
N-Linked Glycosylation on Anthrax Toxin Receptor 1 Is Essential for Seneca Valley Virus Infection

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. SVV Purification
2.2. SVV-ANTXR1 Interaction
2.3. Glycopeptide Analysis of ANTXR1 Glycosylation Sites
2.4. Co-Immunoprecipitation Studies
2.5. Cell Blocking Studies
2.6. Plaque Formation Assay
2.7. Focused Classification/Refinement of SVV-ANTXR1 Asymmetric Unit and Modeling of Glycans
3. Results
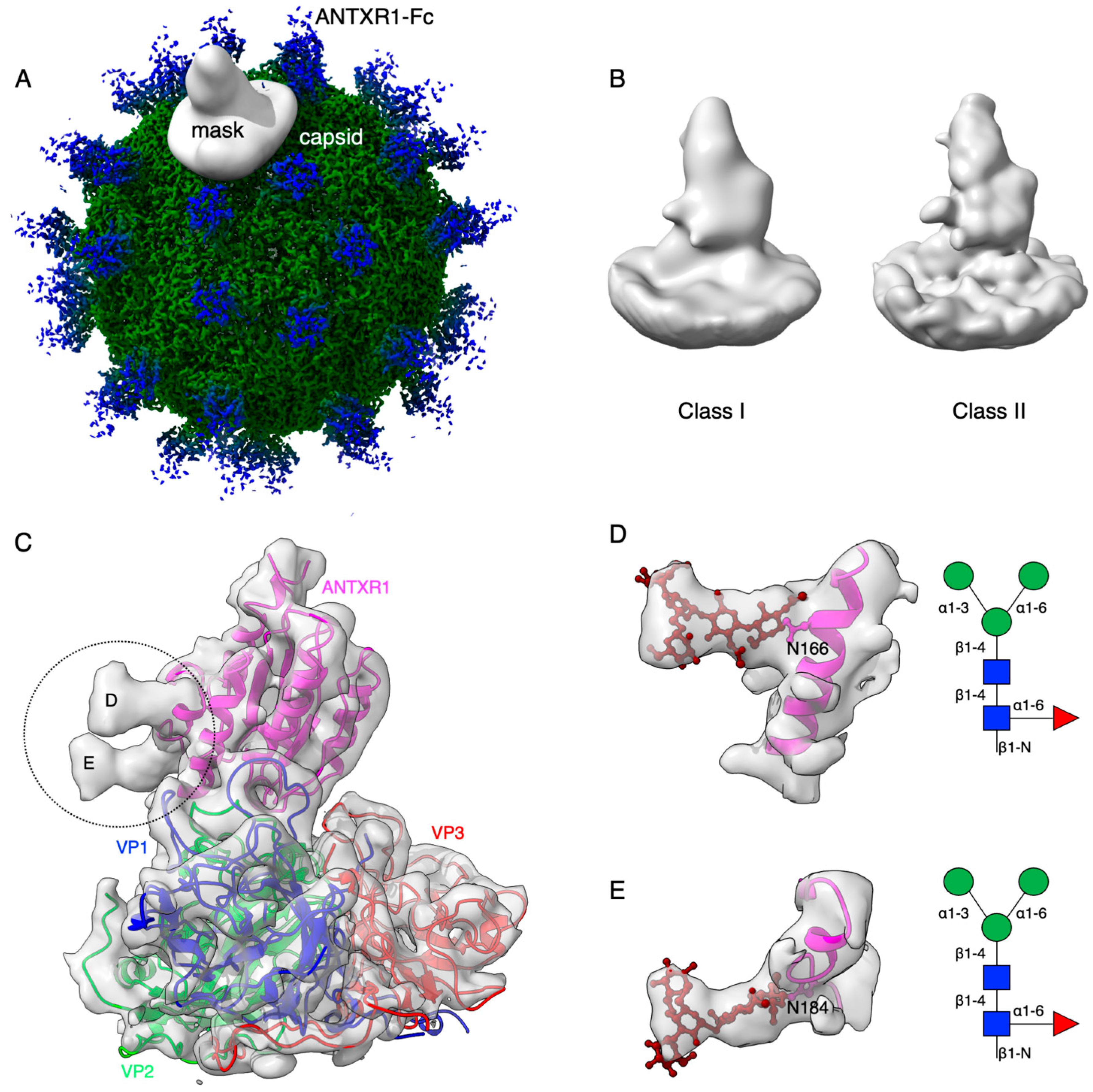
3.1. Profiling of Glycosylation Sites in ANTXR1
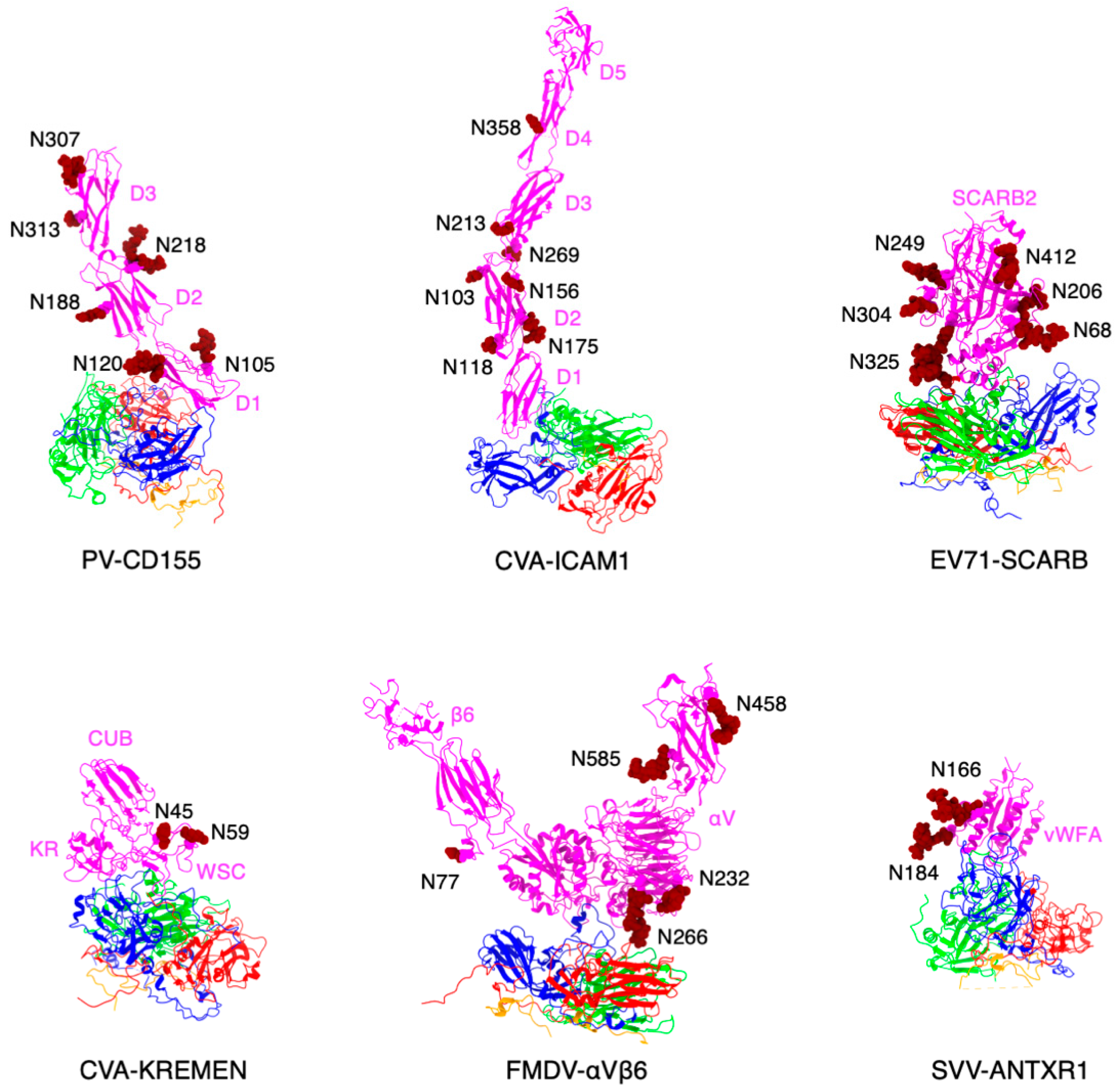
3.2. Cryo-Electron Microscopy Reconstruction of ANTXR1 Glycosylation Sites
3.3. ANTXR1 Glycosylation Is Essential for SVV Attachment and Entry
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hales, L.M.; Knowles, N.J.; Reddy, P.S.; Xu, L.; Hay, C.; Hallenbeck, P.L. Complete genome sequence analysis of Seneca Valley virus-001, a novel oncolytic picornavirus. J. Gen. Virol. 2008, 89, 1265–1275. [Google Scholar] [CrossRef] [PubMed]
- Strauss, M.; Jayawardena, N.; Sun, E.; Easingwood, R.A.; Burga, L.N.; Bostina, M. Cryo-Electron Microscopy Structure of Seneca Valley Virus Procapsid. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkataraman, S.; Reddy, S.P.; Loo, J.; Idamakanti, N.; Hallenbeck, P.L.; Reddy, V.S. Structure of Seneca Valley Virus-001: An oncolytic picornavirus representing a new genus. Structure 2008, 16, 1555–1561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Zhang, X.; Yan, R.; Yang, P.; Wu, Y.; Yang, D.; Bian, C.; Zhao, J. Emergence of a novel recombinant Seneca Valley virus in Central China, 2018. Emerg. Microbes Infect. 2018, 7, 180. [Google Scholar] [CrossRef] [Green Version]
- Burke, M.J. Oncolytic Seneca Valley Virus: Past perspectives and future directions. Oncolytic Virother. 2016, 5, 81–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudin, C.M.; Poirier, J.T.; Senzer, N.N.; Stephenson, J., Jr.; Loesch, D.; Burroughs, K.D.; Reddy, P.S.; Hann, C.L.; Hallenbeck, P.L. Phase I clinical study of Seneca Valley Virus (SVV-001), a replication-competent picornavirus, in advanced solid tumors with neuroendocrine features. Clin. Cancer Res. 2011, 17, 888–895. [Google Scholar] [CrossRef] [Green Version]
- Morton, C.L.; Houghton, P.J.; Kolb, E.A.; Gorlick, R.; Reynolds, C.P.; Kang, M.H.; Maris, J.M.; Keir, S.T.; Wu, J.; Smith, M.A. Initial testing of the replication competent Seneca Valley virus (NTX-010) by the pediatric preclinical testing program. Pediatr. Blood Cancer 2010, 55, 295–303. [Google Scholar] [CrossRef] [Green Version]
