
Expansion of CD56dimCD16neg NK Cell Subset and Increased Inhibitory KIRs in Hospitalized COVID-19 Patients

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design, Participants and Methods
2.2. Cytokine Quantification
2.3. NK Cell Immunophenotyping
2.4. Statistical Analysis
3. Results
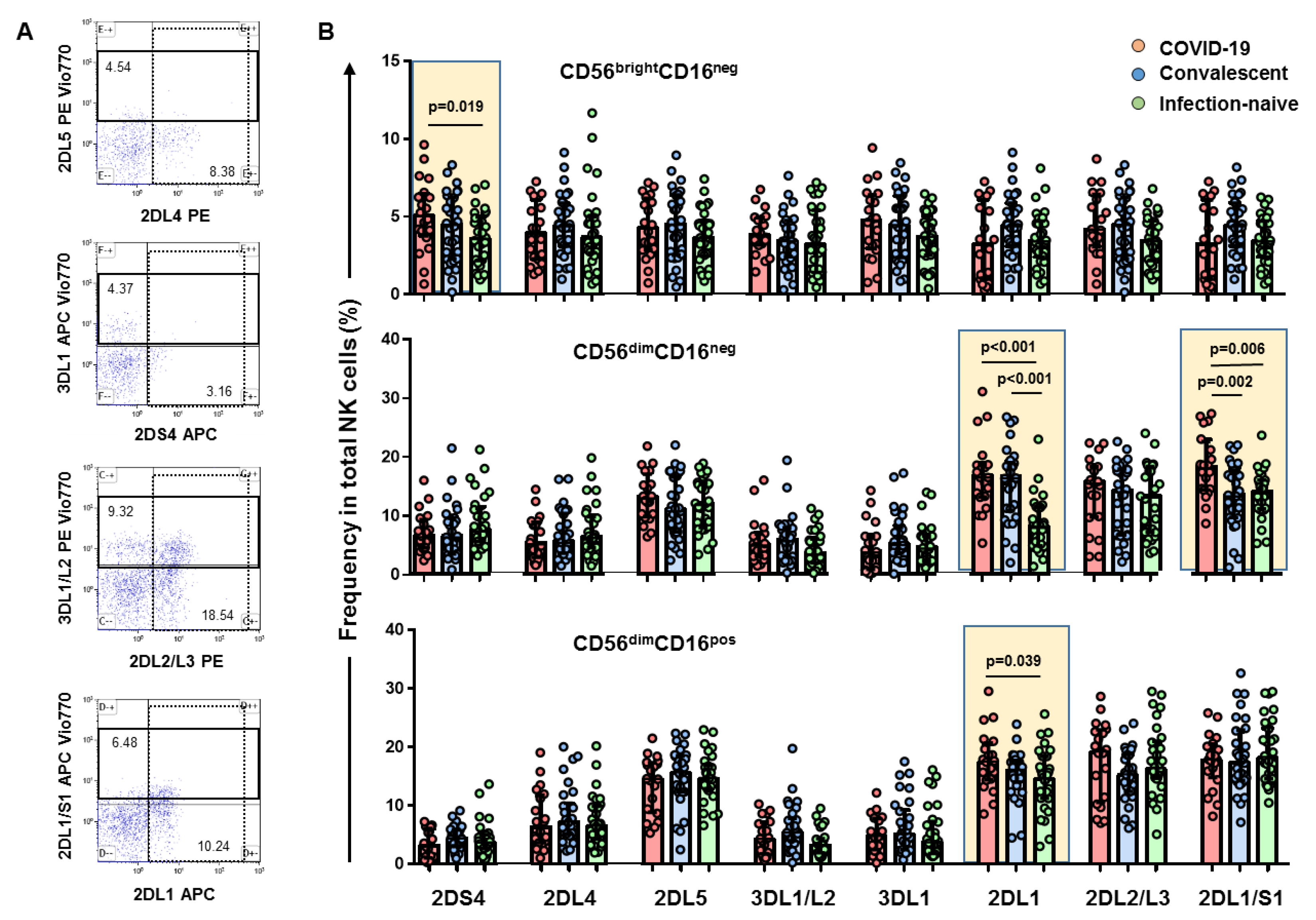
3.1. High Levels of CD56brightCD16neg NK Cell Subset Positive for KIR2DS4 Receptor in Hospitalized COVID-19 Patients
3.2. Expansion of CD56dimCD16neg NK Cell Subset and Higher Frequency of KIR2DL1 and KIR2DL1/S1 Inhibitor Receptors in Hospitalized COVID-19 Patients
3.3. Depletion of CD56dimCD16dim and CD56dimCD16bright NK Cell Subsets with a Higher Frequency of KIR2DL1 Inhibitory Receptor
3.4. NKs Cell Subset and KIRs Associations to Inflammatory Markers
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.-M.; Wang, W.; Song, Z.-G.; Hu, Y.; Tao, Z.-W.; Tian, J.-H.; Pei, Y.-Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A novel coronavirus from patients with pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef]
- World Health Organization. Weekly Epidemiological Update-19 October 2021. Available online: https://www.who.int/publications/m/item/weekly-epidemiologicalupdate-19-october-2021 (accessed on 19 October 2021).
