
Nuances of Whitefly Vector–Crinivirus Interactions Revealed in the Foregut Retention and Transmission of Lettuce Chlorosis Virus by Two Bemisia tabaci Cryptic Species

Abstract
:1. Introduction
2. Materials and Methods
2.1. Maintenance of Viruses and Whitefly Colonies
2.2. Virion Purification and Quantification
2.3. VRT Assays
2.4. Statistical Analysis
3. Results
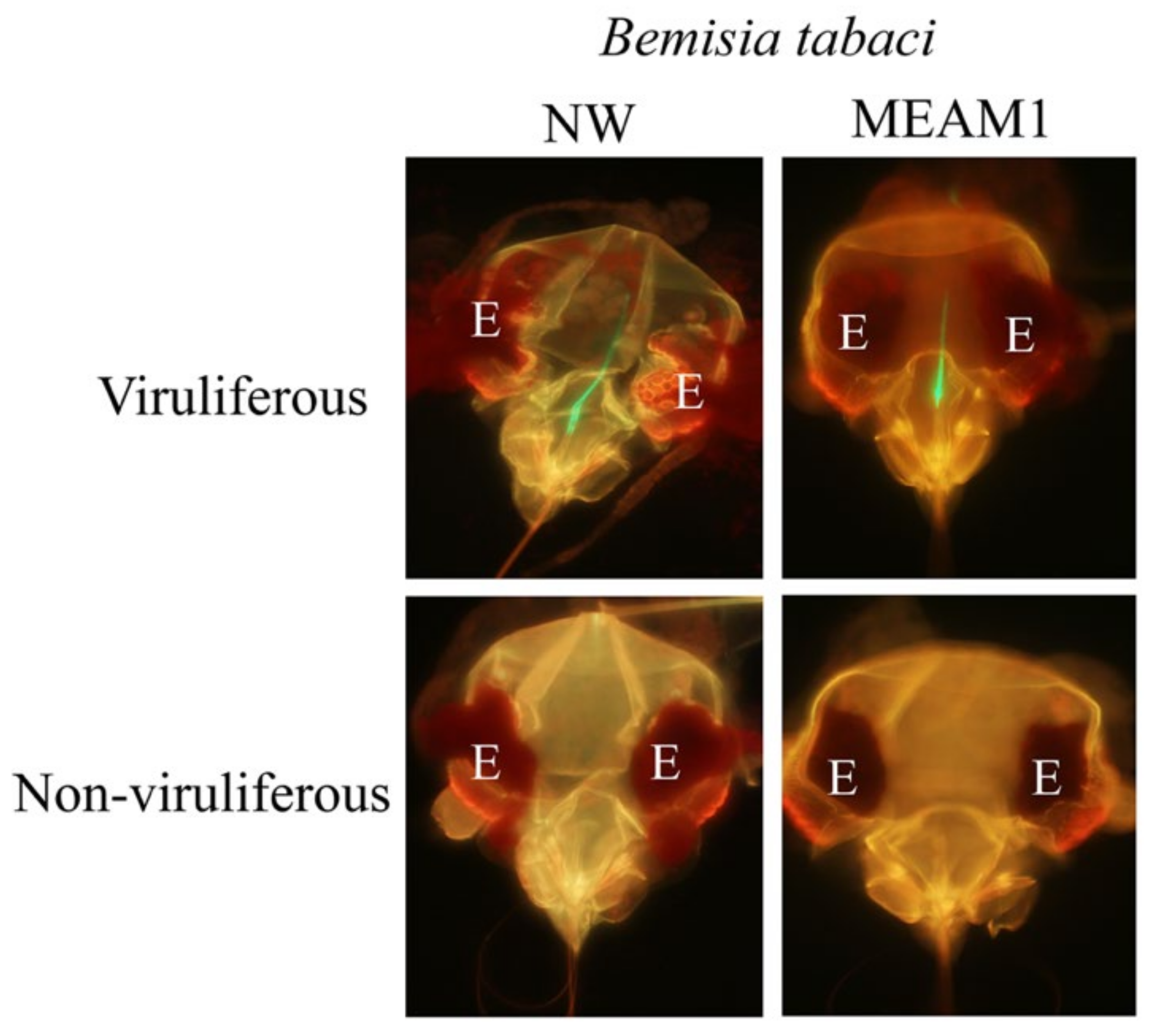
3.1. In Planta Acquired GH-LCV Is Retained in the Foreguts of B. tabaci NW and MEAM1 Vectors and Transmitted
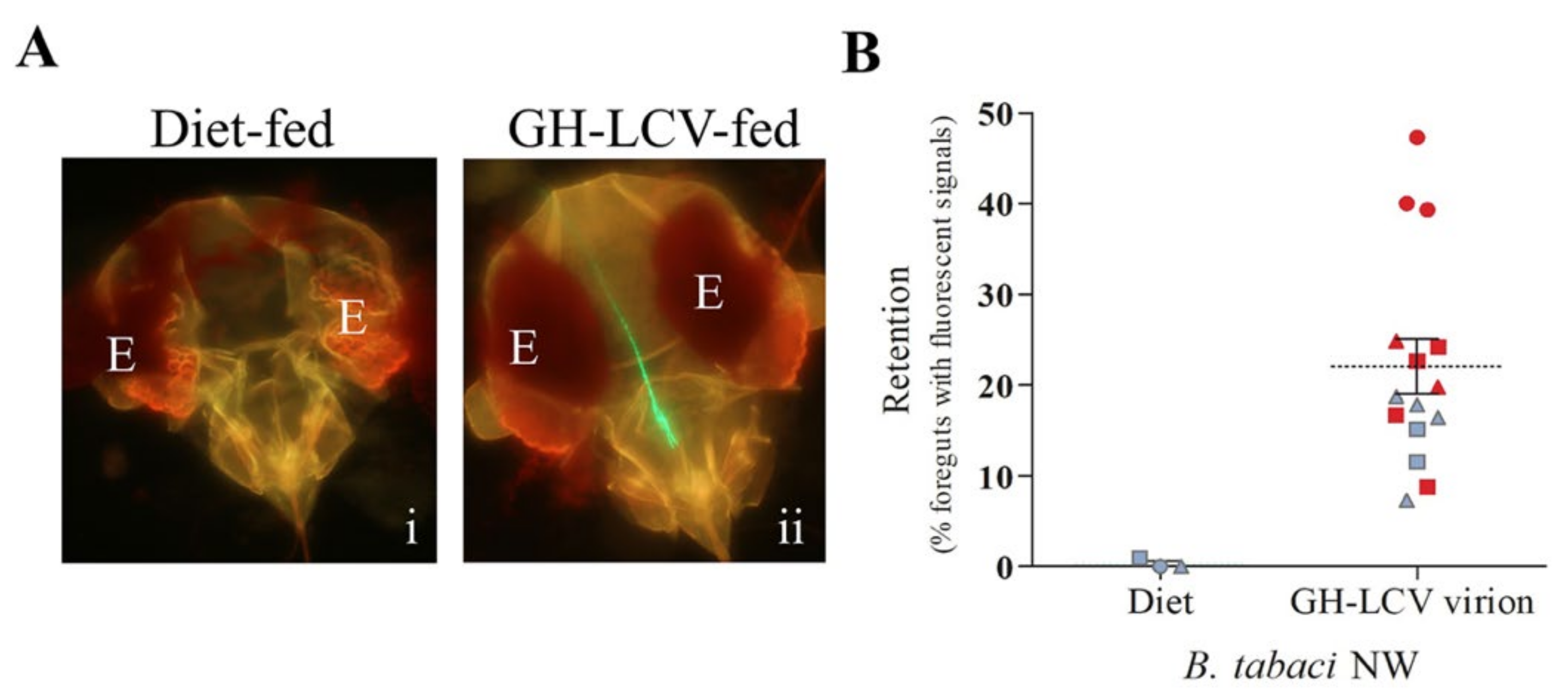
3.2. Purified GH-LCV Virions Are Retained in the Foreguts of B. tabaci NW Vectors and Transmitted upon Inoculation Access Feeding
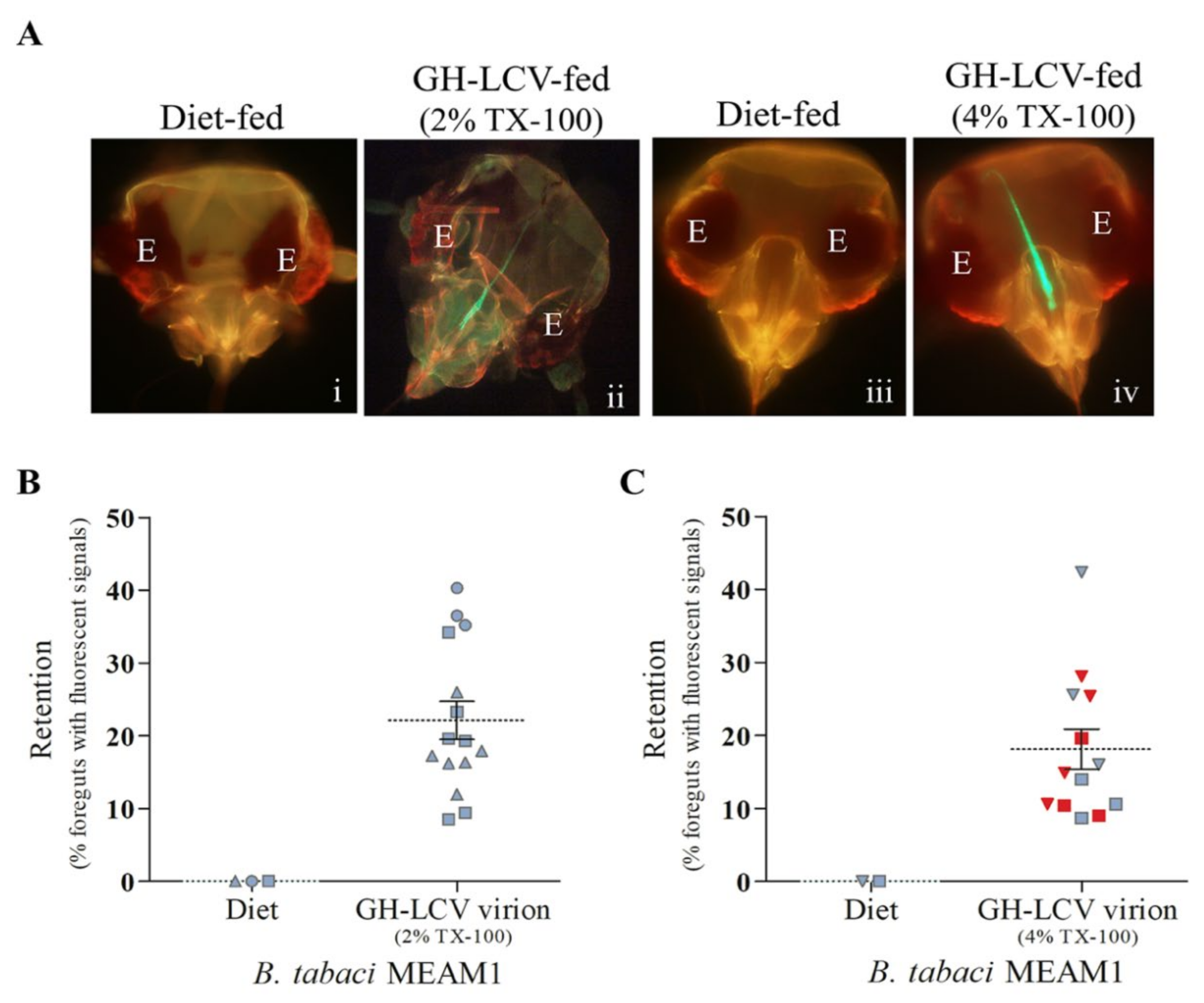
3.3. Purified GH-LCV Virions Are Retained in the Foreguts of B. tabaci MEAM1 Vectors, and Transmission Is Influenced by the TX-100 Concentration Used for Virion Purification
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Whitfield, A.E.; Falk, B.W.; Rotenberg, D. Insect vector-mediated transmission of plant viruses. Virology 2015, 479–480, 278–289. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.S.; Drucker, M.; Ng, J.C.K. Direct and indirect influences of virus-insect vector-plant interactions on non-circulative, semi-persistent virus transmission. Curr. Opin. Virol. 2018, 33, 129–136. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, S.; Michalakis, Y.; Van Munster, M.; Blanc, S. Plant feeding by insect vectors can affect life cycle, population genetics and evolution of plant viruses. Funct. Ecol. 2013, 27, 610–622. [Google Scholar] [CrossRef] [Green Version]
- Tzanetakis, I.E.; Martin, R.R.; Wintermantel, W.M. Epidemiology of criniviruses: An emerging problem in world agriculture. Front. Microbiol. 2013, 4, 119. [Google Scholar] [CrossRef] [Green Version]
