
Inhibition of Orbivirus Replication by Fluvastatin and Identification of the Key Elements of the Mevalonate Pathway Involved


 ,
, 
Abstract
:1. Introduction
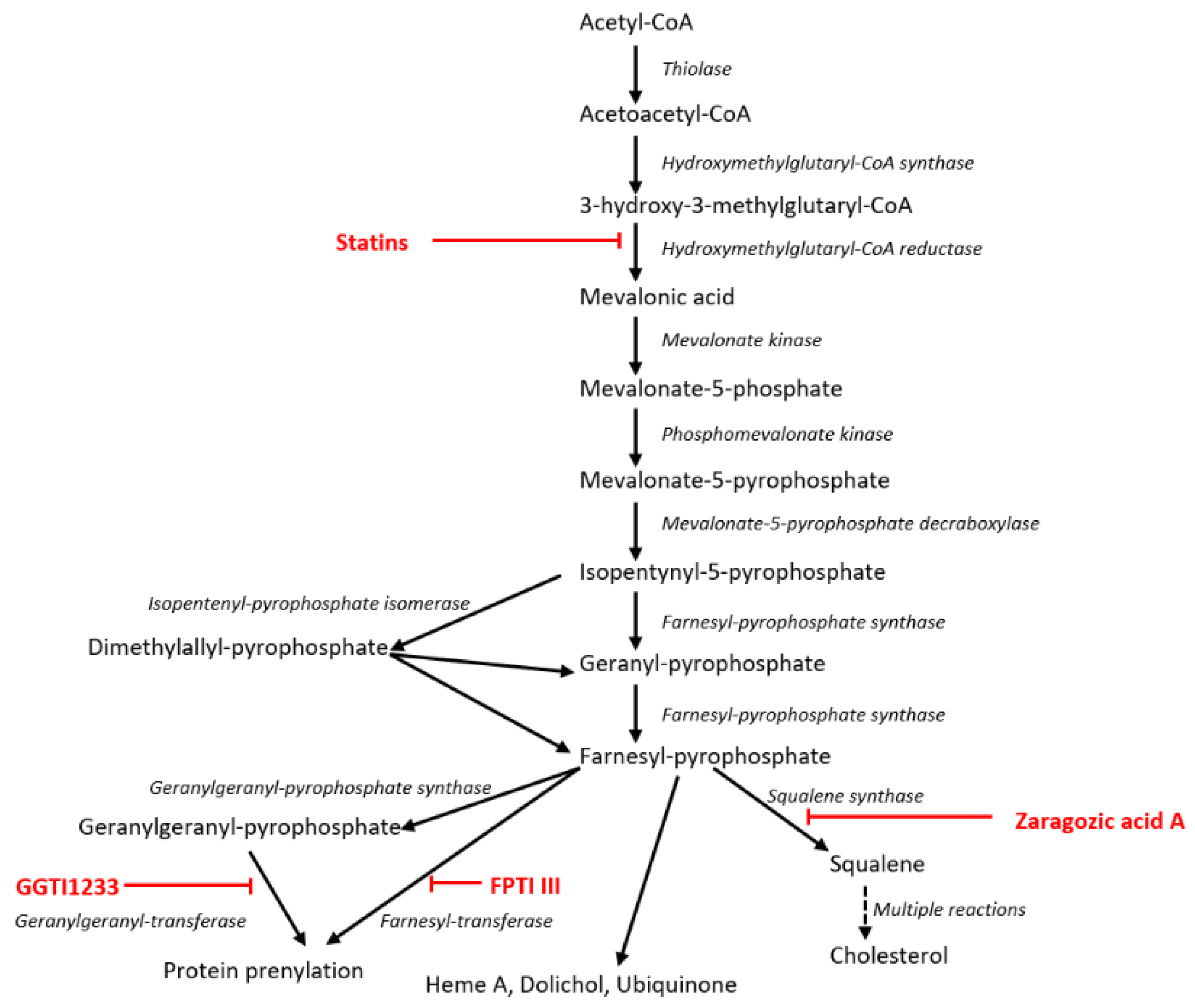
2. Materials and Methods
2.1. Lipid Depletion of Foetal Bovine Serum (FBS)
2.2. Chemical Compounds
2.3. Dosage of Fluvastatin in Mice
2.4. Cell Lines and Viruses
2.5. Plaque Assay
2.6. Cytotoxicity of Fluvastatin Treatment Assessed by an MTT Cell Proliferation Assay
2.7. Antibodies to Great Island Virus (GIV) NS4, Bluetongue Virus 8 Fourth Non-Structural Protein (BTV-8 NS4) and BTV-8 VP2
2.8. RNA Extraction and Real-Time Polymerase Chain Reaction (PCR)
2.9. Identification of Culicoides 3-Hydroxy-3-Methylglutaryl-CoA (HMG-CoA) Reductase
2.10. Pharmacological Inhibition of Virus Replication
2.10.1. Yellow Fever Virus 17D Strain
2.10.2. Orbiviruses (BTV-8 and GIV) in Mammalian and/or Insect Cells
2.11. Treatment of IFNAR(−/−) Mice with Fluvastatin and Infection with BTV
2.12. Statistical Analyses
3. Results
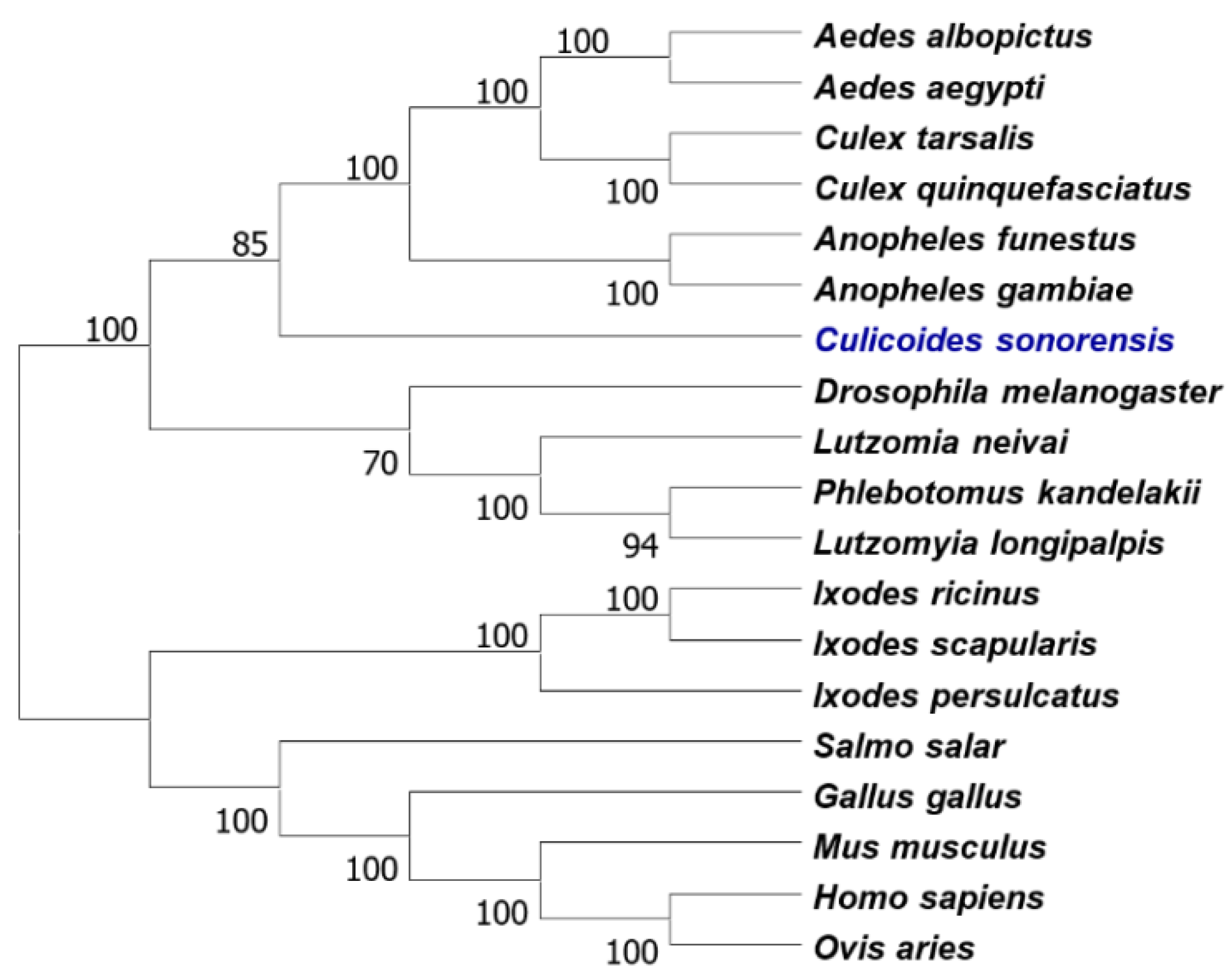
3.1. Identification of Culicoides HMG-CoA Reductase
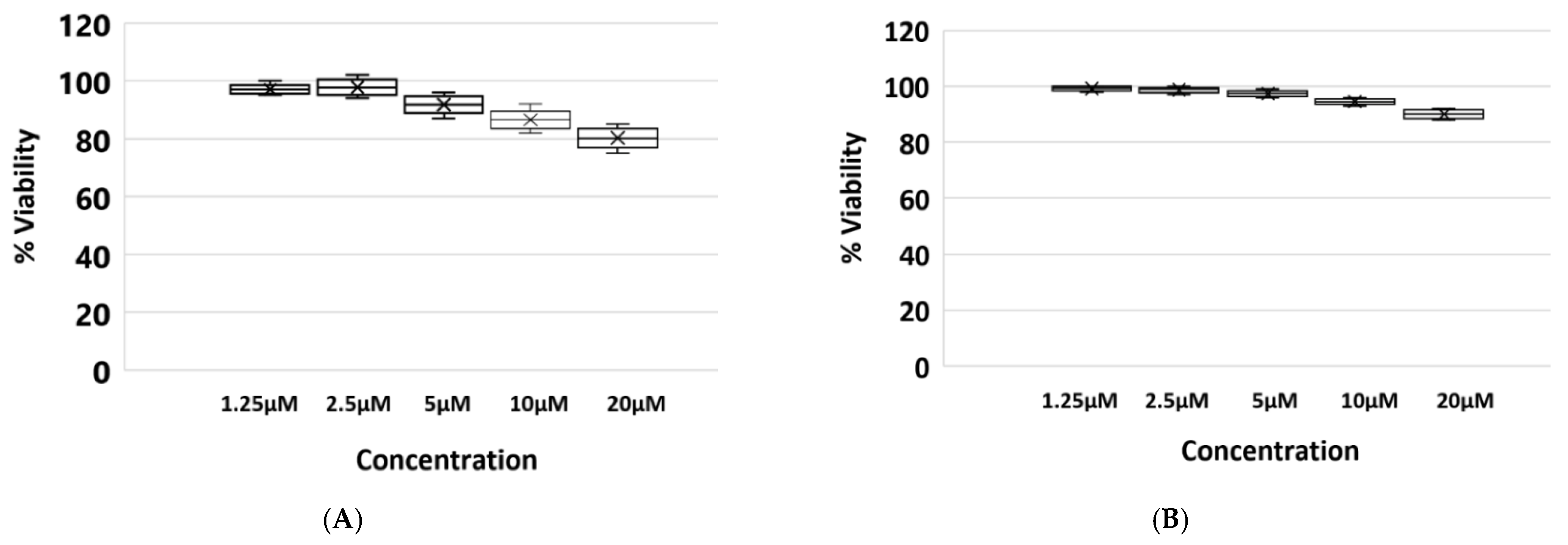
3.2. Cell Viability Assay in Presence of Fluvastatin
3.3. Fluvastatin and Replication of Yellow Fever Virus YFV17D in BSR Cells
3.4. Fluvastatin and Replication of Orbiviruses in Mammalian Cells
3.5. Fluvastatin and Replication of BTV-8 in Culicoides KC Cells
3.6. Fluvastatin and Replication of Orbiviruses in IFNAR(−/−) Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Attoui, H.; Mertens, P.P.C.; Becnel, J.; Belaganahalli, M.; Bergoin, M.; Brussaard, C.P.; Chappell, J.D.; Ciarlet, M.; del Vas, M.; Dermody, T.S.; et al. Reoviridae. In Virus Taxonomy. The Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier-Academic Press: London, UK, 2012. [Google Scholar]
- Attoui, H.; Mohd Jaafar, F. Zoonotic and emerging orbivirus infections. Rev. Sci. Tech. 2015, 34, 353–361. [Google Scholar] [CrossRef] [Green Version]