- Guan, W.-J.; Ni, Z.-Y.; Hu, Y.; Liang, W.-H.; Ou, C.-Q.; He, J.-X.; Liu, L.; Shan, H.; Lei, C.-L.; Hui, D.S.C.; et al. Clinical characteristics of coronavirus disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef]
- Ferretti, A.P.; Tomasz, K.; Yifan, W.; Nguyen, D.M.V.; Adam, W.; Dunlap, G.S.; Qikai, X.; Nancy, N.; Perullo, C.R.; Cristofaro, A.W.; et al. Unbiased screens show CD8 T cells of COVID-19 patients recognize shared epitopes in SARS-CoV-2 that largely reside outside the spike protein. Immunity 2020, 53, 1095–1107.e3. [Google Scholar] [CrossRef]
- Alba, G.; Daniela, W.; Ramirez, S.I.; Jose, M.; Dan, J.M.; RydyznskiModerbacher, C.; Rawlings, S.A.; Aaron, S.; Lakshmanane, P.; Jadi, R.S.; et al. Targets of T cell responses to SARS-CoV-2 coronavirus in humans with COVID-19 disease and unexposed individuals. Cell 2020, 181, 1489–1501.e15. [Google Scholar]
- Abel, A.M.; Yang, C.; Thakar, M.S.; Malarkannan, S. Natural killer cells: Development, maturation, and clinical utilization. Front. Immunol. 2018, 9, 1869. [Google Scholar] [CrossRef] [Green Version]
- Andoniou, C.E.; Andrews, D.M.; Degli-Esposti, M.A. Natural killer cells in viral infection: More than just killers. Immunol. Rev. 2006, 214, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Vivier, E.; Tomasello, E.; Baratin, M.; Walzer, T.; Ugolini, S. Functions of natural killer cells. Nat. Immunol. 2008, 9, 503–510. [Google Scholar] [CrossRef]
- Moretta, A.; Bottino, C.; Vitale, M.; Pende, D.; Cantoni, C.; Mingari, M.C.; Biassoni, R.; Moretta, L. Activating receptors and coreceptors involved in human natural killer cell mediated cytolysis. Annu. Rev. Immunol. 2001, 19, 197–223. [Google Scholar] [CrossRef]
- Fan, Y.Y.; Yang, B.Y.; Wu, C.Y. Phenotypically and functionally distinct subsets of natural killer cells in human PBMCs. Cell Biol. Int. 2008, 32, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Amand, M.; Iserentant, G.; Poli, A.; Sleiman, M.; Fievez, V.; Sanchez, I.P.; Sauvageot, N.; Michel, T.; Aouali, N.; Janji, B.; et al. Human CD56(dim)CD16(dim) cells as an individualized natural killer cell subset. Front. Immunol. 2017, 8, 699. [Google Scholar] [CrossRef]
- Cao, X. COVID-19: Immunopathology and its implications for therapy. Nat. Rev. Immunol. 2020, 20, 269–270. [Google Scholar] [CrossRef] [Green Version]
- Masselli, E.; Vaccarezza, M.; Carubbi, C.; Pozzi, G.; Presta, V.; Mirandola, P.; Vitale, M. NK cells: A double edge sword against SARS-CoV-2. Adv. Biol. Regul. 2020, 77, 100737. [Google Scholar] [CrossRef]
- Forconi, C.S.; Oduor, C.I.; Oluoch, P.O.; Ong’echa, J.M.; Münz, C.; Bailey, J.A.; Moormann, A.M. A new hope for CD56negCD16pos NK cells as unconventional cytotoxic mediators: An adaptation to chronic diseases. Front. Cell Infect. Microbiol. 2020, 10, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soleimanian, S.; Yaghobi, R. Harnessing memory NK cell to protect against COVID-19. Front. Pharmacol. 2020, 11, 1309. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Zhou, L.; Hu, Z.; Zhang, S.; Yang, S.; Tao, Y.; Xie, C.; Ma, K.; Shang, K.; Wang, W.; et al. Dysregulation of immune response in patients with Coronavirus 2019 (COVID-19) in Wuhan, China. Clin. Infect. Dis. 2020, 71, 762–768. [Google Scholar] [CrossRef] [PubMed]
- Osman, M.; Faridi, R.M.; Sligl, W.; Shabani-Rad, M.-T.; Dharmani-Khan, P.; Parker, A.; Kalra, A.; Tripathi, M.B.; Storek, J.; Tervaert, J.W.C.; et al. Impaired natural killer cell counts and cytolytic activity in patients with severe COVID-19. Blood Adv. 2020, 4, 5035–5039. [Google Scholar] [CrossRef] [PubMed]