- Wisler, G.C.; Duffus, J.E.; Liu, H.Y.; Li, R.H. Ecology and epidemiology of whitefly-transmitted Closteroviruses. Plant Dis. 1998, 82, 270–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klaassen, V.A.; Boeshore, M.L.; Koonin, E.V.; Tian, T.Y.; Falk, B.W. Genome structure and phylogenetic analysis of lettuce infectious yellows virus, a whitefly-transmitted, bipartite closterovirus. Virology 1995, 208, 99–110. [Google Scholar] [CrossRef] [Green Version]
- Salem, N.M.; Chen, A.Y.S.; Tzanetakis, I.E.; Mongkolsiriwattana, C.; Ng, J.C.K. Further complexity of the genus Crinivirus revealed by the complete genome sequence of Lettuce chlorosis virus (LCV) and the similar temporal accumulation of LCV genomic RNAs 1 and 2. Virology 2009, 390, 45–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Napuli, A.J.; Alzhanova, D.V.; Doneanu, C.E.; Barofsky, D.F.; Koonin, E.V.; Dolja, V.V. The 64-kilodalton capsid protein homolog of Beet yellows virus is required for assembly of virion tails. J. Virol. 2003, 77, 2377–2384. [Google Scholar] [CrossRef] [Green Version]
- Prokhnevsky, A.I.; Peremyslov, V.V.; Napuli, A.J.; Dolja, V.V. Interaction between long-distance transport factor and Hsp70-related movement protein of beet yellows virus. J. Virol. 2002, 76, 11003–11011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, A.Y.S.; Walker, G.P.; Carter, D.; Ng, J.C.K. A virus capsid component mediates virion retention and transmission by its insect vector. Proc. Natl. Acad. Sci. USA 2011, 108, 16777–16782. [Google Scholar] [CrossRef] [Green Version]
- Kubota, K.; Ng, J.C.K. Lettuce chlorosis virus P23 suppresses RNA silencing and induces local necrosis with increased severity at raised temperatures. Phytopathology 2016, 106, 653–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiss, Z.A.; Medina, V.; Falk, B.W. Crinivirus replication and host interactions. Front. Microbiol. 2013, 4, 99. [Google Scholar] [CrossRef] [Green Version]