- Maan, S.; Maan, N.S.; Nomikou, K.; Veronesi, E.; Bachanek-Bankowska, K.; Belaganahalli, M.N.; Attoui, H.; Mertens, P.P. Complete genome characterisation of a novel 26th bluetongue virus serotype from Kuwait. PLoS ONE 2011, 6, e26147. [Google Scholar] [CrossRef] [Green Version]
- Ries, C.; Vogtlin, A.; Hussy, D.; Jandt, T.; Gobet, H.; Hilbe, M.; Burgener, C.; Schweizer, L.; Hafliger-Speiser, S.; Beer, M.; et al. Putative Novel Atypical BTV Serotype ‘36’ Identified in Small Ruminants in Switzerland. Viruses 2021, 13, 721. [Google Scholar] [CrossRef]
- Attoui, H.; Mendez-Lopez, M.R.; Rao, S.; Hurtado-Alendes, A.; Lizaraso-Caparo, F.; Mohd Jaafar, F.; Samuel, A.R.; Belhouchet, M.; Pritchard, L.I.; Melville, L.; et al. Peruvian horse sickness virus and Yunnan orbivirus, isolated from vertebrates and mosquitoes in Peru and Australia. Virology 2009, 394, 298–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breard, E.; Belbis, G.; Viarouge, C.; Nomikou, K.; Haegeman, A.; De Clercq, K.; Hudelet, P.; Hamers, C.; Moreau, F.; Lilin, T.; et al. Evaluation of adaptive immune responses and heterologous protection induced by inactivated bluetongue virus vaccines. Vaccine 2015, 33, 512–518. [Google Scholar] [CrossRef]
- Calvo-Pinilla, E.; de la Poza, F.; Gubbins, S.; Mertens, P.P.; Ortego, J.; Castillo-Olivares, J. Antiserum from mice vaccinated with modified vaccinia Ankara virus expressing African horse sickness virus (AHSV) VP2 provides protection when it is administered 48h before, or 48h after challenge. Antivir. Res. 2015, 116, 27–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smee, D.F.; Sidwell, R.W.; Clark, S.M.; Barnett, B.B.; Spendlove, R.S. Inhibition of bluetongue and Colorado tick fever orbiviruses by selected antiviral substances. Antimicrob. Agents Chemother. 1981, 20, 533–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, L.; Musiienko, V.; Bai, Z.; Qin, A.; Schneller, S.W.; Li, Q. Novel virostatic agents against bluetongue virus. PLoS ONE 2012, 7, e43341. [Google Scholar] [CrossRef] [Green Version]
