
Regulation of Krüppel-Like Factor 15 Expression by Herpes Simplex Virus Type 1 or Bovine Herpesvirus 1 Productive Infection



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Plasmids
2.3. SDS-Polyacrylamide Gels and Western Blots
2.4. Nuclear and Cytoplasmic Fractionation
2.5. Immunofluorescence
2.6. Transfection and Dual-Luciferase Reporter Assays
2.7. Analysis of KLF15 and Scrambled siRNA on Cell Viability and Productive Infection
3. Results
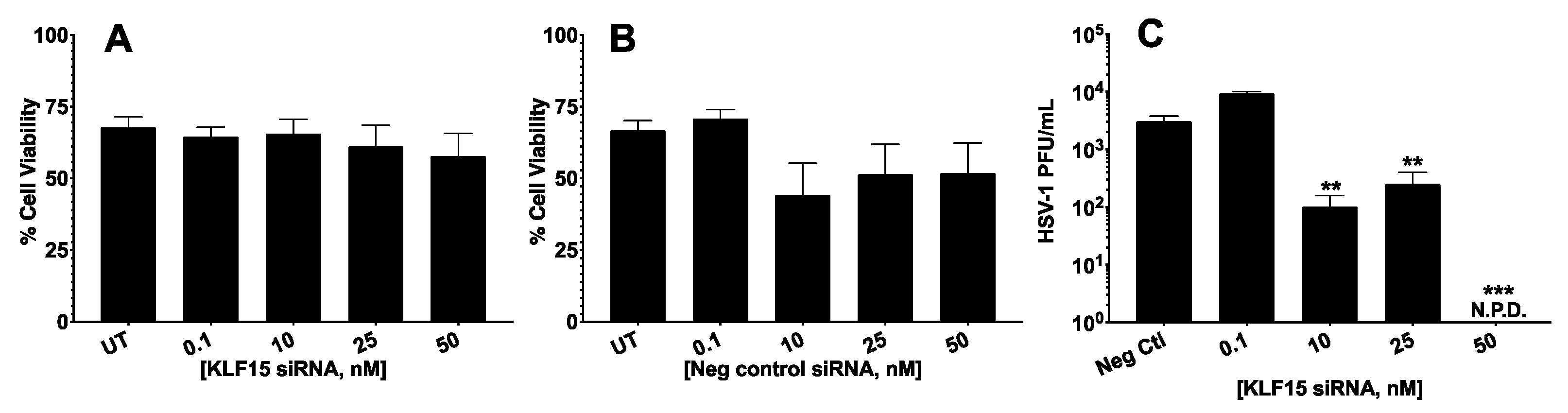
3.1. Silencing KLF15 Significantly Reduced HSV-1 Productive Infection
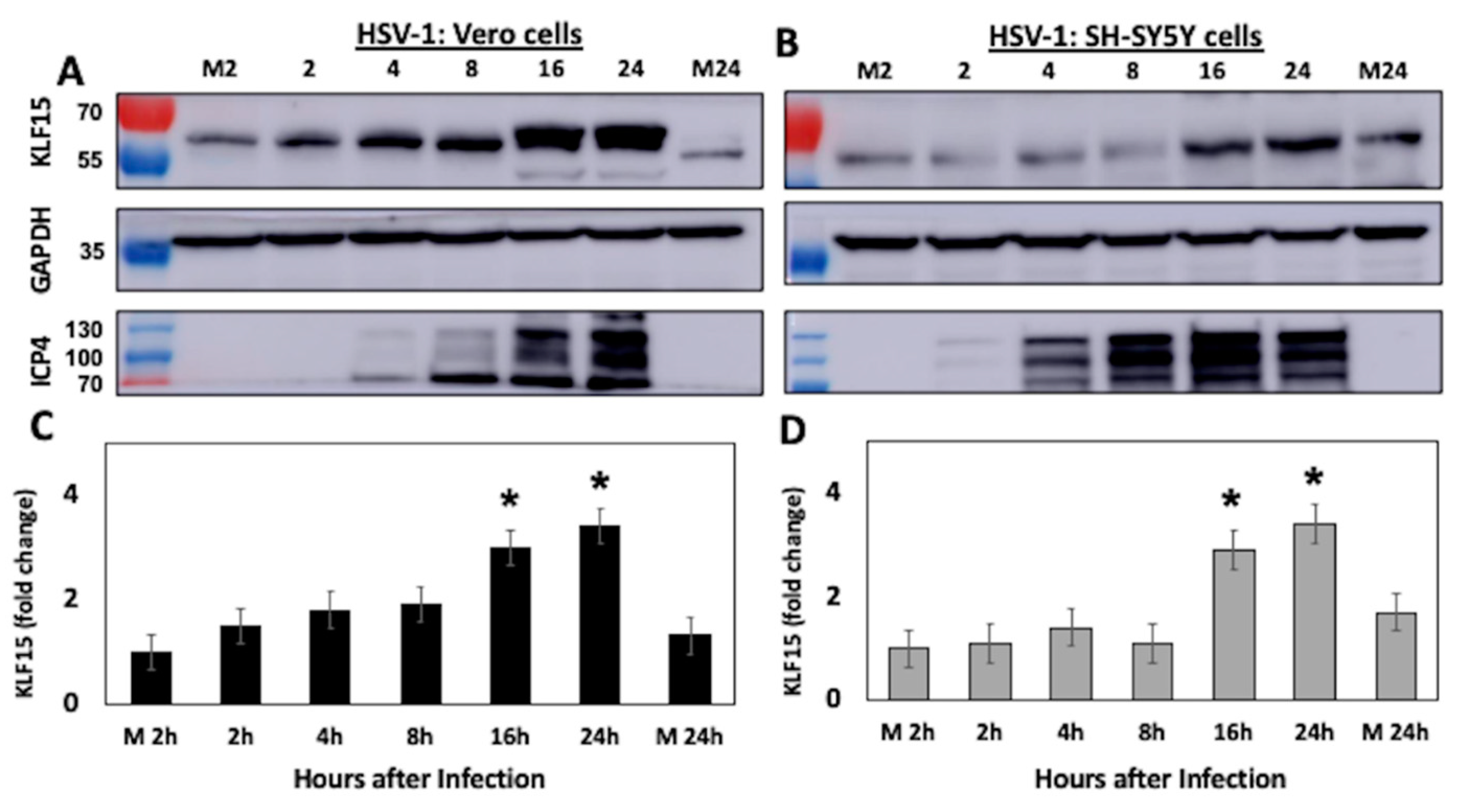
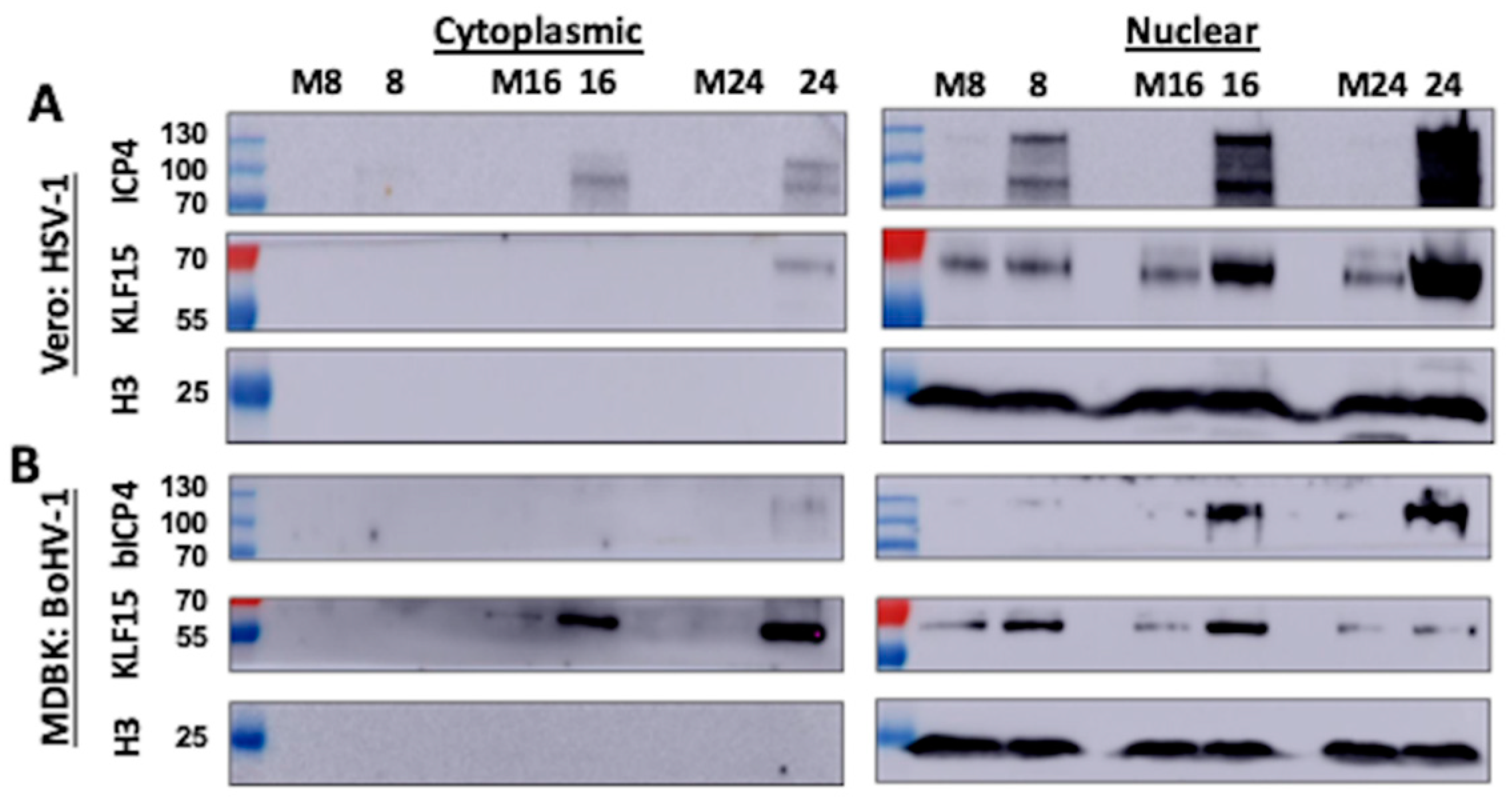
3.2. KLF15 Steady-State Protein Levels Increase Following HSV-1 Infection
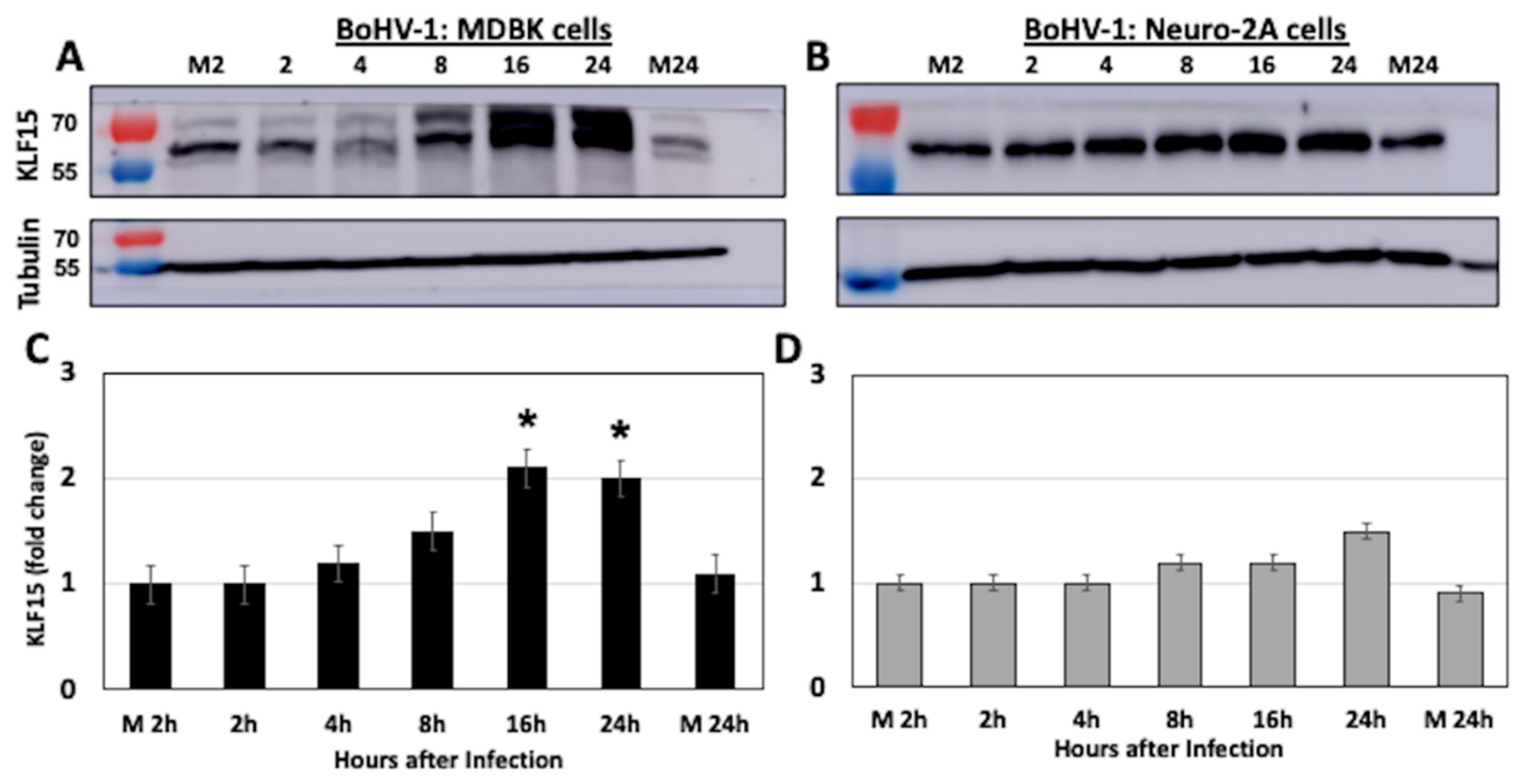
3.3. KLF15 Steady-State Protein Levels Increase Following BoHV-1 Infection
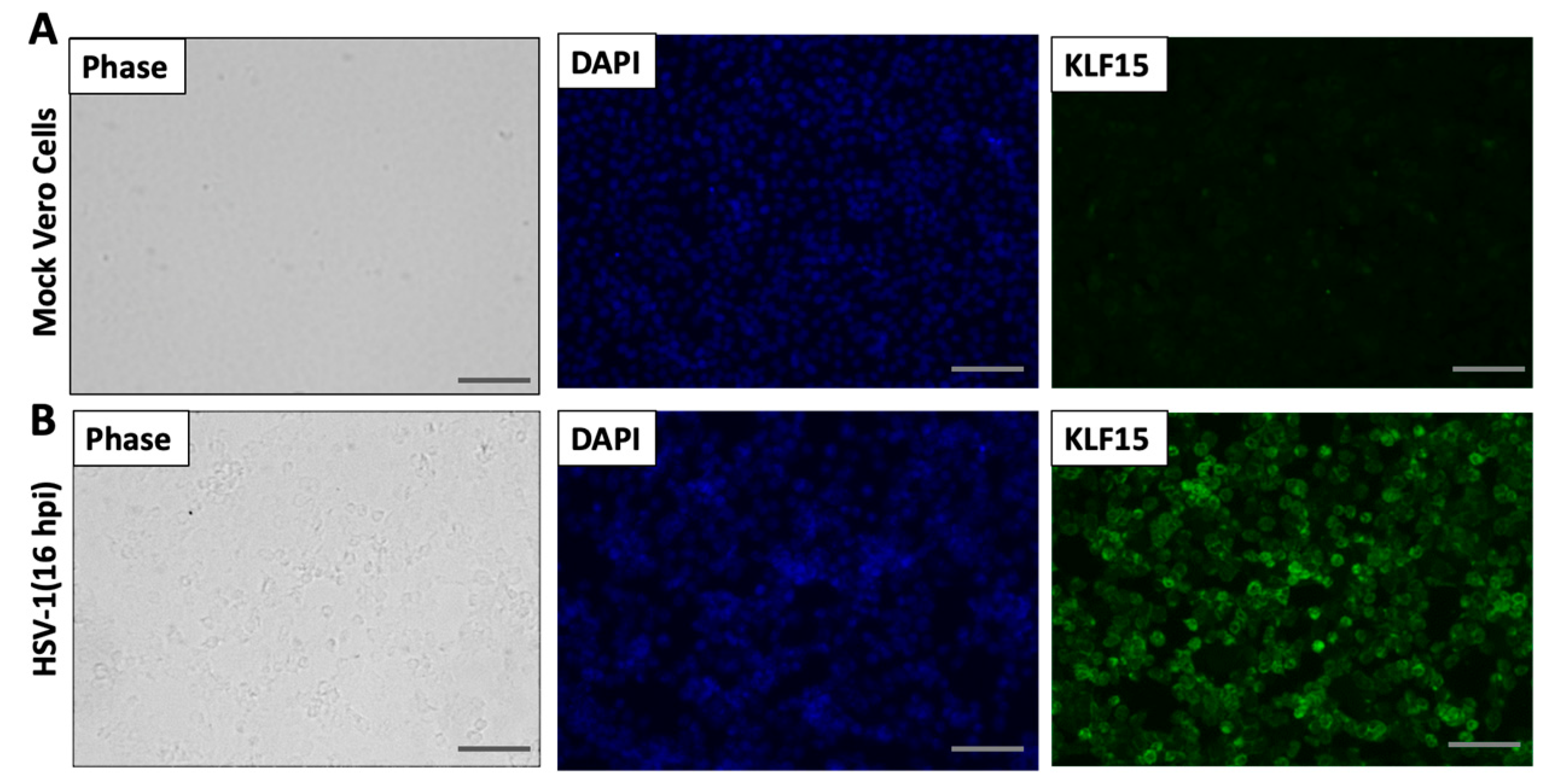
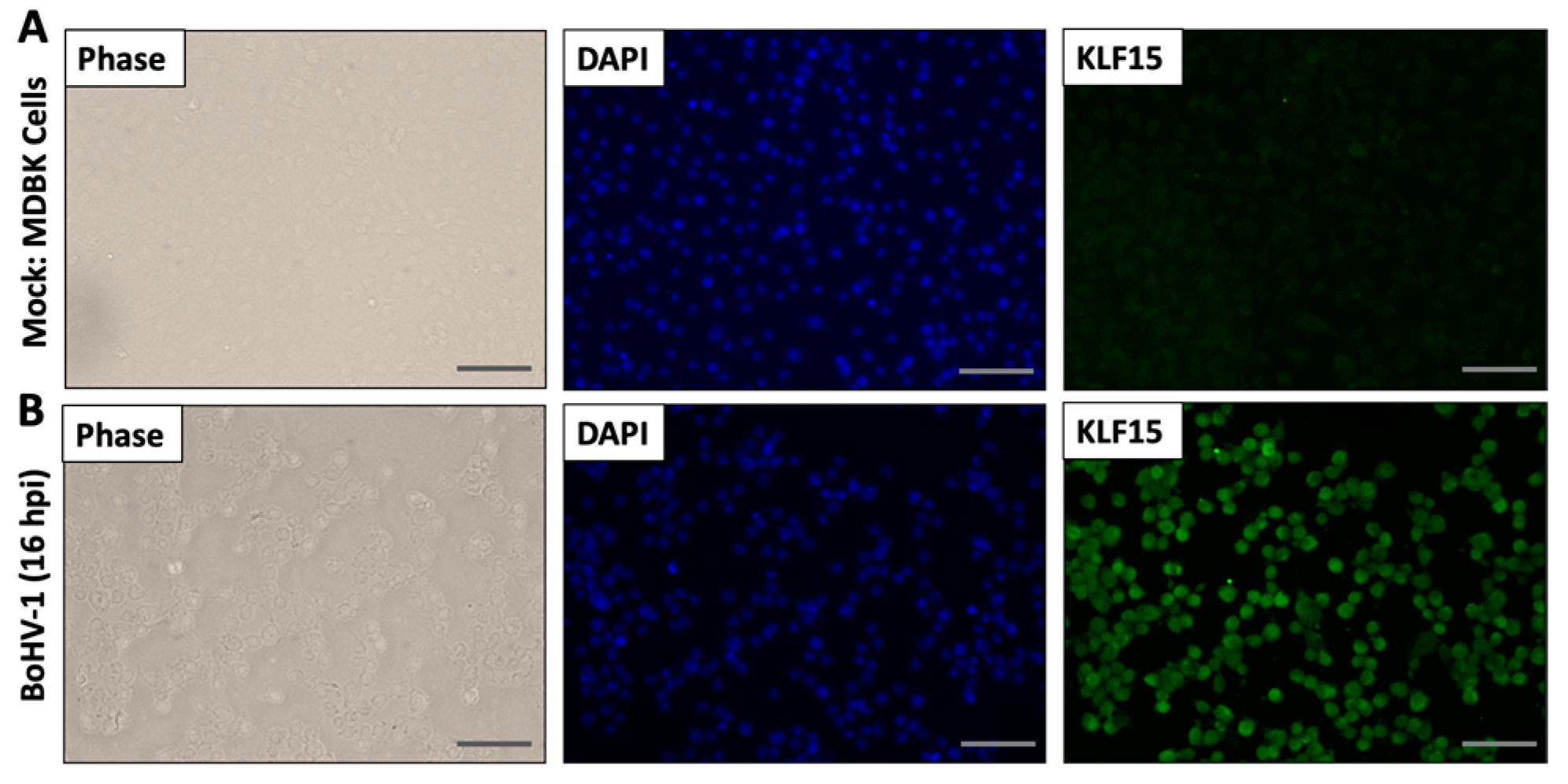
3.4. Analysis of KLF15 Localization Following Infection
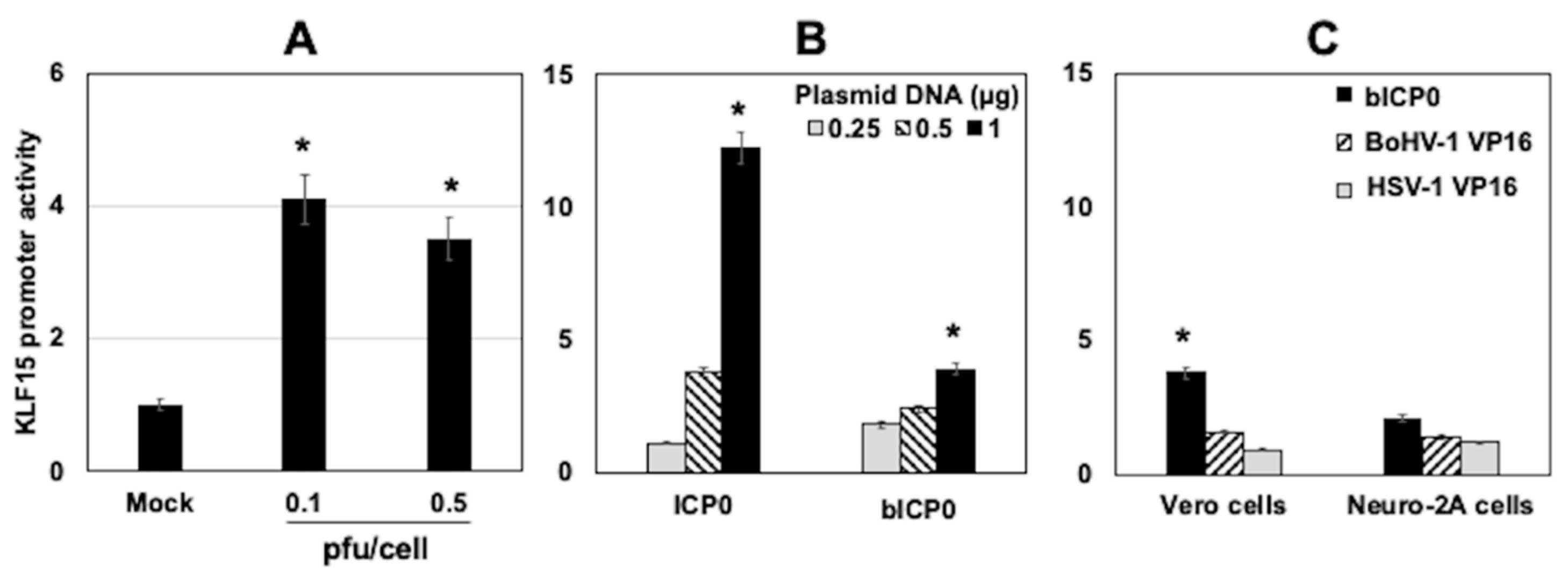
3.5. KLF15 Promoter Activity Is Stimulated by HSV-1 Infection and ICP0
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Jones, C. Alphaherpesvirus Latency: Its Role in Disease and Survival of the Virus in Nature. Adv. Viruses Res. 1998, 51, 81–133. [Google Scholar] [CrossRef]
- Honess, R.W.; Roizman, B. Regulation of herpes virus macromalecular synthesis: Cascade regulation of three groups of viral proteins. J. Virol. 1974, 14, 8–19. [Google Scholar] [CrossRef] [Green Version]
- Honess, R.W.; Roizman, B. Regulation of herpesvirus macromolecular synthesis: Sequential transition of polypeptide synthesis requires functional viral polypeptides. Proc. Natl. Acad. Sci. USA 1975, 72, 1276–1280. [Google Scholar] [CrossRef] [Green Version]
- Misra, V.; Walker, S.; Hayes, S.; O’Hare, P. The bovine herpesvirus alpha gene trans-inducing factor activates transcription by mechanisms different from those of its herpes simplex virus type 1 counterpart VP16. J. Virol. 1995, 69, 5209–5216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kristie, T.M. Dynamic modulation of HSV chromatin drives initiation of infection and proides targets for epigenetic therapies. Virology 2015, 479–480, 555–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Hare, P. The virion transactivator of herpes simplex virus. Semin. Virol. 1993, 4, 145–155. [Google Scholar] [CrossRef]
