
Emerging Viral Pathogens in Sturgeon Aquaculture in Poland: Focus on Herpesviruses and Mimivirus Detection


Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus Isolation on WSSK-1 Cell Line
2.2. Nucleic Acid Extraction for PCR Detection and Sequencing
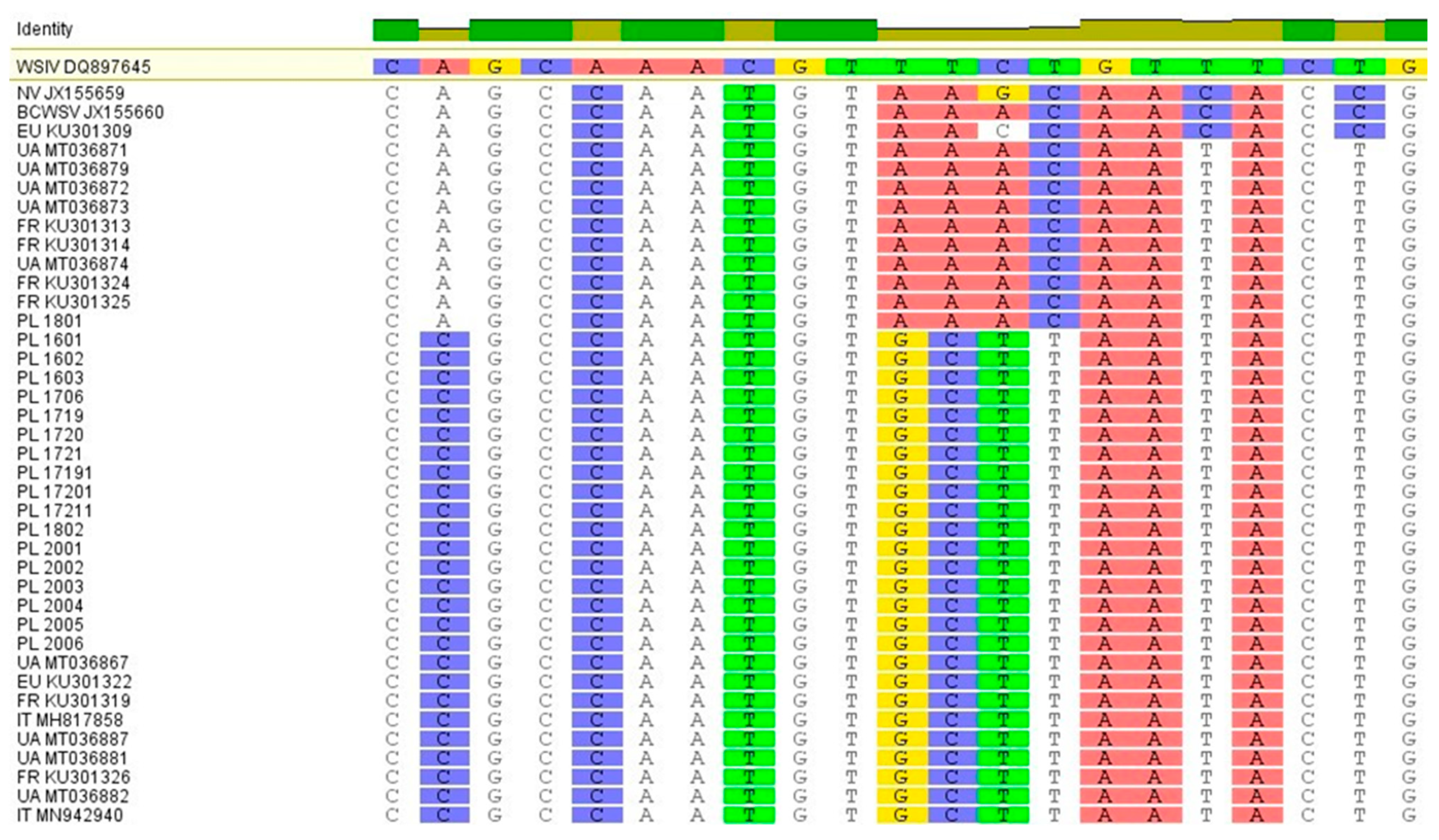