- Alonso, C.; Utrilla-Trigo, S.; Calvo-Pinilla, E.; Jimenez-Cabello, L.; Ortego, J.; Nogales, A. Inhibition of Orbivirus Replication by Aurintricarboxylic Acid. Int. J. Mol. Sci. 2020, 21, 7294. [Google Scholar] [CrossRef] [PubMed]
- Mertens, P.P.; Brown, F.; Sangar, D.V. Assignment of the genome segments of bluetongue virus type 1 to the proteins which they encode. Virology 1984, 135, 207–217. [Google Scholar] [CrossRef]
- Belhouchet, M.; Mohd Jaafar, F.; Firth, A.E.; Grimes, J.M.; Mertens, P.P.; Attoui, H. Detection of a fourth orbivirus non-structural protein. PLoS ONE 2011, 6, e25697. [Google Scholar] [CrossRef] [Green Version]
- Ratinier, M.; Caporale, M.; Golder, M.; Franzoni, G.; Allan, K.; Nunes, S.F.; Armezzani, A.; Bayoumy, A.; Rixon, F.; Shaw, A.; et al. Identification and characterization of a novel non-structural protein of bluetongue virus. PLoS Pathog. 2011, 7, e1002477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyatt, A.D.; Zhao, Y.; Roy, P. Release of bluetongue virus-like particles from insect cells is mediated by BTV nonstructural protein NS3/NS3A. Virology 1993, 193, 592–603. [Google Scholar] [CrossRef] [PubMed]
- Fu, H. Mechanisms Controlling the Infection of Culicoides Biting Midges with Bluetongue Virus. Ph.D. Thesis, University of Hertfordshire, Hertfordshire, UK, 1995. [Google Scholar]
- Sirtori, C.R. The pharmacology of statins. Pharmacol. Res. 2014, 88, 3–11. [Google Scholar] [CrossRef]
- Buhaescu, I.; Izzedine, H. Mevalonate pathway: A review of clinical and therapeutical implications. Clin. Biochem. 2007, 40, 575–584. [Google Scholar] [CrossRef]
- Belles, X.; Martin, D.; Piulachs, M.D. The mevalonate pathway and the synthesis of juvenile hormone in insects. Annu. Rev. Entomol. 2005, 50, 181–199. [Google Scholar] [CrossRef] [Green Version]
- Peng, L.F.; Schaefer, E.A.; Maloof, N.; Skaff, A.; Berical, A.; Belon, C.A.; Heck, J.A.; Lin, W.; Frick, D.N.; Allen, T.M.; et al. Ceestatin, a novel small molecule inhibitor of hepatitis C virus replication, inhibits 3-hydroxy-3-methylglutaryl-coenzyme A synthase. J. Infect. Dis. 2011, 204, 609–616. [Google Scholar] [CrossRef]