- Perez-Gomez, A.; Vitalle, J.; Gasca-Capote, M.C.; Gutierrez-Valencia, A.; Trujillo-Rodriguez, M.; Serna-Gallego, A.; Muñoz-Muela, E.; Jimenez-Leon, M.R.; Benhnia, M.R.; Rivas-Jeremias, I.; et al. Dendritic cell deficiencies persist seven months after SARS-CoV-2 infection. Cell Mol. Immunol. 2021, 18, 2128–2139. [Google Scholar] [CrossRef]
- Maucourant, C.; Filipovic, I.; Ponzetta, A.; Aleman, S.; Cornillet, M.; Hertwig, L.; Strunz, B.; Lentini, A.; Reinius, B.; Brownlie, D.; et al. Natural killer cell immunotypes related to COVID-19 disease severity. Sci. Immunol. 2020, 5, eabd6832. [Google Scholar] [CrossRef] [PubMed]
- Varchetta, S.; Mele, D.; Oliviero, B.; Mantovani, S.; Ludovisi, S.; Cerino, A.; Bruno, R.; Castelli, A.; Mosconi, M.; Vecchia, M.; et al. Unique immunological profile in patients with COVID-19. Cell Mol. Immunol. 2021, 18, 604–612. [Google Scholar] [CrossRef]
- Freud, A.G.; Caligiuri, M.A. Human natural killer cell development. Immunol. Rev. 2006, 214, 56–72. [Google Scholar] [CrossRef]
- Leem, G.; Cheon, S.; Lee, H.; Choi, J.S.; Jeong, S.; Kim, E.S.; Jeong, H.W.; Jeong, H.; Park, S.H.; Kim, Y.S.; et al. Abnormality in the NK-cell population is prolonged in severe COVID-19 patients. J. Allergy Clin. Immunol. 2021, 148, 996–1006.e18. [Google Scholar] [CrossRef]
- Stabile, H.; Nisti, P.; Morrone, S.; Pagliara, D.; Bertaina, A.; Locatelli, F.; Santoni, A.; Gismondi, A. Multifunctional human CD56 low CD16 low natural killer cells are the prominent subset in bone marrow of both healthy pediatric donors and leukemic patients. Haematologica 2015, 100, 489–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, B.M.; De Oliveira, M.H.S.; Benoit, S.; Plebani, M.; Lippi, G. Hematologic, biochemical and immune biomarker abnormalities associated with severe illness and mortality in coronavirus disease 2019 (COVID-19): A meta-analysis. Clin. Chem. Lab. Med. 2020, 58, 1021–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernal, E.; Gimeno, L.; Alcaraz, M.J.; Quadeer, A.A.; Moreno, M.; Martínez-Sánchez, M.V.; Campillo, J.A.; Gomez, J.M.; Pelaez, A.; García, E.; et al. Activating killer-cell immunoglobulin-like receptors are associated with the severity of Coronavirus Disease 2019. J. Infect. Dis. 2021, 224, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Dębska-Zielkowska, J.; Moszkowska, G.; Zieliński, M.; Zielińska, H.; Dukat-Mazurek, A.; Trzonkowski, P.; Stefańska, K. KIR receptors as key regulators of NK cells activity in health and disease. Cells 2021, 10, 1777. [Google Scholar] [CrossRef]
- Ruetsch, C.; Brglez, V.; Crémoni, M.; Zorzi, K.; Fernandez, C.; Boyer-Suavet, S.; Benzaken, S.; Demonchy, E.; Risso, K.; Courjon, J.; et al. Functional exhaustion of type I and II interferons production in severe COVID-19 patients. Front. Med. 2021, 7, 603961. [Google Scholar] [CrossRef] [PubMed]
- Osman, M.S.; van Eeden, C.; Tervaert, J.W.C. Fatal COVID-19 infections: Is NK cell dysfunction a link with autoimmune HLH? Autoimmun. Rev. 2020, 19, 102561. [Google Scholar] [CrossRef] [PubMed]
- Ekşioğlu-Demiralp, E.; Alan, S.; Sili, U.; Bakan, D.; Ocak, İ.; Yürekli, R.; Alpay, N.; Görçin, S.; Yıldız, A. Peripheral innate and adaptive immune cells during COVID-19: Functional neutrophils, pro-inflammatory monocytes, and half-dead lymphocytes. Cytom. B Clin. Cytom. 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- Bergantini, L.; d’Alessandro, M.; Cameli, P.; Cavallaro, D.; Gangi, S.; Cekorja, B.; Sestini, P.; Bargagli, E. NK and T Cell Immunological Signatures in Hospitalized Patients with COVID-19. Cells 2021, 10, 3182. [Google Scholar] [CrossRef]