- Qiao, W.; Medina, V.; Kuo, Y.W.; Falk, B.W. A distinct, non-virion plant virus movement protein encoded by a crinivirus essential for systemic infection. MBio 2018, 9, e02230-18. [Google Scholar] [CrossRef] [Green Version]
- Ng, J.C.K.; Falk, B.W. Virus-vector interactions mediating nonpersistent and semipersistent transmission of plant viruses. Annu. Rev. Phytopathol. 2006, 44, 183–212. [Google Scholar] [CrossRef]
- Ng, J.C.K.; Zhou, J.S. Insect vector-plant virus interactions associated with non-circulative, semi-persistent transmission: Current perspectives and future challenges. Curr. Opin. Virol. 2015, 15, 48–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, L.R.; Medina, V.; Tian, T.Y.; Turina, M.; Falk, B.W.; Ng, J.C.K. A mutation in the Lettuce infectious yellows virus minor coat protein disrupts whitefly transmission but not in planta systemic movement. J. Virol. 2010, 84, 12165–12173. [Google Scholar] [CrossRef] [Green Version]
- Tian, T.Y.; Rubio, L.; Yeh, H.H.; Crawford, B.; Falk, B.W. Lettuce infectious yellows virus: In vitro acquisition analysis using partially purified virions and the whitefly Bemisia tabaci. J. Gen. Virol. 1999, 80, 1111–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, J.C.K. A quantum dot-immunofluorescent labeling method to investigate the interactions between a crinivirus and Its whitefly Vector. Front. Microbiol. 2013, 4, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.S.; Chen, A.Y.S.; Drucker, M.; Lopez, N.H.; Carpenter, A.; Ng, J.C.K. Whitefly feeding behavior and retention of a foregut-borne crinivirus exposed to artificial diets with different pH values. Insect Sci. 2017, 24, 1079–1092. [Google Scholar] [CrossRef]