2.3. Sequence Analysis and Phylogenetic Tree Analysis
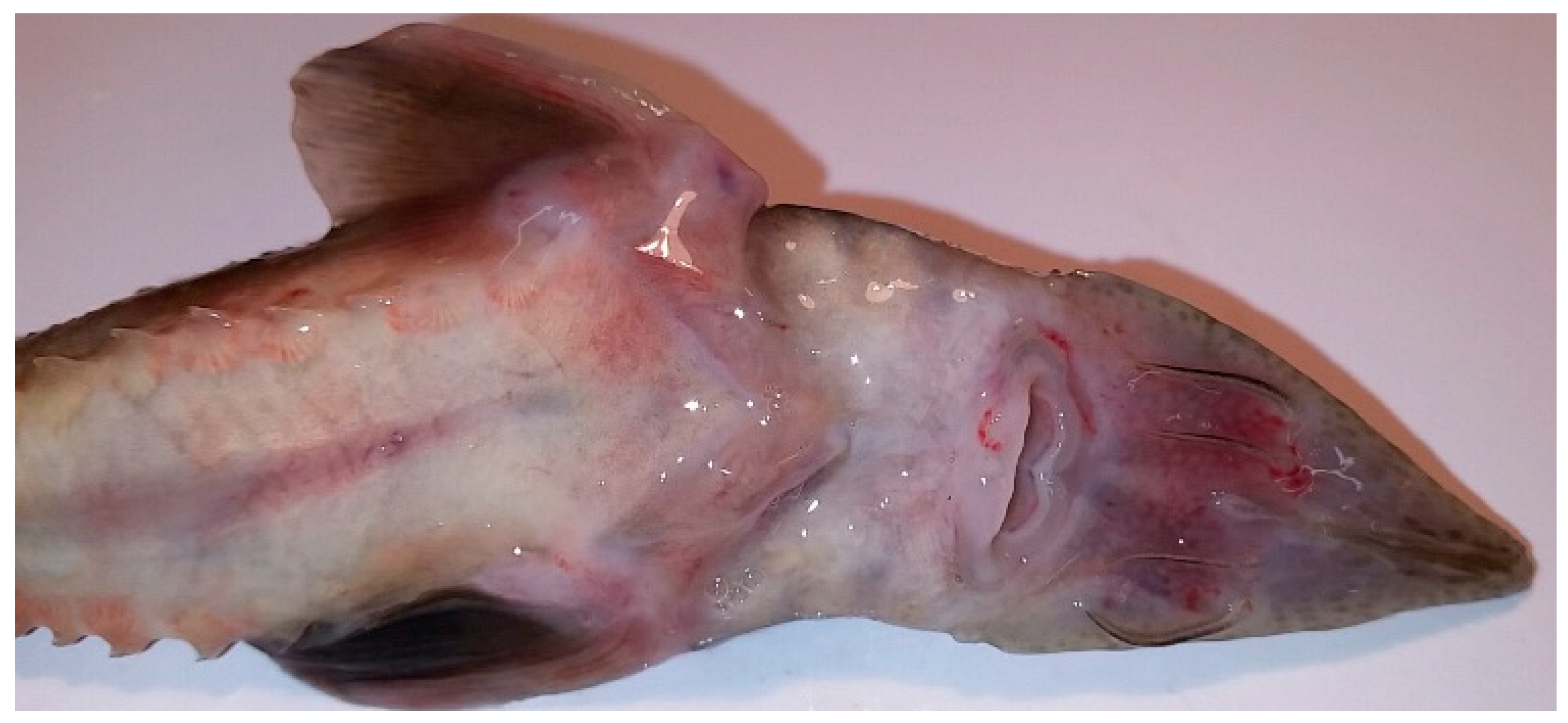
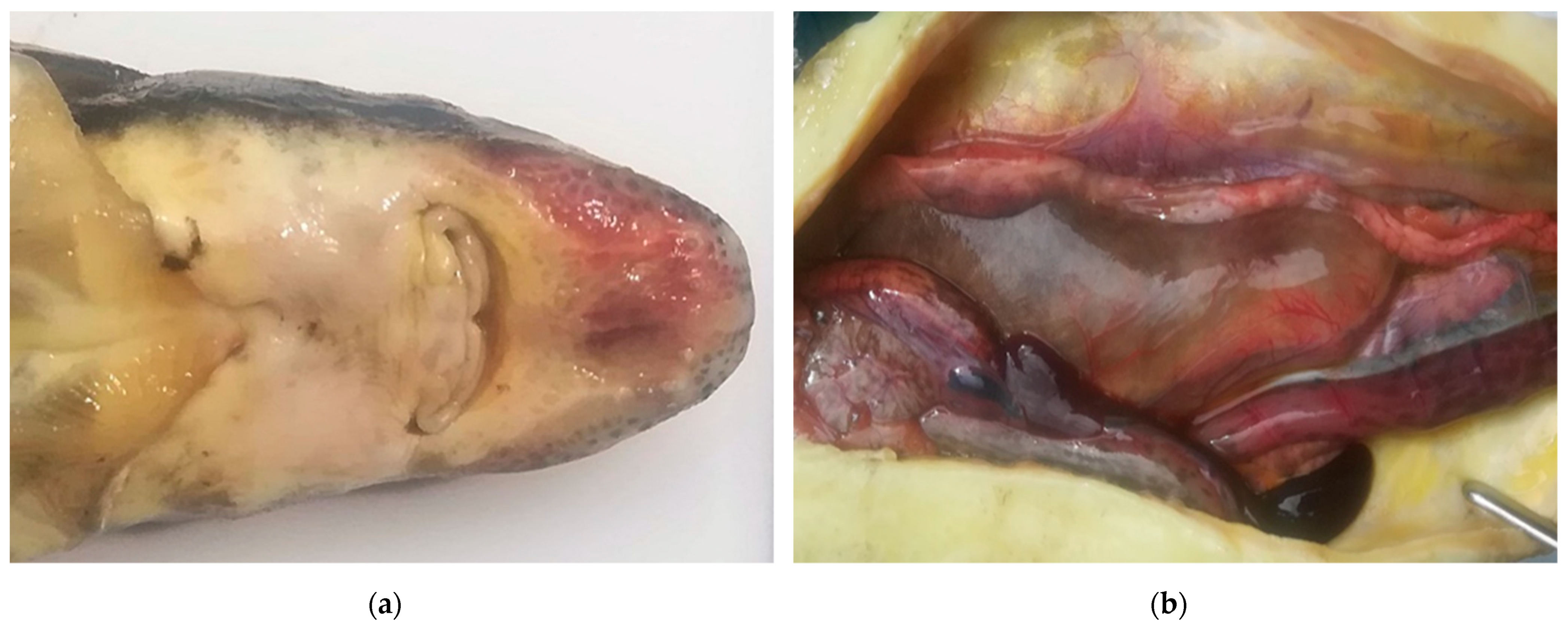
3. Results
Virus Isolation on WSSK-1 Cell Line and PCR Detection and Sequencing
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IUCN 2021. The IUCN Red List of Threatened Species. Version 2021-1. Available online: www.icunredlist.org (accessed on 1 May 2021).
- EUMOFA. The Caviar Market. Production, Trade and Consumption in and Outside the EU. Available online: www.eumofa.eu (accessed on 1 May 2021).
- Available online: www.cites.org (accessed on 1 May 2021).