- Wirth, U.V.; Gunkel, K.; Engels, M.; Schwyzer, M. Spatial and temporal distribution of bovine herpesvirus 1 transcripts. J. Virol. 1989, 63, 4882–4889. [Google Scholar] [CrossRef] [Green Version]
- Wirth, U.V.; Vogt, B.; Schwyzer, M. The three major immediate-early transcripts of bovine herpesvirus 1 arise from two divergent and spliced transcription units. J. Virol. 1991, 65, 195–205. [Google Scholar] [CrossRef] [Green Version]
- Wirth, U.V.; Fraefel, C.; Vogt, B.; Vlcek, C.; Paces, V.; Schwyzer, M. Immediate-early RNA 2.9 and early RNA 2.6 of bovine herpesvirus 1 are 3’ coterminal and encode a putative zinc finger transactivator protein. J. Virol. 1992, 66, 2763–2772. [Google Scholar] [CrossRef] [Green Version]
- Jones, C. Herpes Simplex Virus Type 1 and Bovine Herpesvirus 1 Latency. Clin. Microbiol. Rev. 2003, 16, 79–95. [Google Scholar] [CrossRef] [Green Version]
- Jones, C. Bovine Herpes Virus 1 (BHV-1) and Herpes Simplex Virus Type 1 (HSV-1) Promote Survival of Latently Infected Sensory Neurons, in Part by Inhibiting Apoptosis. J. Cell Death 2013, 6, JCD.S10803-16. [Google Scholar] [CrossRef] [PubMed]
- Perng, G.-C.; Jones, C. Towards an Understanding of the Herpes Simplex Virus Type 1 Latency-Reactivation Cycle. Interdiscip. Perspect. Infect. Dis. 2010, 2010, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, W.; Jones, C. Open reading frame 2, encoded by the latency-related gene of bovine herpesvirus 1, has antiapoptotic activity in transiently transfected neuroblastoma cells. J. Virol. 2008, 82, 10940–10945. [Google Scholar] [CrossRef] [Green Version]
- Perng, G.-C.; Jones, C.; Ciacci-Zanella, J.; Stone, M.; Henderson, G.; Yukht, A.; Slanina, A.J.; Hoffman, F.M.; Ghiasi, H.; Nesburn, A.B.; et al. Virus-induced neuronal apoptosis blocked by the herpes simplex virus latency-associated transcript (LAT). Science 2000, 287, 1500–1503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciacci-Zanella, J.; Stone, M.; Henderson, G.; Jones, C. The latency-related gene of bovine herpesvirus 1 inhibits programmed cell death. J. Virol. 