. PLoS ONE 2011, 6, e29595. [Google Scholar] [CrossRef] [PubMed]
- Maule, A.J.; Caranta, C.; Boulton, M.I. Sources of natural resistance to plant viruses: Status and prospects. Mol. Plant Pathol. 2007, 8, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Yoon, Y.J.; Venkatesh, J.; Lee, J.H.; Kim, J.; Lee, H.E.; Kim, D.S.; Kang, B.C. Genome Editing of eIF4E1 in Tomato Confers Resistance to Pepper Mottle Virus. Front. Plant Sci. 2020, 11, 1098. [Google Scholar] [CrossRef]
- Gao, L.; Luo, J.Y.; Ding, X.N.; Wang, T.; Hu, T.; Song, P.W.; Zhai, R.; Zhang, H.Y.; Zhang, K.; Li, K.; et al. Soybean RNA interference lines silenced for eIF4E show broad potyvirus resistance. Mol. Plant Pathol. 2020, 21, 303–317. [Google Scholar] [CrossRef] [PubMed]
- Sanfacon, H. Plant Translation Factors and Virus Resistance. Viruses 2015, 7, 3392–3419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swisher Grimm, K.D.; Porter, L.D. Development and Validation of KASP Markers for the Identification of Pea seedborne mosaic virus Pathotype P1 Resistance in Pisum sativum. Plant Dis. 2020, 104, 1824–1830. [Google Scholar] [CrossRef]
- Zhai, Y.S.; Deng, Y.Q.; Cheng, G.Y.; Peng, L.; Zheng, Y.R.; Yang, Y.Q.; Xu, J.S. Sugarcane Elongin C is involved in infection by sugarcane mosaic disease pathogens. Biochem. Biophys. Res. Commun. 2015, 466, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Cheng, G.; Yang, Z.; Wang, T.; Xu, J. Identification of Sugarcane Host Factors Interacting with the 6K2 Protein of the Sugarcane Mosaic Virus. Int. J. Mol. Sci. 2019, 20, 3867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.S.; Deng, Y.Q.; Cheng, G.Y.; Zhai, Y.S.; Peng, L.; Dong, M.; Xu, Q.; Yang, Y.Q. Sugarcane mosaic virus infection of model plants Brachypodium distachyon and Nicotiana benthamiana. J. Intergr. Agric. 2019, 18, 2294–2301. [Google Scholar] [CrossRef]
- Zhang, J.S.; Zhang, X.T.; Tang, H.B.; Zhang, Q.; Hua, X.T.; Ma, X.K.; Zhu, F.; Jones, T.; Zhu, X.G.; Bowers, J.; et al. Allele-defined genome of the autopolyploid sugarcane Saccharum spontaneum L. Nat. Genet. 2018, 50. [Google Scholar] [CrossRef] [Green Version]
- Iskandar, H.M.; Simpson, R.S.; Casu, R.E.; Bonnett, G.D.; Maclean, D.J.; Manners, J.M. Comparison of reference genes for quantitative real-time polymerase chain reaction analysis of gene expression in sugarcane. Plant Mol. Biol. Report. 2004, 22, 325–337. [Google Scholar] [CrossRef]
- Ling, H.; Wu, Q.; Guo, J.; Xu, L.; Que, Y. Comprehensive selection of reference genes for gene expression normalization in sugarcane by real time quantitative RT-PCR. PLoS ONE 2014, 9, e97469. [Google Scholar] [CrossRef] [Green Version]
- Joshi, B.; Lee, K.; Maeder, D.L.; Jagus, R. Phylogenetic analysis of eIF4E-family members. BMC Evol. Biol. 2005, 5, 48. [Google Scholar] [CrossRef] [Green Version]
- Kropiwnicka, A.; Kuchta, K.; Lukaszewicz, M.; Kowalska, J.; Jemielity, J.; Ginalski, K.; Darzynkiewicz, E.; Zuberek, J. Five eIF4E isoforms from Arabidopsis thaliana are characterized by distinct features of cap analogs binding. Biochem. Biophys. Res. Commun. 2015, 456, 47–52. [Google Scholar] [CrossRef]
- Gebhardt, A.; Habjan, M.; Benda, C.; Meiler, A.; Haas, D.A.; Hein, M.Y.; Mann, A.; Mann, M.; Habermann, B.; Pichlmair, A. mRNA export through an additional cap-binding complex consisting of NCBP1 and NCBP3. Nat. Commun. 2015, 6, 8192. [Google Scholar] [CrossRef] [Green Version]