- Skaff, D.A.; Ramyar, K.X.; McWhorter, W.J.; Barta, M.L.; Geisbrecht, B.V.; Miziorko, H.M. Biochemical and structural basis for inhibition of Enterococcus faecalis hydroxymethylglutaryl-CoA synthase, mvaS, by hymeglusin. Biochemistry 2012, 51, 4713–4722. [Google Scholar] [CrossRef] [Green Version]
- Quetglas, J.I.; Hernaez, B.; Galindo, I.; Munoz-Moreno, R.; Cuesta-Geijo, M.A.; Alonso, C. Small rho GTPases and cholesterol biosynthetic pathway intermediates in African swine fever virus infection. J. Virol. 2012, 86, 1758–1767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Rodriguez, A.V.; Tosteson, M.T. Role of simvastatin and methyl-beta-cyclodextrin [corrected] on inhibition of poliovirus infection. Biochem. Biophys. Res. Commun. 2006, 347, 51–59. [Google Scholar] [CrossRef]
- Potena, L.; Frascaroli, G.; Grigioni, F.; Lazzarotto, T.; Magnani, G.; Tomasi, L.; Coccolo, F.; Gabrielli, L.; Magelli, C.; Landini, M.P.; et al. Hydroxymethyl-glutaryl coenzyme a reductase inhibition limits cytomegalovirus infection in human endothelial cells. Circulation 2004, 109, 532–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gower, T.L.; Graham, B.S. Antiviral activity of lovastatin against respiratory syncytial virus in vivo and in vitro. Antimicrob. Agents Chemother. 2001, 45, 1231–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothwell, C.; Lebreton, A.; Young Ng, C.; Lim, J.Y.; Liu, W.; Vasudevan, S.; Labow, M.; Gu, F.; Gaither, L.A. Cholesterol biosynthesis modulation regulates dengue viral replication. Virology 2009, 389, 8–19. [Google Scholar] [CrossRef] [Green Version]
- Kapadia, S.B.; Chisari, F.V. Hepatitis C virus RNA replication is regulated by host geranylgeranylation and fatty acids. Proc. Natl. Acad. Sci. USA 2005, 102, 2561–2566. [Google Scholar] [CrossRef] [Green Version]
- Behnam, M.A.; Nitsche, C.; Boldescu, V.; Klein, C.D. The Medicinal Chemistry of Dengue Virus. J. Med. Chem. 2016, 59, 5622–5649. [Google Scholar] [CrossRef] [Green Version]