1999, 73, 9734–9740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inman, M.; Perng, G.-C.; Henderson, G.; Ghiasi, H.; Nesburn, A.B.; Wechsler, S.L.; Jones, C. Region of herpes simplex virus Type 1 latency-associated transcript sufficient for wild-type spontaneous reactivation promotes cell survival in tissue culture. J. Virol. 2001, 75, 3636–3646. [Google Scholar] [CrossRef] [Green Version]
- Phelan, D.; Barrozo, E.R.; Bloom, D.C. HSV1 latent transcription and non-coding RNA: A critical retrospective. J. Neuroimmunol. 2017, 308, 65–101. [Google Scholar] [CrossRef]
- Jones, C. Reactivation from latency by alpha-herpesvirinae submfamily members: A stressful situation. Curr. Top. Virol. 2014, 12, 99–118. [Google Scholar]
- Glaser, R.; Kiecolt-Glaser, J.K.; Speicher, C.E.; Holliday, J.E. Stress, loneliness, and changes in herpesvirus latency. J. Behav. Med. 1985, 8, 249–260. [Google Scholar] [CrossRef]
- Padgett, D.A.; Sheridan, J.; Dorne, J.; Berntson, G.G.; Candelora, J.; Glaser, R. Social stress and the reactivation of latent herpes simplex virus type 1. Proc. Natl. Acad. Sci. USA 1998, 95, 7231–7235. [Google Scholar] [CrossRef] [Green Version]
- Jones, C. Latency of Bovine Herpesvirus 1 (BoHV-1) in Sensory Neurons. In Herpesviridae; Omgradi, J., Ed.; IntechOpen: London, UK, 2016. [Google Scholar]
- Du, T.; Zhou, G.; Roizman, B. Induction of apoptosis accelerates reactivation of latent HSV-1 in ganglionic organ cultures and replication in cell cultures. Proc. Natl. Acad. Sci. USA 2012, 109, 14616–14621. [Google Scholar] [CrossRef] [Green Version]
- Harrison, K.; Zhu, L.; Thunuguntla, P.; Jones, C. Antagonizing the glucocorticoid receptor impairs explant-induced reactivation in mice latently infected with herpes simplex virus 1. J. Virol. 2019, 93, e00418-19. [Google Scholar] [CrossRef] [Green Version]
- Kook, I.; Henley, C.; Meyer, F.; Hoffmann, F.G.; Jones, C. Bovine herpesvirus 1 productive infection and immediate early transcription unit 1 promoter are stimulated by the synthetic corticosteroid dexamethasone. Virology 2015, 484, 377–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Mayet, F.S.; Sawant, L.; Thunuguntla, P.; Jones, C. Combinatorial Effects of the Glucocorticoid Receptor and Krüppel-Like Transcription Factor 15 on Bovine Herpesvirus 1 Transcription and Productive Infection. J. Virol. 2017, 91, e00904-17. [Google Scholar] [CrossRef] [Green Version]