- Li, J.J.; Liang, X.Z.; Wang, X.L.; Shi, Y.; Gu, Q.S.; Kuo, Y.W.; Falk, B.W.; Yan, F.M. Direct evidence for the semipersistent transmission of Cucurbit chlorotic yellows virus by a whitefly vector. Sci. Rep. 2016, 6, 36604. [Google Scholar] [CrossRef] [Green Version]
- Ng, J.C.K.; Chen, A.Y.S. Acquisition of lettuce infectious yellows virus by Bemisia tabaci perturbs the transmission of lettuce chlorosis virus. Virus Res. 2011, 156, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.Y.S.; Pavitrin, A.; Ng, J.C.K. Agroinoculation of the cloned infectious cDNAs of Lettuce chlorosis virus results in systemic plant infection and production of whitefly transmissible virions. Virus Res. 2012, 169, 310–315. [Google Scholar] [CrossRef] [PubMed]
- McLain, J.; Castle, S.; Holmes, G.; Creamer, R. Physiochemical characterization and field assessment of lettuce chlorosis virus. Plant. Dis. 1998, 82, 1248–1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wisler, G.C.; Duffus, J.E. Transmission properties of whitefly-borne criniviruses and their impact on virus epidemiology. In Virus-Insect-Plant Interactions; Harris, K.F., Smith, O.P., Duffus, J.E., Eds.; Academic Press: San Diego, CA, USA, 2001; pp. 293–308. [Google Scholar]
- Sheveleva, A.A.; Nikitin, N.A.; Trifonova, E.A.; Zakubanskiy, A.V.; Chirkov, S.N. Improved method of purification of plum pox virus and serological analysis of the coat protein. Agric. Biol. 2016, 51, 385–391. [Google Scholar] [CrossRef] [Green Version]
- Van Oosten, H.J. Purification of plum pox (sharka) virus with the use of Triton X-100. Neth. J. Plant Pathol. 1972, 78, 33–44. [Google Scholar] [CrossRef]
- Cilia, M.; Peter, K.A.; Bereman, M.S.; Howe, K.; Fish, T.; Smith, D.; Gildow, F.; MacCoss, M.J.; Thannhauser, T.W.; Gray, S.M. Discovery and targeted LC-MS/MS of purified polerovirus reveals differences in the virus-host interactome associated with altered aphid transmission. PLoS ONE 2012, 7, e48177. [Google Scholar] [CrossRef]