- Bronzi, P.; Rosenthal, H. Present and future sturgeon and caviar production and marketing: A global market overview. J. Appl. Ichthyol. 2014, 30, 1536–1546. [Google Scholar] [CrossRef]
- Kolman, R.; Kapusta, A.; Duda, A.; Wiszniewski, G. Review of the current status of the Atlantic sturgeon Acipenser oxyrinchus oxyrinchus Mitchill 1815, in Poland: Principles, previous experience, and results. J. Appl. Ichthyol. 2011, 27, 186–191. [Google Scholar] [CrossRef]
- Mugetti, D.; Pastorino, P.; Menconi, V.; Pedron, C.; Prearo, M. The Old and the New on Viral Diseases in Sturgeon. Pathogens 2020, 9, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelley, G.O.; Waltzek, T.B.; McDowell, T.S.; Yun, S.C.; LaPatra, S.E.; Hedrick, R.P. Genetic Relationships among Herpes-Like Viruses Isolated from Sturgeon. J. Aquat. Anim. Health 2005, 17, 297–303. [Google Scholar] [CrossRef]
- Kurobe, T.; Kelley, G.O.; Waltzek, T.B.; Hedrick, R.P. Revised Phylogenetic Relationships among Herpesviruses Isolated from Sturgeons. J. Aquat. Anim. Health 2008, 20, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Lapatra, S.E.; Groff, J.M.; Keith, I.; Hogans, W.E.; Groman, D. Case report: Concurrent herpesviral and presumptive iridoviral infection associated with disease in cultured shortnose sturgeon, Acipenser brevirostrum (L.), from the Atlantic coast of Canada. J. Fish Dis. 2014, 37, 141–147. [Google Scholar] [CrossRef]
- Hedrick, R.; Groff, J.; McDowell, T.; Wingfield, W. An iridovirus infection of the integument of the white sturgeon Acipenser transmontanus. Dis. Aquat. Org. 1990, 8, 39–44. [Google Scholar] [CrossRef]
- Adkison, M.; Cambre, M.; Hedrick, R.P. Identification of an iridovirus in Russian sturgeon (Acipenser guldenstadi) from Northern Europe. Bull. Eur. Assoc. Fish Pathol. 1998, 18, 29–32. [Google Scholar]
- Clouthier, S.C.; Vanwalleghem, E.; Copeland, S.; Klassen, C.; Hobbs, G.; Nielsen, O.; Anderson, E.D. A new species of nu-cleo-cytoplasmic large DNA virus (NCLDV) associated with mortalities in Manitoba lake sturgeon Acipenser fulvescens. Dis. Aquat. Org. 2013, 102, 195–209. [Google Scholar] [CrossRef] [Green Version]
- Bigarré, L.; Lesne, M.; Lautraite, A.; Chesneau, V.; Leroux, A.; Jamin, M.; Boitard, P.M.; Toffan, A.; Prearo, M.; Labrut, S.; et al. Molecular identification of iridoviruses infecting various sturgeon species in Europe. J. Fish Dis. 2016, 40, 105–118. [Google Scholar] [CrossRef]
- Ciulli, S.; Volpe, E.; Sirri, R.; Passalacqua, P.; Bianchi, F.C.; Serratore, P.; Mandrioli, L. Outbreak of mortality in Russian (Acipenser gueldenstaedtii) and Siberian (Acipenser baerii) sturgeons associated with sturgeon nucleo-cytoplasmatic large DNA virus. Vet. Microbiol. 2016, 191, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Rud, Y.; Bigarré, L.; Pallandre, L.; Briand, F.X.; Buchatsky, L. First genetic characterization of sturgeon mimiviruses in Ukraine. J. Fish Dis. 2020, 43, 1391–1400. [Google Scholar] [CrossRef] [PubMed]
- Shchelkunov, I.S.; Shchelkunova, T.I.; Shchelkunov, A.I.; Kolbassova, Y.P.; Didenko, L.V.; Bykovsky, A.P. First detection of a viral agent causing disease in farmed sturgeon in Russia. Dis. Aquat. Org. 2009, 86, 193–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doszpoly, A.; Shchelkunov, I.S. Partial genome analysis of Siberian sturgeon alloherpesvirus suggests its close relation to AciHV-2—Short communication. Acta Vet. Hung. 2010, 58, 269–274. [Google Scholar] [CrossRef]