- Erlandsson, A.C.; Bladh, L.-C.; Stierna Yucel-Lindberg, P.; Hammersten, O.; Modeer, T.; Harmenberg, T.; Wikstrom, A.C. Herpes simplex virus type 1 infection and glucocorticoid treatment regulate viral yield, glucocorticoid receptor and NF-kB levels. J. Endocrinol. 2002, 175, 165–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostler, J.; Harrison, K.S.; Schroeder, K.; Thunuguntla, P.; Jones, C. The glucocorticoid receptor (GR) stimulates Herpes Simplex Virus 1 productive infection, in part because the infected cell protein 0 (ICP0) promoter is cooperatively transactivated by the GR and Kruppel-like transcription factor 15. J. Virol. 2019, 93, e02063-18. [Google Scholar] [CrossRef] [Green Version]
- Oakley, R.H.; Cidlowski, A.C. The biology of the glucocorticoid receptor: New signaling mechanisms in health and disease. J. Allergy Clin. Immunol. 2013, 132, 1033–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Workman, A.; Eudy, J.; Smith, L.; Da Silva, L.F.; Sinani, D.; Bricker, H.; Cook, E.; Doster, A.; Jones, C. Cellular Transcription Factors Induced in Trigeminal Ganglia during Dexamethasone-Induced Reactivation from Latency Stimulate Bovine Herpesvirus 1 Productive Infection and Certain Viral Promoters. J. Virol. 2011, 86, 2459–2473. [Google Scholar] [CrossRef] [Green Version]
- McDonnell, B.B.; Yang, V.W. Mammalian Kruppel-like factors in health and diseases. Physiol. Rev. 2010, 90, 1337–1381. [Google Scholar] [CrossRef]
- Sinani, D.; Cordes, E.; Workman, E.; Thunuguntia, P.; Jones, C. Stress induced cellular transcription factors expressed in trigeminal ganglionic neurons stimulate the herpes simplex virus type 1 (HSV-1) infected cell protein 0 (ICP0) promoter. J. Virol. 2013, 87, 1183–1192. [Google Scholar] [CrossRef] [Green Version]
- Mallipattu, S.K.; Guo, Y.; Revelo, M.P.; Roa-Peña, L.; Miller, T.; Ling, J.; Shankland, S.J.; Bialkowska, A.B.; Ly, V.; Estrada, C.; et al. Kruppel–Like Factor 15 Mediates Glucocorticoid-Induced Restoration of Podocyte Differentiation Markers. J. Am. Soc. Nephrol. 2017, 28, 166–184. [Google Scholar] [CrossRef] [Green Version]
