
Neutralising Antibodies against Enterovirus and Parechovirus in IVIG Reflect General Circulation: A Tool for Sero-Surveillance

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. IVIG Selection
2.2. Selection of NPEV and PeV strains
2.3. Cell Line Selection
2.4. Dilution Neutralisation Assay
2.5. Statistical Analysis
3. Results
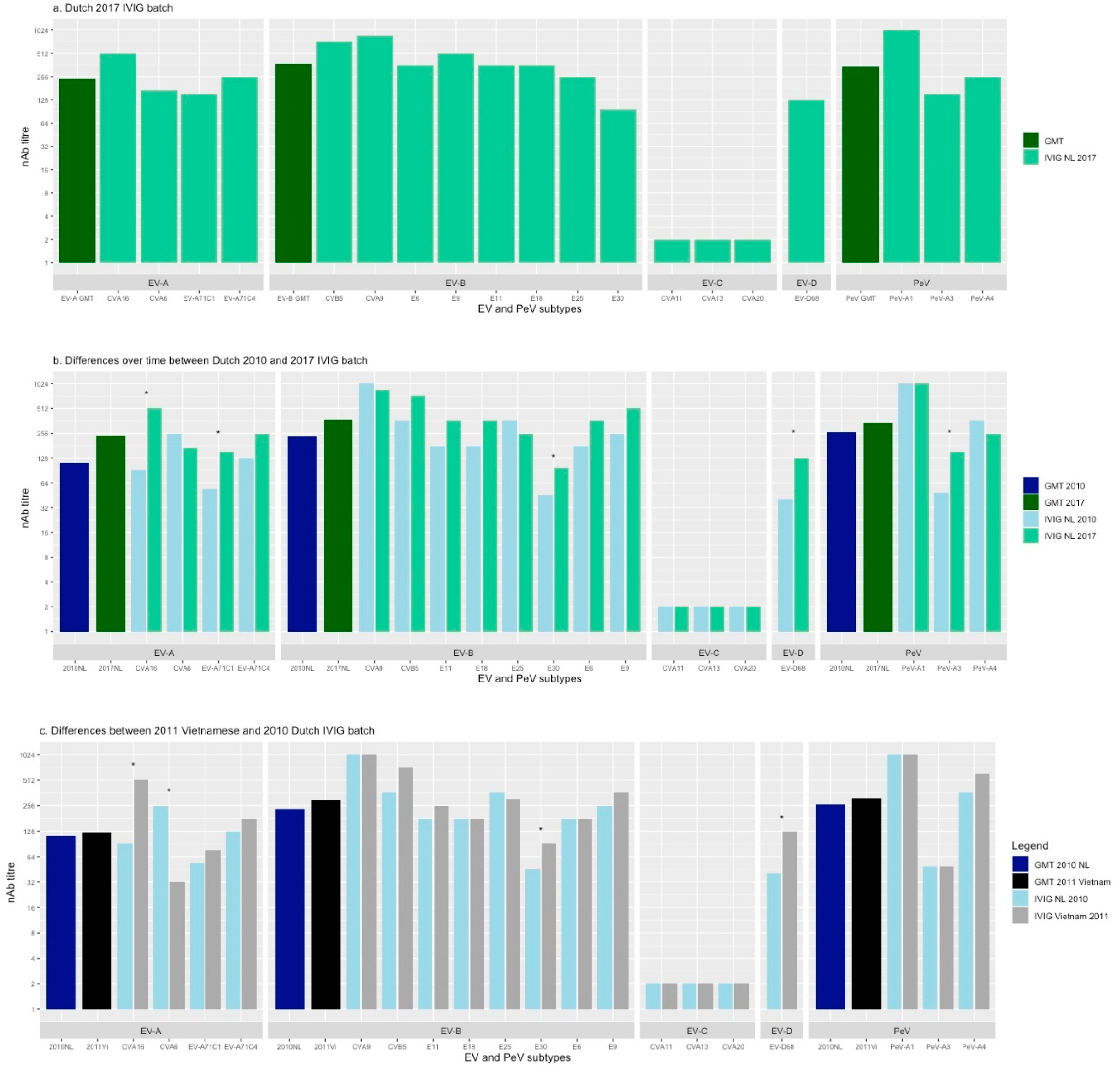
3.1. Neutralising Ab Titres Against EV and PeV in Dutch 2017 IVIG
3.2. Neutralising Ab Titres over Time in Dutch IVIG
3.3. Neutralising Ab Titres in 2010 Dutch and 2011 Vietnamese IVIG batch
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anastasina, M.; Domanska, A.; Palm, K.; Butcher, S. Human picornaviruses associated with neurological diseases and their neutralization by antibodies. J. Gen. Virol. 2017, 98, 1145–1158. [Google Scholar] [CrossRef] [PubMed]
- Pons-Salort, M.; Parker, E.P.K.; Grassly, N.C. The epidemiology of non-polio enteroviruses: Recent Advances and Outstanding Questions. Curr. Opin. Infect. Dis. 2015, 28, 479–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Crom, S.C.M.; Rossen, J.W.A.; Van Furth, A.M.; Obihara, C.C. Enterovirus and parechovirus infection in children: A brief overview. Eur. J. Nucl. Med. Mol. Imaging 2016, 175, 1023–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rantakallio, P.; Leskinen, M.; Von Wendt, L. Incidence and Prognosis of Central Nervous System Infections in a Birth Cohort of 12,000 Children. Scand. J. Infect. Dis. 1986, 18, 287–294. [Google Scholar] [CrossRef]
- Vollbach, S.; Müller, A.; Drexler, J.F.; Simon, A.; Drosten, C.; Eis-Hübinger, A.M.; Panning, M. Prevalence, type and concentration of human enterovirus and parechovirus in cerebrospinal fluid samples of pediatric patients over a 10-year period: A retrospective study. Virol. J. 2015, 12, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Van Der Sanden, S.M.; Koen, G.; Van Eijk, H.; Koekkoek, S.M.; De Jong, M.D.; Wolthers, K.C. Prediction of Protection against Asian Enterovirus 71 Outbreak Strains by Cross-neutralizing Capacity of Serum from Dutch Donors, The Netherlands. Emerg. Infect. Dis. 2016, 22, 1562–1569. [Google Scholar] [CrossRef] [Green Version]
- Pons-Salort, M.; Grassly, N.C. Serotype-specific immunity explains the incidence of diseases caused by human enteroviruses. Science 2018, 361, 800–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galama, J.M.; Vogels, M.T.; Jansen, G.H.; Gielen, M.; Heessen, F.W. Antibodies against enteroviruses in intravenous Ig preparations: Great variation in titres and poor correlation with the incidence of circulating serotypes. J. Med. Virol. 1997, 53, 273–276. [Google Scholar] [CrossRef] [Green Version]
- Wildenbeest, J.G.; Broek, P.J.V.D.; Benschop, K.S.; Koen, G.; Wierenga, P.C.; Vossen, A.C.; Kuijpers, T.W.; Wolthers, K.C. Pleconaril revisited: Clinical course of chronic enteroviral meningoencephalitis after treatment correlates with in vitro susceptibility. Antivir. Ther. 2011, 17, 459–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benschop, K.S.M.; Rahamat-Langendoen, J.C.; Van Der Avoort, H.G.A.M.; Claas, E.C.J.; Pas, S.D.; Schuurman, R.; Verweij, J.J.; Wolthers, K.C.; Niesters, H.G.M.; Koopmans, M.P.G.; et al. VIRO-TypeNed, systematic molecular surveillance of enteroviruses in the Netherlands between 2010 and 2014. Eurosurveillance 2016, 21. [Google Scholar] [CrossRef] [Green Version]