- Clouthier, S.C.; Van Walleghem, E.; Anderson, E.D. Sturgeon nucleo-cytoplasmic large DNA virus phylogeny and PCR tests. Dis. Aqua. Org. 2015, 117, 93–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clouthier, S.C.; Caskenette, A.; Van Walleghem, E.; Schroeder, T.; Macdonald, D.; Anderson, E.D. Molecular phylogeny of sturgeon mimiviruses and Bayesian hierarchical modeling of their effect on wild Lake Sturgeon (Acipenser fulvescens) in Central Canada. Infect. Genet. Evol. 2020, 84, 104491. [Google Scholar] [CrossRef] [PubMed]
- Pallandre, L.; Lesne, M.; de Boisséson, C.; Briand, F.X.; Charrier, A.; Waltzek, T.; Daniel, P.; Tragnan, A.; Debeuf, B.; Chesneau, V.; et al. Acipenser iridovirus-European encodes a replication facto r C (RFC) sub-unit. Arch. Virol. 2018, 163, 2985–2995. [Google Scholar] [CrossRef] [PubMed]
- Pallandre, L.; Lesne, M.; de Boisséson, C.; Charrier, A.; Daniel, P.; Tragnan, A.; Debeuf, B.; Chesneau, V.; Bigarré, L. Genetic identification of two Acipenser iridovirus-European variants using high-resolution melting analysis. J. Virol. Methods 2019, 265, 105–112. [Google Scholar] [CrossRef]
- Drennan, J.D.; Lapatra, S.E.; Samson, C.A.; Ireland, S.; Eversman, K.F.; Cain, K.D. Evaluation of lethal and non-lethal sam-pling methods for the detection of white sturgeon iridovirus infection in white sturgeon, Acipenser transmontanus (Richard-son). J. Fish Dis. 2007, 30, 367–379. [Google Scholar] [CrossRef]
- Hofsoe-Oppermann, P.; Kiełpińska, J.; Panicz, R.; Bergmann, S.M. Detection of white sturgeon iridovirus (WSIV) in sturgeons Actinopterygii: Acipenseriformes: Acipenseridae from aquaculture facilities located in Poland, Germany, and Italy. Acta Ichthyol. Piscat. 2019, 49, 257–264. [Google Scholar] [CrossRef]
- Hofsoe-Oppermann, P.; Kiełpińska, J.; Panicz, R.; Bergmann, S.M. Detection of white sturgeon iridovirus (WSIV) in wild sturgeons in Poland. J. Vet. Res. 2020, 64, 363–368. [Google Scholar] [CrossRef]
- Hedrick, R.P.; Groff, J.M.; McDowell, T.S. Isolation of an epitheliotropic herpesvirus from white sturgeon (Acipenser trans-montanus). Dis. Aquat. Organ. 1991, 11, 49–56. [Google Scholar] [CrossRef]
- GENEIOUS Software Version 10. Available online: www.geneious.com (accessed on 1 April 2021).
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef] [PubMed]
- Watson, L.R.; Yun, S.C.; Groff, J.M.; Hedrick, R.P. Characteristics and pathogenicity of a novel herpesvirus isolated from adult and subadult white sturgeon Acipenser transmontanus. Dis. Aquat. Org. 1995, 22, 199–210. [Google Scholar] [CrossRef] [Green Version]
- Hanson, L.; Doszpoly, A.; van Beurden, S.; Viadanna, P.D.O.; Waltzek, T. Alloherpesviruses of Fish. In Aquaculture Virology; Elsevier BV: Amsterdam, The Netherlands, 2016; pp. 153–172. [Google Scholar]
- Doszpoly, A.; Kalabekov, I.M.; Breyta, R.; Shchelkunov, I.S. Isolation and characterization of an atypical Siberian sturgeon herpesvirus strain in Russia: Novel North American Acipenserid herpesvirus 2 strain in Europe? J. Fish Dis. 2017, 40, 1363–1372. [Google Scholar] [CrossRef]
- Axén, C.; Vendramin, N.; Toffan, A. Outbreak of Mortality Associated with Acipenser Iridovirus European (AcIV-E) Detection in Siberian Sturgeon (Acipenser baerii) Farmed in Sweden. Fishes 2018, 3, 42. [Google Scholar] [CrossRef] [Green Version]
- Mugetti, D.; Pastorino, P.; Menconi, V.; Messina, M.; Masoero, L.; Ceresa, L.; Pedron, C.; Prearo, M. Increase in susceptibility of new sturgeon species to Acipenser Iridovirus European (AcIV-E) infection. Pathogens 2020, 9, 156. [Google Scholar] [CrossRef] [Green Version]