- Russell, W.C. A Sensitive and Precise Plaque Assay for Herpes Virus. Nature 1962, 195, 1028–1029. [Google Scholar] [CrossRef]
- Tremblay, R.G.; Sikorska, M.; Sandhu, J.K.; Lanthier, P.; Ribecco-Lutkiewicz, M.; Bani-Yaghoub, M. Differentiation of mouse Neuro 2A cells into dopamine neurons. J. Neurosci. Methods 2010, 186, 60–67. [Google Scholar] [CrossRef]
- Thunuguntla, P.; El-Mayet, F.S.; Jones, C. Bovine herpesvirus 1 can efficiently infect the human (SH-SY5Y) but not the mouse neuroblastoma cell line (Neuro-2A). Virus Res. 2017, 232, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Boutell, C.; Everret, R.D. Regulation of alphaherpesvirus infections by the ICP0 family of proteins. J. Gen. Virol. 2013, 94, 465–481. [Google Scholar] [CrossRef] [PubMed]
- Kaczynski, J.; Cook, T.; Urrutia, R. Sp1- and Krüppel-like transcription factors. Genome Biol. 2003, 4, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bieker, J.J. Krüppel-like Factors: Three Fingers in Many Pies. J. Biol. Chem. 2001, 276, 34355–34358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, K.A.; Tjian, R. Sp1 binds to promoter sequences and activates herpes simples virus ’immediate-early’ gene transcription in vitro. Nature 1985, 317, 179–182. [Google Scholar] [CrossRef] [PubMed]
- Black, A.R.; Black, J.D.; Azizkhan-Clifford, J. Sp1 and Kruppel-like transcription factor family of transcription factors in cell growth and cancer. J. Cell Physiol. 2001, 188, 143–160. [Google Scholar] [CrossRef]
- Mangan, S.; Alon, U. Structure and function of the feed-forward loop network motif. Proc. Natl. Acad. Sci. USA 2003, 100, 11980–11985. [Google Scholar] [CrossRef] [Green Version]
- Sasse, S.K.; Zuo, Z.; Kadiyala, V.; Zhang, L.; Pufall, M.A.; Jain, M.K.; Phang, T.L.; Stormo, G.D.; Gerber, A.N. Response Element Composition Governs Correlations between Binding Site Affinity and Transcription in Glucocorticoid Receptor Feed-forward Loops. J. Biol. Chem. 2015, 290, 19756–19769. [Google Scholar] [CrossRef] [Green Version]