- del Real, G.; Jimenez-Baranda, S.; Mira, E.; Lacalle, R.A.; Lucas, P.; Gomez-Mouton, C.; Alegret, M.; Pena, J.M.; Rodriguez-Zapata, M.; Alvarez-Mon, M.; et al. Statins inhibit HIV-1 infection by down-regulating Rho activity. J. Exp. Med. 2004, 200, 541–547. [Google Scholar] [CrossRef] [Green Version]
- Ky, B.; Rader, D.J. The effects of statin therapy on plasma markers of inflammation in patients without vascular disease. Clin. Cardiol. 2005, 28, 67–70. [Google Scholar] [CrossRef]
- Houten, S.M.; Schneiders, M.S.; Wanders, R.J.; Waterham, H.R. Regulation of isoprenoid/cholesterol biosynthesis in cells from mevalonate kinase-deficient patients. J. Biol. Chem. 2003, 278, 5736–5743. [Google Scholar] [CrossRef] [Green Version]
- Agnese, S.T.; Spierto, F.W.; Hannon, W.H. Evaluation of four reagents for delipidation of serum. Clin. Biochem. 1983, 16, 98–100. [Google Scholar] [CrossRef]
- Brovkovych, V.; Aldrich, A.; Li, N.; Atilla-Gokcumen, G.E.; Frasor, J. Removal of Serum Lipids and Lipid-Derived Metabolites to Investigate Breast Cancer Cell Biology. Proteomics 2019, 19, e1800370. [Google Scholar] [CrossRef]
- Siekmeier, R.; Lattke, P.; Mix, C.; Park, J.W.; Jaross, W. Dose dependency of fluvastatin pharmacokinetics in serum determined by reversed phase HPLC. J. Cardiovasc. Pharmacol. Ther. 2001, 6, 137–145. [Google Scholar] [CrossRef]
- Nair, A.B.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef] [Green Version]
- Espano, E.; Nam, J.H.; Song, E.J.; Song, D.; Lee, C.K.; Kim, J.K. Lipophilic statins inhibit Zika virus production in Vero cells. Sci. Rep. 2019, 9, 11461. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Maeda, N.; Yoshida, H.; Urade, M.; Saito, S. Plaque formation of herpes virus hominis type 2 and rubella virus in variants isolated from the colonies of BHK21/WI-2 cells formed in soft agar. Arch. Virol. 1977, 53, 269–273. [Google Scholar] [CrossRef]
- Wechsler, S.J.; McHolland, L.E.; Wilson, W.C. A RNA virus in cells from Culicoides variipennis. J. Invertebr. Pathol. 1991, 57, 200–205. [Google Scholar] [CrossRef]