- Kwak, K.T.; Gardner, I.A.; Farver, T.B.; Hedrick, R.P. Rapid detection of white sturgeon iridovirus (WSIV) using a polymerase chain reaction (PCR) assay. Aquaculture 2006, 254, 92–101. [Google Scholar] [CrossRef]
- Raverty, S.; Hedrick, R.; Justine, H. Diagnosis of sturgeon iridovirus infection in farmed white sturgeon in British Columbia. Can. Vet. J. 2003, 44, 327–328. [Google Scholar] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | Sequence | Source |
|---|---|---|
| Host 16S rRNA | ||
| Real Time PCR | Aci16SFw: AAGGAACTCGGCAAACACGA Aci16SRv: GCTACCTTCGCACGGTCAAA Aci16SPb: Cy5-AGAGGTCCCGCCTGCCCTG-BHQ2 | [15] |
| Mimivirus | ||
| cPCR set A cPCR set D | F-oPVP339: GATAGGGTACAAGAGACATTC R-oPVP340: TCAACCAAATATTCTTGCTTG F-oPVP339: GATAGGGTACAAGAGACATTC R-oPVP342: TCAACCAAATATTCTTGCTTG | [13] |
| Real Time PCR | F-oPVP346: TCAAAGTCTGGGACCTCTA R-oPVP347: AGAGATGTTCAACTGGATGT tqPVP20:FAM-TTGTGAATCATATCGCCAGTCAT-BHQ1 | [13] |
| Herpesvirus | ||
| cPCR AciHV-1 cPCR AciHV-2 | F-308: 5′-ACCTCGTGTTGATCG-3′ R-309: 5′-TCAAAACTTCCGGGT-3′ TermF2 5′-GCMMGRGGACAGAWCCCMG-3′ Termsal-3Rdeg 5′-GGTGCACACRCCMADIGACG-3′, | [8] |
| Number of Farm /Year of Sampling | Description of Fish | Number of Sample | Number of Samples Positive/All | Number of Positive Samples (Real Time) | GenBank Accession Number |
|---|---|---|---|---|---|
| 1/2016 | Siberian sturgeon 1000–2000 g, 47 fish | PL1601-PL1647 | 7/47 | 1601 (s), 1602 (s), 1603 (s), 1615, 1616, 1625, 1626 | MG212658, MG212659, MG212660 |
| 2/2016 | Siberian sturgeon 5–10 g, 6 fish | PL1648-Pl1653 | 1/6 | 1650 | - |
| 3/2017 | Siberian sturgeon 500–700 g, 12 fish | PL1701-PL1712 | 6/12 | 1706 (s), 1702, 1703, 1704, 1705, 1708 | MG212661 |
| 4/2017 | Siberian sturgeon 15–50 g, 6 fish | PL1713-PL1718 | 0/6 | - | - |
| 5/2017 | Siberian sturgeon 30–50 g, 11 fish | PL1719-PL1729 | 8/11 | 1719 (s), 1719.1 (s), 1720 (s), 1720.1 (s), 1721 (s), 1721.1 (s), 1722, 1726 | MG212662, MG212663, MG212664, MG212665, MG212666, MG212667 |
| 6/2017 | Siberian sturgeon 20–50 g, 10 fish | PL1730-PL1739 | 0/10 | - | - |
| 7/2017 | Russian sturgeon 10–15 g, 10 fish | PL1740-PL1750 | 0/10 | - | - |
| 8/2018 | Russian sturgeon 1000–1500 g, 6 fish | PL1801-PL1806 | 2/6 | 1801 (s), 1802 (s) | MN542940, MN542941 |
| 9/2020 | Siberian sturgeon 10–50 g, 28 fish | PL2001-PL2028 | 6/28 | 2001 (s), 2002 (s), 2003 (s), 2004 (s), 2005 (s), 2006 (s) | MT735127, MT735128, MT735129, MT735130, MT735131, MT735132 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stachnik, M.; Matras, M.; Borzym, E.; Maj-Paluch, J.; Reichert, M. Emerging Viral Pathogens in Sturgeon Aquaculture in Poland: Focus on Herpesviruses and Mimivirus Detection. Viruses 2021, 13, 1496. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081496
Stachnik M, Matras M, Borzym E, Maj-Paluch J, Reichert M. Emerging Viral Pathogens in Sturgeon Aquaculture in Poland: Focus on Herpesviruses and Mimivirus Detection. Viruses. 2021; 13(8):1496. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081496
Chicago/Turabian StyleStachnik, Magdalena, Marek Matras, Ewa Borzym, Joanna Maj-Paluch, and Michał Reichert. 2021. "Emerging Viral Pathogens in Sturgeon Aquaculture in Poland: Focus on Herpesviruses and Mimivirus Detection" Viruses 13, no. 8: 1496. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081496