- Sasse, S.K.; Mailloux, C.M.; Barczak, A.J.; Wang, Q.; Altonsy, M.O.; Jain, M.K.; Haldar, S.M.; Gerber, A.N. The Glucocorticoid Receptor and KLF15 Regulate Gene Expression Dynamics and Integrate Signals through Feed-Forward Circuitry. Mol. Cell. Biol. 2013, 33, 2104–2115. [Google Scholar] [CrossRef] [Green Version]
- Inman, M.; Lovato, L.; Doster, A.; Jones, C. A Mutation in the Latency-Related Gene of Bovine Herpesvirus 1 Disrupts the Latency Reactivation Cycle in Calves. J. Virol. 2002, 76, 6771–6779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, C.; Newby, T.J.; Holt, T.; Doster, A.; Stone, M.; Ciacci-Zanella, J.; Webster, C.J.; Jackwood, M.W. Analysis of latency in cattle after inoculation with a temperature sensitive mutant of bovine herpesvirus 1 (RLB106). Vaccine 2000, 18, 3185–3195. [Google Scholar] [CrossRef]
- Winkler, M.; Doster, A.; Sur, J.-H.; Jones, C. Analysis of bovine trigeminal ganglia following infection with bovine herpesvirus 1. Vet. Microbiol. 2002, 86, 139–155. [Google Scholar] [CrossRef]
- Winkler, M.T.; Doster, A.; Jones, C. Persistence and reactivation of bovine herpesvirus 1 in the tonsil of latently infected calves. J. Virol. 2000, 74, 5337–5346. [Google Scholar] [CrossRef]
- Frizzo da Silva, L.; Kook, I.; Doster, A.; Jones, C. Bovine herpesvirus 1 regulatory proteins, bICP0 and VP16, are readily detected in trigeminal ganglionic neurons expressing the glucocorticoid receptor during the early stages of reactivation from latency. J. Virol. 2013, 87, 11214–11222. [Google Scholar] [CrossRef] [Green Version]
- Kook, I.; Doster, A.; Jones, C. Bovine herpesvirus 1 regulatory proteins are detected in trigeminal ganglionic neurons during the early stages of stress-induced escape from latency. J. Neurovirol. 2015, 21, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Ostler, J.B.; Thunungutla, P.; Hendrickson, B.Y.; Jones, C. Transactivation of HSV-1 infected cell protein 4 (ICP4) enhancer by glucocorticoid receptor and stress-induced transcription factors requires overlapping Krüppel like transcription factor 4/Sp1 binding sites. J. Virol. 2021, 95, e01776-20. [Google Scholar] [CrossRef] [PubMed]
- El-Mayet, F.S.; Sawant, L.; Thunuguntla, P.; Zhao, J.; Jones, C. Two Pioneer Transcription Factors, Krüppel-Like Transcription Factor 4 and Glucocorticoid Receptor, Cooperatively Transactivate the Bovine Herpesvirus 1 ICP0 Early Promoter and Stimulate Productive Infection. J. Virol. 2020, 94. [Google Scholar] [CrossRef] [PubMed]