- Poirier, J.T.; Dobromilskaya, I.; Moriarty, W.F.; Peacock, C.D.; Hann, C.L.; Rudin, C.M. Selective tropism of Seneca Valley virus for variant subtype small cell lung cancer. J. Natl. Cancer Inst. 2013, 105, 1059–1065. [Google Scholar] [CrossRef] [Green Version]
- Reddy, P.S.; Burroughs, K.D.; Hales, L.M.; Ganesh, S.; Jones, B.H.; Idamakanti, N.; Hay, C.; Li, S.S.; Skele, K.L.; Vasko, A.J.; et al. Seneca Valley virus, a systemically deliverable oncolytic picornavirus, and the treatment of neuroendocrine cancers. J. Natl. Cancer Inst. 2007, 99, 1623–1633. [Google Scholar] [CrossRef]
- Burke, M.J.; Ahern, C.; Weigel, B.J.; Poirier, J.T.; Rudin, C.M.; Chen, Y.; Cripe, T.P.; Bernhardt, M.B.; Blaney, S.M. Phase I trial of Seneca Valley Virus (NTX-010) in children with relapsed/refractory solid tumors: A report of the Children’s Oncology Group. Pediatr. Blood Cancer 2015, 62, 743–750. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Zhang, J.; Wang, M.; Pan, S.; Mou, C.; Chen, Z. Pathogenicity of two Chinese Seneca Valley virus (SVV) strains in pigs. Microb. Pathog. 2019, 136, 103695. [Google Scholar] [CrossRef]
- Liu, J.; Ren, X.; Li, Z.; Xu, G.; Lu, R.; Zhang, K.; Ning, Z. Genetic and phylogenetic analysis of reemerged novel Seneca Valley virus strains in Guangdong province, 2017. Transbound. Emerg. Dis. 2018, 65, 614–617. [Google Scholar] [CrossRef]
- Liu, J.; Zha, Y.; Li, H.; Sun, Y.; Wang, F.; Lu, R.; Ning, Z. Novel Recombinant Seneca Valley Virus Isolated from Slaughtered Pigs in Guangdong Province. Virol. Sin. 2019, 34, 722–724. [Google Scholar] [CrossRef]
- Qian, S.; Fan, W.; Qian, P.; Chen, H.; Li, X. Isolation and full-genome sequencing of Seneca Valley virus in piglets from China, 2016. Virol. J. 2016, 13, 173. [Google Scholar] [CrossRef] [Green Version]
- Resende, T.P.; Marthaler, D.G.; Vannucci, F.A. A novel RNA-based in situ hybridization to detect Seneca Valley virus in neonatal piglets and sows affected with vesicular disease. PLoS ONE 2017, 12, e0173190. [Google Scholar] [CrossRef] [Green Version]
- Saporiti, V.; Fritzen, J.T.T.; Feronato, C.; Leme, R.A.; Lobato, Z.I.P.; Alfieri, A.F.; Alfieri, A.A. A ten years (2007–2016) retrospective serological survey for Seneca Valley virus infection in major pig producing states of Brazil. Vet. Res. Commun. 2017, 41, 317–321. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, H.; Sun, W.; Jiao, C.; Xiao, P.; Han, J.; Nan, F.; Xie, C.; Ha, Z.; Li, Z.; et al. Genetic evolution and epidemiological analysis of Seneca Valley virus (SVV) in China. Virus Res. 2021, 291, 198177. [Google Scholar] [CrossRef] [PubMed]