- Buckley, A.; Dawson, A.; Moss, S.R.; Hinsley, S.A.; Bellamy, P.E.; Gould, E.A. Serological evidence of West Nile virus, Usutu virus and Sindbis virus infection of birds in the UK. J. Gen. Virol. 2003, 84 (Pt 10), 2807–2817. [Google Scholar] [CrossRef]
- Mohd Jaafar, F.; Belhouchet, M.; Vitour, D.; Adam, M.; Breard, E.; Zientara, S.; Mertens, P.P.; Attoui, H. Immunisation with bacterial expressed VP2 and VP5 of bluetongue virus (BTV) protect alpha/beta interferon-receptor knock-out (IFNAR(−/−)) mice from homologous lethal challenge. Vaccine 2014, 32, 4059–4067. [Google Scholar] [CrossRef]
- Attoui, H.; Billoir, F.; Cantaloube, J.F.; Biagini, P.; de Micco, P.; de Lamballerie, X. Strategies for the sequence determination of viral dsRNA genomes. J. Virol. Methods 2000, 89, 147–158. [Google Scholar] [CrossRef]
- Hofmann, M.; Griot, C.; Chaignat, V.; Perler, L.; Thur, B. Bluetongue disease reaches Switzerland. Schweiz. Arch. Tierheilkd. 2008, 150, 49–56. [Google Scholar] [CrossRef]
- Debernard, S.; Rossignol, F.; Couillaud, F. The HMG-CoA reductase inhibitor fluvastatin inhibits insect juvenile hormone biosynthesis. Gen. Comp. Endocrinol. 1994, 95, 92–98. [Google Scholar] [CrossRef]
- Zapata, R.; Martin, D.; Piulachs, M.D.; Belles, X. Effects of hypocholesterolaemic agents on the expression and activity of 3-hydroxy-3-methylglutaryl-CoA reductase in the fat body of the German cockroach. Arch. Insect. Biochem. Physiol. 2002, 49, 177–186. [Google Scholar] [CrossRef]
- Zapata, R.; Piulachs, M.D.; Belles, X. Inhibitors of 3-hydroxy-3-methylglutaryl-CoA reductase lower fecundity in the German cockroach: Correlation between the effects on fecundity in vivo with the inhibition of enzymatic activity in embryo cells. Pest. Manag. Sci. 2003, 59, 1111–1117. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Tree, M.O.; Londono-Renteria, B.; Troupin, A.; Clark, K.M.; Colpitts, T.M.; Conway, M.J. Dengue virus reduces expression of low-density lipoprotein receptor-related protein 1 to facilitate replication in Aedes aegypti. Sci. Rep. 2019, 9, 6352. [Google Scholar] [CrossRef] [Green Version]