- Everett, R.D. ICP0, a regulator of herpes simplex virus during lytic and latent infection. Bioessays 2000, 22, 761–770. [Google Scholar] [CrossRef]
- Cliffe, A.R.; Knipe, D.M. Herpes Simplex Virus ICP0 Promotes both Histone Removal and Acetylation on Viral DNA during Lytic Infection. J. Virol. 2008, 82, 12030–12038. [Google Scholar] [CrossRef] [Green Version]
- Poon, A.P.W.; Gu, H.; Roizman, B. ICP0 and the US3 protein kinase of herpes simplex virus 1 independently block histone deacetylation to enable gene expression. Proc. Natl. Acad. Sci. USA 2006, 103, 9993–9998. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Jones, C. The Bovine Herpesvirus 1 Immediate-Early Protein (bICP0) Associates with Histone Deacetylase 1 To Activate Transcription. J. Virol. 2001, 75, 9571–9578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Jiang, Y.; Zhou, J.; Jones, C. The bovine herpes virus 1 (BHV-1) immediate early protein (bICP0) interacts with the histone acetyltransferase p300, and these interactions correlate with stimulation of gC promoter activity. J. Gen. Virol. 2006, 87, 1843–1851. [Google Scholar] [CrossRef] [PubMed]
- Boutell, C.; Sadis, S.; Everett, R.D. Herpes Simplex Virus Type 1 Immediate-Early Protein ICP0 and Its Isolated RING Finger Domain Act as Ubiquitin E3 Ligases In Vitro. J. Virol. 2002, 76, 841–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boutell, C.; Everett, R.D. The Herpes Simplex Virus Type 1 (HSV-1) Regulatory Protein ICP0 Interacts with and Ubiquitinates p53. J. Biol. Chem. 2003, 278, 36596–36602. [Google Scholar] [CrossRef] [Green Version]
- Diao, L.; Qiao, W.; Chen, Q.; Wang, C.; Geng, Y. bICP0 and its RING domain act as ubiquitin E3 ligases in vitro. Chin. Sci. Bull. 2005, 50, 636–640. [Google Scholar] [CrossRef]
- Parkinson, J.; Everett, R.D. Alphaherpesvirus Proteins Related to Herpes Simplex Virus Type 1 ICP0 Induce the Formation of Colocalizing, Conjugated Ubiquitin. J. Virol. 2001, 75, 5357–5362. [Google Scholar] [CrossRef] [Green Version]