- Rossmann, M.G.; He, Y.; Kuhn, R.J. Picornavirus-receptor interactions. Trends Microbiol. 2002, 10, 324–331. [Google Scholar] [CrossRef]
- Tuthill, T.J.; Groppelli, E.; Hogle, J.M.; Rowlands, D.J. Picornaviruses. Curr. Top. Microbiol. Immunol. 2010, 343, 43–89. [Google Scholar] [CrossRef] [PubMed]
- Vidarsson, G.; Dekkers, G.; Rispens, T. IgG subclasses and allotypes: From structure to effector functions. Front. Immunol. 2014, 5, 520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Luo, Y.; Wu, Y.; Tsao, J.; Luo, M. Sialylation of the host receptor may modulate entry of demyelinating persistent Theiler’s virus. J. Virol. 2000, 74, 1477–1485. [Google Scholar] [CrossRef] [Green Version]
- Fry, E.E.; Lea, S.M.; Jackson, T.; Newman, J.W.; Ellard, F.M.; Blakemore, W.E.; Abu-Ghazaleh, R.; Samuel, A.; King, A.M.; Stuart, D.I. The structure and function of a foot-and-mouth disease virus-oligosaccharide receptor complex. EMBO J. 1999, 18, 543–554. [Google Scholar] [CrossRef] [Green Version]
- Zocher, G.; Mistry, N.; Frank, M.; Hahnlein-Schick, I.; Ekstrom, J.O.; Arnberg, N.; Stehle, T. A sialic acid binding site in a human picornavirus. PLoS. Pathog. 2014, 10, e1004401. [Google Scholar] [CrossRef] [Green Version]
- Hafenstein, S.; Bowman, V.D.; Chipman, P.R.; Bator Kelly, C.M.; Lin, F.; Medof, M.E.; Rossmann, M.G. Interaction of decay-accelerating factor with coxsackievirus B3. J. Virol. 2007, 81, 12927–12935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plevka, P.; Hafenstein, S.; Harris, K.G.; Cifuente, J.O.; Zhang, Y.; Bowman, V.D.; Chipman, P.R.; Bator, C.M.; Lin, F.; Medof, M.E.; et al. Interaction of decay-accelerating factor with echovirus 7. J. Virol. 2010, 84, 12665–12674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miles, L.A.; Burga, L.N.; Gardner, E.E.; Bostina, M.; Poirier, J.T.; Rudin, C.M. Anthrax toxin receptor 1 is the cellular receptor for Seneca Valley virus. J. Clin. Investig. 2017, 127, 2957–2967. [Google Scholar] [CrossRef] [Green Version]
- Fu, S.; Tong, X.; Cai, C.; Zhao, Y.; Wu, Y.; Li, Y.; Xu, J.; Zhang, X.C.; Xu, L.; Chen, W.; et al. The structure of tumor endothelial marker 8 (TEM8) extracellular domain and implications for its receptor function for recognizing anthrax toxin. PLoS ONE 2010, 5, e11203. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.H.; Liu, S.; Bankston, L.A.; Liddington, R.C.; Leppla, S.H. Selection of anthrax toxin protective antigen variants that discriminate between the cellular receptors TEM8 and CMG2 and achieve targeting of tumor cells. J. Biol. Chem. 2007, 282, 9834–9845. [Google Scholar] [CrossRef] [Green Version]