- Mackenzie, J.M.; Khromykh, A.A.; Parton, R.G. Cholesterol manipulation by West Nile virus perturbs the cellular immune response. Cell Host Microbe 2007, 2, 229–239. [Google Scholar] [CrossRef] [Green Version]
- Haeuptle, M.A.; Welti, M.; Troxler, H.; Hulsmeier, A.J.; Imbach, T.; Hennet, T. Improvement of dolichol-linked oligosaccharide biosynthesis by the squalene synthase inhibitor zaragozic acid. J. Biol. Chem. 2011, 286, 6085–6091. [Google Scholar] [CrossRef] [Green Version]
- Jabbar, T.K.; Calvo-Pinilla, E.; Mateos, F.; Gubbins, S.; Bin-Tarif, A.; Bachanek-Bankowska, K.; Alpar, O.; Ortego, J.; Takamatsu, H.H.; Mertens, P.P.; et al. Protection of IFNAR (−/−) mice against bluetongue virus serotype 8, by heterologous (DNA/rMVA) and homologous (rMVA/rMVA) vaccination, expressing outer-capsid protein VP2. PLoS ONE 2013, 8, e60574. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Gutierrez, M.; Castellanos, J.E.; Gallego-Gomez, J.C. Statins reduce dengue virus production via decreased virion assembly. Intervirology 2011, 54, 202–216. [Google Scholar] [CrossRef]
- Mohan, K.V.; Muller, J.; Atreya, C.D. Defective rotavirus particle assembly in lovastatin-treated MA104 cells. Arch. Virol. 2008, 153, 2283–2290. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Pinilla, E.; Rodriguez-Calvo, T.; Sevilla, N.; Ortego, J. Heterologous prime boost vaccination with DNA and recombinant modified vaccinia virus Ankara protects IFNAR(−/−) mice against lethal bluetongue infection. Vaccine 2009, 28, 437–445. [Google Scholar] [CrossRef]
- Martinez-Gutierrez, M.; Correa-Londono, L.A.; Castellanos, J.E.; Gallego-Gomez, J.C.; Osorio, J.E. Lovastatin delays infection and increases survival rates in AG129 mice infected with dengue virus serotype 2. PLoS ONE 2014, 9, e87412. [Google Scholar] [CrossRef] [PubMed]