- Kapsenberg, J.G.; Ras, A.; Korte, J. Improvement of Enterovirus Neutralization by Treatment with Sodium Deoxycholate or Chloroform. Intervirology 1979, 12, 329–334. [Google Scholar] [CrossRef]
- Karelehto, E.; Van Der Sanden, S.; Geraets, J.A.; Domanska, A.; Van Der Linden, L.; Hoogendoorn, D.; Koen, G.; Van Eijk, H.; Shakeel, S.; Beaumont, T.; et al. Strain-dependent neutralization reveals antigenic variation of human parechovirus 3. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Karelehto, E.; Brouwer, L.; Benschop, K.; Kok, J.; Basile, K.; McMullan, B.; Rawlinson, W.; Druce, J.; Nicholson, S.; Selvarangan, R.; et al. Seroepidemiology of Parechovirus A3 Neutralizing Antibodies, Australia, the Netherlands, and United States. Emerg. Infect. Dis. 2019, 25, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.; Cheng, T.; Yin, Z.; Liu, D.; Xu, L.; Li, Y.; Wang, W.; Liu, J.; Que, Y.; Ye, X.; et al. Serological survey of neutralizing antibodies to eight major enteroviruses among healthy population. Emerg. Microbes Infect. 2018, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Van Der Sanden, S.M.G.; Koopmans, M.P.G.; Van Der Avoort, H.G.A.M. Detection of human enteroviruses and parechoviruses as part of the national enterovirus surveillance in the Netherlands, 1996–2011. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 1525–1531. [Google Scholar] [CrossRef] [PubMed]
- Janes, V.A.; Minnaar, R.; Koen, G.; Van Eijk, H.; Haan, K.D.-D.; Pajkrt, D.; Wolthers, K.C.; Benschop, K.S. Presence of human non-polio enterovirus and parechovirus genotypes in an Amsterdam hospital in 2007 to 2011 compared to national and international published surveillance data: A comprehensive review. Eurosurveillance 2014, 19. [Google Scholar] [CrossRef] [PubMed]
- Meijer, A.; van der Sanden, S.; Snijders, B.E.; Jaramillo-Gutierrez, G.; Bont, L.; van der Ent, C.K.; Overduin, P.; Jenny, S.L.; Jusic, E.; van der Avoort, H.G.; et al. Emergence and epidemic occurrence of enterovirus 68 respiratory infections in The Netherlands in 2010. Virology 2012, 423, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Karelehto, E.; Koen, G.; Benschop, K.; Van Der Klis, F.; Pajkrt, D.; Wolthers, K. Enterovirus D68 serosurvey: Evidence for endemic circulation in the Netherlands, 2006 to 2016. Eurosurveillance 2019, 24, 1800671. [Google Scholar] [CrossRef] [Green Version]
- Kamau, E.; Harvala, H.; Blomqvist, S.; Nguyen, D.; Horby, P.; Pebody, R.; Simmonds, P. Increase in Enterovirus D68 Infections in Young Children, United Kingdom, 2006–2016. Emerg. Infect. Dis. 2019, 25, 1200–1203. [Google Scholar] [CrossRef]