- Jayawardena, N.; Burga, L.N.; Easingwood, R.A.; Takizawa, Y.; Wolf, M.; Bostina, M. Structural basis for anthrax toxin receptor 1 recognition by Seneca Valley Virus. Proc. Natl. Acad. Sci. USA 2018, 115, E10934–E10940. [Google Scholar] [CrossRef] [Green Version]
- Raman, R.; Tharakaraman, K.; Sasisekharan, V.; Sasisekharan, R. Glycan-protein interactions in viral pathogenesis. Curr. Opin. Struct. Biol. 2016, 40, 153–162. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhu, Z.; Chen, W.; Cai, Z.; Xu, B.; Tan, Z.; Wu, A.; Ge, X.; Guo, X.; Tan, Z.; et al. Cell membrane proteins with high N-glycosylation, high expression and multiple interaction partners are preferred by mammalian viruses as receptors. Bioinformatics 2019, 35, 723–728. [Google Scholar] [CrossRef] [PubMed]
- Strauss, M.; Filman, D.J.; Belnap, D.M.; Cheng, N.; Noel, R.T.; Hogle, J.M. Nectin-like interactions between poliovirus and its receptor trigger conformational changes associated with cell entry. J. Virol. 2015, 89, 4143–4157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, C.; Bator-Kelly, C.M.; Rieder, E.; Chipman, P.R.; Craig, A.; Kuhn, R.J.; Wimmer, E.; Rossmann, M.G. The crystal structure of coxsackievirus A21 and its interaction with ICAM-1. Structure 2005, 13, 1019–1033. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Zhou, D.; Ni, T.; Karia, D.; Kotecha, A.; Wang, X.; Rao, Z.; Jones, E.Y.; Fry, E.E.; Ren, J.; et al. Hand-foot-and-mouth disease virus receptor KREMEN1 binds the canyon of Coxsackie Virus A10. Nat. Commun. 2020, 11, 38. [Google Scholar] [CrossRef]
- He, Y.; Chipman, P.R.; Howitt, J.; Bator, C.M.; Whitt, M.A.; Baker, T.S.; Kuhn, R.J.; Anderson, C.W.; Freimuth, P.; Rossmann, M.G. Interaction of coxsackievirus B3 with the full length coxsackievirus-adenovirus receptor. Nat. Struct. Biol. 2001, 8, 874–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, D.; Zhao, Y.; Kotecha, A.; Fry, E.E.; Kelly, J.T.; Wang, X.; Rao, Z.; Rowlands, D.J.; Ren, J.; Stuart, D.I. Unexpected mode of engagement between enterovirus 71 and its receptor SCARB2. Nat. Microbiol. 2019, 4, 414–419. [Google Scholar] [CrossRef]