- Norman, K.L.; Lee, P.W. Reovirus as a novel oncolytic agent. J. Clin. Investig. 2000, 105, 1035–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Chester, C.; Rajasekaran, N.; He, Z.; Kohrt, H.E. Strategic Combinations: The Future of Oncolytic Virotherapy with Reovirus. Mol. Cancer Ther. 2016, 15, 767–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Wu, T.Y.; Zhao, M.F.; Li, C.J. The balance of protein farnesylation and geranylgeranylation during the progression of nonalcoholic fatty liver disease. J. Biol. Chem. 2020, 295, 5152–5162. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
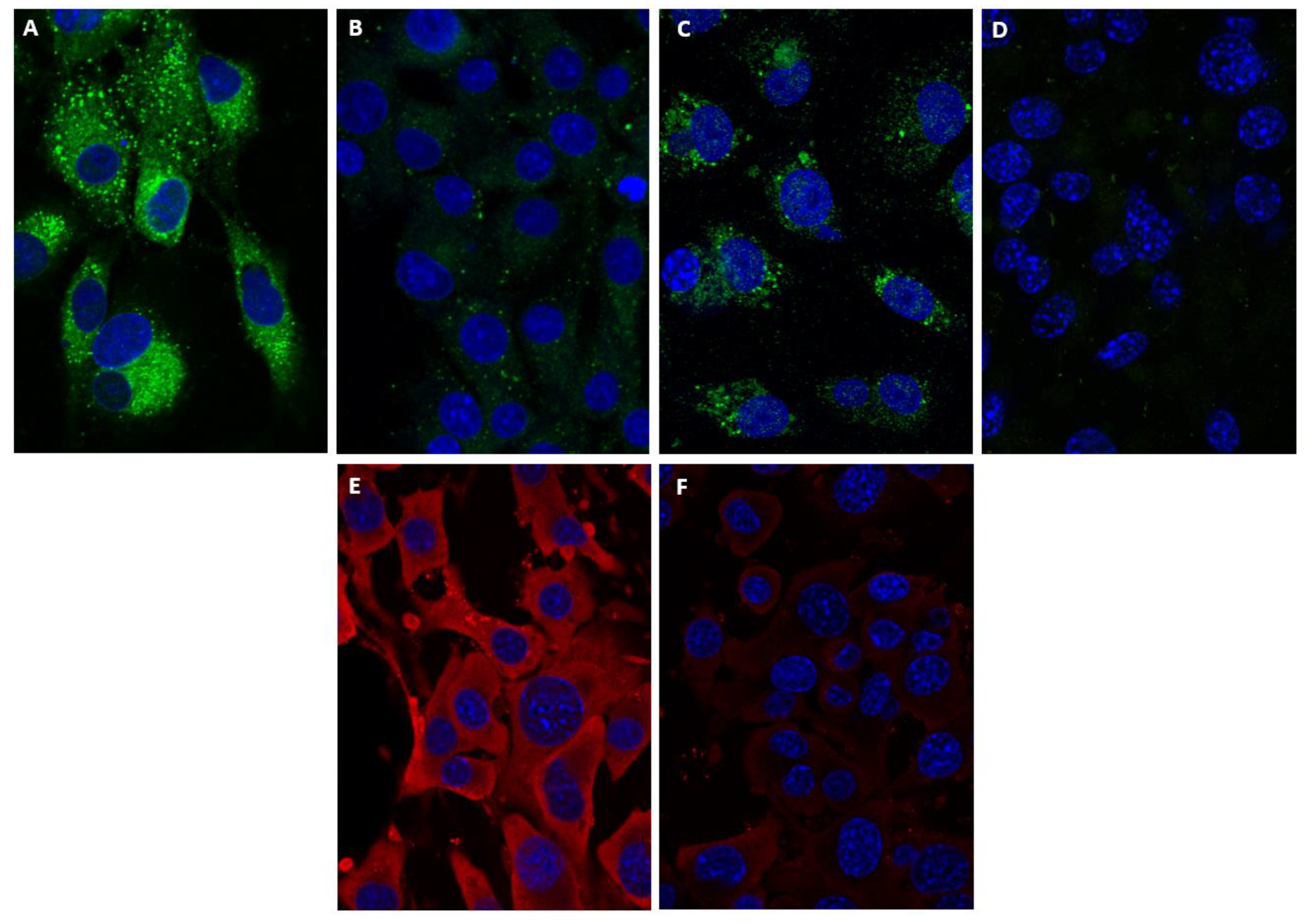{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Fluvastatin Dose | Route of Administration | Days Administered | Challenge Virus |
|---|---|---|---|---|
| A | None (solvent diluted in PBS) | oral | Daily (days 0 to 12) | None |
| B | None (solvent diluted in PBS) | oral | Daily (days 0 to 12) | BTV-8 (10 PFU/mouse) |
| C | 350 µg Fluvastatin/mouse (14.8 mg/kg/body weight: mouse-adapted dose) | oral | Daily (days 0 to 12) | None |
| D | 28 µg Fluvastatin/mouse (1.2 mg/kg body weight: higher end human daily dose/kg body weight) | oral | Daily (days 0 to 7) | BTV-8 (10 PFU/mouse) |
| E | 350 µg Fluvastatin/mouse | oral | Daily (days −1 to 12) | BTV-8 (10 PFU/mouse) |
| F | 350 µg Fluvastatin/mouse | oral | Daily (days −1 to 12) | BTV-8 (10 PFU/mouse) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohd Jaafar, F.; Monsion, B.; Belhouchet, M.; Mertens, P.P.C.; Attoui, H. Inhibition of Orbivirus Replication by Fluvastatin and Identification of the Key Elements of the Mevalonate Pathway Involved. Viruses 2021, 13, 1437. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081437
Mohd Jaafar F, Monsion B, Belhouchet M, Mertens PPC, Attoui H. Inhibition of Orbivirus Replication by Fluvastatin and Identification of the Key Elements of the Mevalonate Pathway Involved. Viruses. 2021; 13(8):1437. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081437
Chicago/Turabian StyleMohd Jaafar, Fauziah, Baptiste Monsion, Mourad Belhouchet, Peter P. C. Mertens, and Houssam Attoui. 2021. "Inhibition of Orbivirus Replication by Fluvastatin and Identification of the Key Elements of the Mevalonate Pathway Involved" Viruses 13, no. 8: 1437. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081437