- Nozu, Y.; Yamaura, R. Use of triton X-100 for isolation of tobacco mosaic virus from some plants other than tobacco. Virology 1971, 43, 514–515. [Google Scholar] [CrossRef]
- Ng, J.C.K.; Tian, T.; Falk, B.W. Quantitative parameters determining whitefly (Bemisia tabaci) transmission of Lettuce infectious yellows virus and an engineered defective RNA. J. Gen. Virol. 2004, 85 Pt 9, 2697–2707. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| 1 EXP | B. tabaci NW | EXP | B. tabaci MEAM1 | ||
|---|---|---|---|---|---|
| 2 Retention | 3 Transmission | Retention | Transmission | ||
| 1 | 5.7 ± 1.9 | 2/4 | 1 | 26.4 ± 3.0 | 3/6 |
| 2 | 23.8 ± 4.4 | 4/6 | 2 | 27.5 ± 4.37 | 4/6 |
| 3 | 18.2 ± 4.8 | 3/4 | 3 | 19.6 ± 2.74 | 4/5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, A.Y.S.; Zhou, J.S.; Liu, J.-X.; Ng, J.C.K. Nuances of Whitefly Vector–Crinivirus Interactions Revealed in the Foregut Retention and Transmission of Lettuce Chlorosis Virus by Two Bemisia tabaci Cryptic Species. Viruses 2021, 13, 1578. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081578
Chen AYS, Zhou JS, Liu J-X, Ng JCK. Nuances of Whitefly Vector–Crinivirus Interactions Revealed in the Foregut Retention and Transmission of Lettuce Chlorosis Virus by Two Bemisia tabaci Cryptic Species. Viruses. 2021; 13(8):1578. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081578
Chicago/Turabian StyleChen, Angel Y. S., Jaclyn S. Zhou, Jin-Xiang Liu, and James C. K. Ng. 2021. "Nuances of Whitefly Vector–Crinivirus Interactions Revealed in the Foregut Retention and Transmission of Lettuce Chlorosis Virus by Two Bemisia tabaci Cryptic Species" Viruses 13, no. 8: 1578. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081578