- Kotecha, A.; Wang, Q.; Dong, X.; Ilca, S.L.; Ondiviela, M.; Zihe, R.; Seago, J.; Charleston, B.; Fry, E.E.; Abrescia, N.G.A.; et al. Rules of engagement between alphavbeta6 integrin and foot-and-mouth disease virus. Nat. Commun. 2017, 8, 15408. [Google Scholar] [CrossRef]
- Zibert, A.; Wimmer, E. N glycosylation of the virus binding domain is not essential for function of the human poliovirus receptor. J. Virol. 1992, 66, 7368–7373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friebe, S.; Deuquet, J.; van der Goot, F.G. Differential dependence on N-glycosylation of anthrax toxin receptors CMG2 and TEM8. PLoS ONE 2015, 10, e0119864. [Google Scholar] [CrossRef] [Green Version]
- Noble, J.E.; Knight, A.E.; Reason, A.J.; Di Matola, A.; Bailey, M.J. A comparison of protein quantitation assays for biopharmaceutical applications. Mol. Biotechnol. 2007, 37, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Poirier, J.T.; Reddy, P.S.; Idamakanti, N.; Li, S.S.; Stump, K.L.; Burroughs, K.D.; Hallenbeck, P.L.; Rudin, C.M. Characterization of a full-length infectious cDNA clone and a GFP reporter derivative of the oncolytic picornavirus SVV-001. J. Gen. Virol. 2012, 93, 2606–2613. [Google Scholar] [CrossRef] [Green Version]
- Kraus, A.A.; Messer, W.; Haymore, L.B.; de Silva, A.M. Comparison of plaque- and flow cytometry-based methods for measuring dengue virus neutralization. J. Clin. Microbiol. 2007, 45, 3777–3780. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera--a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [Green Version]
- Sun, G.; Yu, X.; Bao, C.; Wang, L.; Li, M.; Gan, J.; Qu, D.; Ma, J.; Chen, L. Identification and characterization of a novel prokaryotic peptide: N-glycosidase from Elizabethkingia meningoseptica. J. Biol. Chem. 2015, 290, 7452–7462. [Google Scholar] [CrossRef] [Green Version]
- Thobhani, S.; Yuen, C.T.; Bailey, M.J.; Jones, C. Identification and quantification of N-linked oligosaccharides released from glycoproteins: An inter-laboratory study. Glycobiology 2009, 19, 201–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, X.; Luke, B.; Andresson, T.; Blonder, J. 18O stable isotope labeling in MS-based proteomics. Brief. Funct. Genom. Proteomic 2009, 8, 136–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goh, J.B.; Ng, S.K. Impact of host cell line choice on glycan profile. Crit. Rev. Biotechnol. 2018, 38, 851–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Leiro, R.; Scheres, S.H.W. A pipeline approach to single-particle processing in RELION. Acta Crystallogr. D Struct. Biol. 2017, 73, 496–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheres, S.H. Processing of Structurally Heterogeneous Cryo-EM Data in RELION. Methods Enzymol. 2016, 579, 125–157. [Google Scholar] [CrossRef]