- Van Sant, C.; Hagglund, R.; Lopez, P.; Roizman, B. The infected cell protein 0 of herpes simplex virus 1 dynamically interacts with proteasomes, binds and activates the cdc34 E2 ubiquitin- conjugating enzyme, and possesses in vitro E3 ubiquitin ligase activity. Proc. Natl. Acad. Sci. USA 2001, 98, 8815–8820. [Google Scholar] [CrossRef] [Green Version]
- Uchida, S.; Tanaka, Y.; Ito, H.; Saitoh-Ohara, F.; Inazawa, J.; Yokoyama, K.K.; Sasaki, S.; Marumo, F. Transcriptional regulation of the CLC-K1 by myc-associated zinc finger protein, a novel zinc finger repressor. Mol. Cell. Biol. 2000, 20, 7319–7331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, K.; Yahagi, N.; Aita, Y.; Murayama, Y.; Sawada, Y.; Piao, X.; Toya, N.; Oya, Y.; Shikama, A.; Takarada, A.; et al. KLF15 enables switching between lipogenesis aand gluconeogenesis during fasting. Cell Rep. 2016, 16, 2373–2386. [Google Scholar]
- Du, X.; Rosenfield, R.L.; Qin, K. KLF15 is a transcriptional regulator of the human 17beta-hydroxysteroid dehydrogenase type 5 gene. A potential link between regulation of testosterone production and fat stores in women. J. Clin. Endocrinol. Metab. 2009, 94, 2594–2601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otteson, D.C.; Lai, H.; Liu, Y.; Zack, D.J. Zinc-finger domains of the transcriptional repressor KLF15 bind multiple sites in rhodopsin and IRBP promoters including the CRS-1 and G-rich repressor elements. BMC Mol. Biol. 2005, 6, 15. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-mayet, F.S.; Harrison, K.S.; Jones, C. Regulation of Krüppel-Like Factor 15 Expression by Herpes Simplex Virus Type 1 or Bovine Herpesvirus 1 Productive Infection. Viruses 2021, 13, 1148. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061148
El-mayet FS, Harrison KS, Jones C. Regulation of Krüppel-Like Factor 15 Expression by Herpes Simplex Virus Type 1 or Bovine Herpesvirus 1 Productive Infection. Viruses. 2021; 13(6):1148. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061148
Chicago/Turabian StyleEl-mayet, Fouad S., Kelly S. Harrison, and Clinton Jones. 2021. "Regulation of Krüppel-Like Factor 15 Expression by Herpes Simplex Virus Type 1 or Bovine Herpesvirus 1 Productive Infection" Viruses 13, no. 6: 1148. https://0-doi-org.brum.beds.ac.uk/10.3390/v13061148