- Dou, Y.; Kalmykova, S.; Pashkova, M.; Oghbaie, M.; Jiang, H.; Molloy, K.R.; Chait, B.T.; Rout, M.P.; Fenyo, D.; Jensen, T.H.; et al. Affinity proteomic dissection of the human nuclear cap-binding complex interactome. Nucleic Acids Res. 2020, 48, 10456–10469. [Google Scholar] [CrossRef] [PubMed]
- Keima, T.; Hagiwara-Komoda, Y.; Hashimoto, M.; Neriya, Y.; Koinuma, H.; Iwabuchi, N.; Nishida, S.; Yamaji, Y.; Namba, S. Deficiency of the eIF4E isoform nCBP limits the cell-to-cell movement of a plant virus encoding triple-gene-block proteins in Arabidopsis thaliana. Sci. Rep. 2017, 7, 39678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez, M.A.; Lin, Z.D.; Moll, T.; Chauhan, R.D.; Hayden, L.; Renninger, K.; Beyene, G.; Taylor, N.J.; Carrington, J.C.; Staskawicz, B.J.; et al. Simultaneous CRISPR/Cas9-mediated editing of cassava eIF4E isoforms nCBP-1 and nCBP-2 reduces cassava brown streak disease symptom severity and incidence. Plant Biotechnol. J. 2019, 17, 421–434. [Google Scholar] [CrossRef] [Green Version]
- Ingelbrecht, I.L.; Irvine, J.E.; Mirkov, T.E. Posttranscriptional gene silencing in transgenic sugarcane. Dissection of homology-dependent virus resistance in a monocot that has a complex polyploid genome. Plant Physiol 1999, 119, 1187–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.Q.; Liu, R.Y.; Zhou, T.; Fan, Z.F. Genetic diversity and population structure of Sugarcane mosaic virus. Virus Res. 2013, 171, 242–246. [Google Scholar] [CrossRef]
- Zafirov, D.; Giovinazzo, N.; Bastet, A.; Gallois, J.-L. When a knockout is an Achilles’ heel: Resistance to one potyvirus species triggers hypersusceptibility to another one in Arabidopsis thaliana. Mol. Plant Pathog. 2021, 22, 334–347. [Google Scholar] [CrossRef]
- Gauffier, C.; Lebaron, C.; Moretti, A.; Constant, C.; Moquet, F.; Bonnet, G.; Caranta, C.; Gallois, J.L. A TILLING approach to generate broad-spectrum resistance to potyviruses in tomato is hampered by eIF4E gene redundancy. Plant J. 2016, 85, 717–729. [Google Scholar] [CrossRef] [Green Version]
- Bastet, A.; Zafirov, D.; Giovinazzo, N.; Guyon-Debast, A.; Nogue, F.; Robaglia, C.; Gallois, J.L. Mimicking natural polymorphism in eIF4E by CRISPR-Cas9 base editing is associated with resistance to potyviruses. Plant Biotechnol. J. 2019, 17, 1736–1750. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.; Dong, M.; Cheng, G.; Liu, S.; Zhang, H.; Shang, H.; Zhou, Y.; Huang, G.; Zhang, M.; Wang, F.; et al. Selective Interaction of Sugarcane eIF4E with VPgs from Sugarcane Mosaic Pathogens. Viruses 2021, 13, 518. https://0-doi-org.brum.beds.ac.uk/10.3390/v13030518
Yang Z, Dong M, Cheng G, Liu S, Zhang H, Shang H, Zhou Y, Huang G, Zhang M, Wang F, et al. Selective Interaction of Sugarcane eIF4E with VPgs from Sugarcane Mosaic Pathogens. Viruses. 2021; 13(3):518. https://0-doi-org.brum.beds.ac.uk/10.3390/v13030518
Chicago/Turabian StyleYang, Zongtao, Meng Dong, Guangyuan Cheng, Shuxian Liu, Hai Zhang, Heyang Shang, Yingshuan Zhou, Guoqiang Huang, Muqing Zhang, Fengji Wang, and et al. 2021. "Selective Interaction of Sugarcane eIF4E with VPgs from Sugarcane Mosaic Pathogens" Viruses 13, no. 3: 518. https://0-doi-org.brum.beds.ac.uk/10.3390/v13030518