- Watanabe, Y.; Allen, J.D.; Wrapp, D.; McLellan, J.S.; Crispin, M. Site-specific glycan analysis of the SARS-CoV-2 spike. Science 2020, 369, 330–333. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kreutzberger, A.J.B.; Odongo, L.; Nelson, E.A.; Nyenhuis, D.A.; Kiessling, V.; Liang, B.; Cafiso, D.S.; White, J.M.; Tamm, L.K. Ebola virus glycoprotein interacts with cholesterol to enhance membrane fusion and cell entry. Nat. Struct. Mol. Biol. 2021, 28, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Wilson, I.A.; Skehel, J.J.; Wiley, D.C. Structure of the haemagglutinin membrane glycoprotein of influenza virus at 3 A resolution. Nature 1981, 289, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Rouvinski, A.; Dejnirattisai, W.; Guardado-Calvo, P.; Vaney, M.C.; Sharma, A.; Duquerroy, S.; Supasa, P.; Wongwiwat, W.; Haouz, A.; Barba-Spaeth, G.; et al. Covalently linked dengue virus envelope glycoprotein dimers reduce exposure of the immunodominant fusion loop epitope. Nat. Commun. 2017, 8, 15411. [Google Scholar] [CrossRef]
- Fontes-Garfias, C.R.; Shan, C.; Luo, H.; Muruato, A.E.; Medeiros, D.B.A.; Mays, E.; Xie, X.; Zou, J.; Roundy, C.M.; Wakamiya, M.; et al. Functional Analysis of Glycosylation of Zika Virus Envelope Protein. Cell Rep. 2017, 21, 1180–1190. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, Y.; Raghwani, J.; Allen, J.D.; Seabright, G.E.; Li, S.; Moser, F.; Huiskonen, J.T.; Strecker, T.; Bowden, T.A.; Crispin, M. Structure of the Lassa virus glycan shield provides a model for immunological resistance. Proc. Natl. Acad. Sci. USA 2018, 115, 7320–7325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vigerust, D.J.; Shepherd, V.L. Virus glycosylation: Role in virulence and immune interactions. Trends Microbiol. 2007, 15, 211–218. [Google Scholar] [CrossRef]
- Carbaugh, D.L.; Lazear, H.M. Flavivirus Envelope Protein Glycosylation: Impacts on Viral Infection and Pathogenesis. J. Virol. 2020, 94. [Google Scholar] [CrossRef] [Green Version]
- Arnberg, N.; Pring-Akerblom, P.; Wadell, G. Adenovirus type 37 uses sialic acid as a cellular receptor on Chang C cells. J. Virol. 2002, 76, 8834–8841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delorme, C.; Brussow, H.; Sidoti, J.; Roche, N.; Karlsson, K.A.; Neeser, J.R.; Teneberg, S. Glycosphingolipid binding specificities of rotavirus: Identification of a sialic acid-binding epitope. J. Virol. 2001, 75, 2276–2287. [Google Scholar] [CrossRef] [Green Version]