- Takamatsu, Y.; Uchida, L.; Nga, P.T.; Okamoto, K.; Nabeshima, T.; Thao, D.T.; Hai do, T.; Tuyet, N.T.; Duc, H.M.; Luat le, X.; et al. An approach for differentiating echovirus 30 and Japanese encephalitis virus infections in acute meningitis/encephalitis: A retrospective study of 103 cases in Vietnam. Virol. J. 2013, 10, 280. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Seo, Y.; Choi, U.Y.; Kim, J.-H.; Kang, J.H. Seroepidemiology of echovirus 30 in Korean children. World J. Pediatr. 2017, 13, 611–614. [Google Scholar] [CrossRef] [PubMed]
- Benschop, K.S.; Broberg, E.K.; Hodcroft, E.; Schmitz, D.; Albert, J.; Baicus, A.; Bailly, J.-L.; Baldvinsdottir, G.; Berginc, N.; Blomqvist, S.; et al. Molecular Epidemiology and Evolutionary Trajectory of Emerging Echovirus 30, Europe. Emerg. Infect. Dis. 2021, 27, 1616–1626. [Google Scholar] [CrossRef] [PubMed]
- Westerhuis, B.; Kolehmainen, P.; Benschop, K.; Nurminen, N.; Koen, G.; Koskiniemi, M.; Simell, O.; Knip, M.; Hyoty, H.; Wolthers, K.; et al. Human parechovirus seroprevalence in Finland and the Netherlands. J. Clin. Virol. 2013, 58, 211–215. [Google Scholar] [CrossRef]
- Aizawa, Y.; Suzuki, Y.; Watanabe, K.; Oishi, T.; Saitoh, A. Clinical utility of serum samples for human parechovirus type 3 infection in neonates and young infants: The 2014 epidemic in Japan. J. Infect. 2016, 72, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Imamura, T.; Okamoto, M.; Nakakita, S.-I.; Suzuki, A.; Saito, M.; Tamaki, R.; Lupisan, S.; Roy, C.N.; Hiramatsu, H.; Sugawara, K.-E.; et al. Antigenic and Receptor Binding Properties of Enterovirus 68. J. Virol. 2014, 88, 2374–2384. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Ranking | 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 |
|---|---|---|---|---|---|---|---|---|
| 1 | CVA9 | E-25 | E-18 | CVB3 | E-16 | E-11 | E-30 | E-5 |
| 2 | EV-A71 | E-7 | CVA6 | CVA9 | E-25 | CVA6 | EV-D68 | CVB5 |
| 3 | PeV-A1 | PeV-A1 | PeV-A1 | E-30 | CVA6 | E-9 | E-6 | CVA6 |
| 4 | E-30 | CVB3 | PeV-A3 | EV-A71 | CVA16 | E-18 | CVB5 | E-25 |
| 5 | PeV-A3 | CVB4 | E-9 | PeV-A1 | EV-D68 | CVB5 | PeV-A3 | E-9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Couderé, K.; van der Straten, K.; Brouwer, L.; Koen, G.; van Eijk, H.; Pajkrt, D.; Murk, J.-L.; Wolthers, K.C. Neutralising Antibodies against Enterovirus and Parechovirus in IVIG Reflect General Circulation: A Tool for Sero-Surveillance. Viruses 2021, 13, 1028. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061028
Couderé K, van der Straten K, Brouwer L, Koen G, van Eijk H, Pajkrt D, Murk J-L, Wolthers KC. Neutralising Antibodies against Enterovirus and Parechovirus in IVIG Reflect General Circulation: A Tool for Sero-Surveillance. Viruses. 2021; 13(6):1028. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061028
Chicago/Turabian StyleCouderé, Karen, Karlijn van der Straten, Lieke Brouwer, Gerrit Koen, Hetty van Eijk, Dasja Pajkrt, Jean-Luc Murk, and Katja C. Wolthers. 2021. "Neutralising Antibodies against Enterovirus and Parechovirus in IVIG Reflect General Circulation: A Tool for Sero-Surveillance" Viruses 13, no. 6: 1028. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061028