- Connolly, J.L.; Barton, E.S.; Dermody, T.S. Reovirus binding to cell surface sialic acid potentiates virus-induced apoptosis. J. Virol. 2001, 75, 4029–4039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, E.C.; Jamshidi, F.; Johansson, S.M.; Oberste, M.S.; Arnberg, N. Sialic acid is a cellular receptor for coxsackievirus A24 variant, an emerging virus with pandemic potential. J. Virol. 2008, 82, 3061–3068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, X.; Thinn, A.M.M.; Wang, Z.; Shan, H.; Zhu, J. The importance of N-glycosylation on beta3 integrin ligand binding and conformational regulation. Sci. Rep. 2017, 7, 4656. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Profiled Glycan | N166 | N184 | N81 (Fc) | |
|---|---|---|---|---|
| Deoxyhex1HexNAc5Hex4 |  | ✓ | ✓ | |
| Deoxyhex2HexNAc4Hex5 |  | ✓ | ✓ | |
| Deoxyhex2HexNAc5Hex4 |  | ✓ | ✓ | |
| Deoxyhex1HexNAc5Hex5 |  | ✓ | ✓ | ✓ |
| Deoxyhex1HexNAc4NeuAc1Hex5 |  | ✓ | ✓ | ✓ |
| Deoxyhex1HexNAc5NeuAc1Hex4 |  | ✓ | ✓ | |
| Deoxyhex1HexNAc5NeuAc1Sulph1Hex4 |  | ✓ | ||
| Deoxyhex2HexNAc4NeuAc1Hex5 |  | ✓ | ||
| Deoxyhex2HexNAc5NeuAc1Hex4 |  | ✓ | ||
| Deoxyhex1HexNAc5NeuAc1Hex5 |  | ✓ | ✓ | |
| Deoxyhex1HexNAc4NeuAc2Hex5 |  | ✓ | ✓ | |
| Deoxyhex1HexNAc5NeuAc2Hex4 |  | ✓ | ||
| Deoxyhex1HexNAc5NeuAc2Hex5 |  | ✓ | ||
| Deoxyhex1HexNAc6NeuAc1Hex6 |  | ✓ | ||
| Deoxyhex1HexNAc5NeuAc2Hex6 |  | ✓ | ✓ | |
| Deoxyhex1HexNAc6NeuAc2Hex6 |  | ✓ | ✓ | |
| Deoxyhex1HexNAc5NeuAc3Hex6 |  | ✓ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jayawardena, N.; Miles, L.A.; Burga, L.N.; Rudin, C.; Wolf, M.; Poirier, J.T.; Bostina, M. N-Linked Glycosylation on Anthrax Toxin Receptor 1 Is Essential for Seneca Valley Virus Infection. Viruses 2021, 13, 769. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050769
Jayawardena N, Miles LA, Burga LN, Rudin C, Wolf M, Poirier JT, Bostina M. N-Linked Glycosylation on Anthrax Toxin Receptor 1 Is Essential for Seneca Valley Virus Infection. Viruses. 2021; 13(5):769. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050769
Chicago/Turabian StyleJayawardena, Nadishka, Linde A. Miles, Laura N. Burga, Charles Rudin, Matthias Wolf, John T. Poirier, and Mihnea Bostina. 2021. "N-Linked Glycosylation on Anthrax Toxin Receptor 1 Is Essential for Seneca Valley Virus Infection" Viruses 13, no. 5: 769. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050769