- Penack, O.; Gentilini, C.; Fischer, L.; Asemissen, A.M.; Scheibenbogen, C.; Thiel, E.; Uharek, L. CD56dimCD16neg cells are responsible for natural cytotoxicity against tumor targets. Leukemia 2005, 19, 835–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Björkström, N.K.; Ponzetta, A. Natural killer cells and unconventional T cells in COVID-19. Curr. Opin. Virol. 2021, 49, 176–182. [Google Scholar] [CrossRef]
- Romee, R.; Foley, B.; Lenvik, T.; Wang, Y.; Zhang, B.; Ankarlo, D.; Luo, X.; Cooley, S.; Verneris, M.; Walcheck, B.; et al. NK cell CD16 surface expression and function is regulated by a disintegrin and metalloprotease-17 (ADAM17). Blood 2013, 121, 3599–3608. [Google Scholar] [CrossRef]
- Littera, R.; Chessa, L.; Deidda, S.; Angioni, G.; Campagna, M.; Lai, S.; Melis, M.; Cipri, S.; Firinu, D.; Santus, S.; et al. Natural killer-cell immunoglobulin-like receptors trigger differences in immune response to SARS-CoV-2 infection. PLoS ONE 2020, 16, e0255608. [Google Scholar] [CrossRef]
- Hsieh, W.C.; Lai, E.Y.; Liu, Y.T.; Wang, Y.F.; Tzeng, Y.S.; Cui, L.; Lai, Y.J.; Huang, H.C.; Huang, J.H.; Ni, H.C.; et al. NK cell receptor and ligand composition influences the clearance of SARS-CoV-2. J. Clin. Investig. 2021, 131, e146408. [Google Scholar] [CrossRef]
- Narni-Mancinelli, E.; Vivier, E. Clues that natural killer cells help to control COVID. Nature 2021, 600, 226–227. [Google Scholar] [CrossRef]
- Mazzoni, A.; Salvati, L.; Maggi, L.; Capone, M.; Vanni, A.; Spinicci, M.; Mencarini, J.; Caporale, R.; Peruzzi, B.; Antonelli, A.; et al. Impaired immune cell cytotoxicity in severe COVID-19 is IL-6 dependent. J. Clin. Investig. 2020, 130, 4694–4703. [Google Scholar] [CrossRef] [PubMed]
- Ghasemzadeh, M.; Ghasemzadeh, A.; Hosseini, E. Exhausted NK cells and cytokine storms in COVID-19: Whether NK cell therapy could be a therapeutic choice. Hum. Immunol. Ahead of Print. 2021. [Google Scholar] [CrossRef] [PubMed]
- Lenart, M.; Kluczewska, A.; Szaflarska, A.; Rutkowska-Zapała, M.; Wąsik, M.; Ziemiańska-Pięta, A.; Kobylarz, K.; Pituch-Noworolska, A.; Siedlar, M. Selective downregulation of natural killer activating receptors on NK cells and upregulation of PD-1 expression on T cells in children with severe and/or recurrent Herpes simplex virus infections. Immunobiology 2021, 226, 152097. [Google Scholar] [CrossRef] [PubMed]
- Campbell, T.M.; McSharry, B.P.; Steain, M.; Ashhurst, T.M.; Slobedman, B.; Abendroth, A.; Mocarski, E. Varicella zoster virus productively infects human natural killer cells and manipulates phenotype. PLoS Pathog. 2018, 14, e1006999. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hospitalized COVID-19 n = 20 | Convalescent n = 30 | Infection Naive n = 30 | |
|---|---|---|---|
| Age (years) | 60 (52–72) | 45 (31–54) | 40 (33–50) |
| Sex (Female n, %) | 1 (5%) | 18 (60%) | 19 (65.5%) |
| Body Mass Index (Kg/m2) | 29.1 (24.9–31.1) | 23.5 (22.0–24.6) | 23.6 (20.8–26.0) |
| Obesity (>30 BMI) | 7 (35%) | 1 (3.3%) | 1 (3.3%) |
| Comorbidities (n, %) | |||
| Hypertension | 7 (35%) | 3 (10%) | 3 (10%) |
| Diabetes | 6 (30%) | 1 (3.5%) | 1 (3.5%) |
| History of positive SARS-CoV-2 RT-PCR | 17 (85%) * | 19 (63.3%) ** | - |
| History of positive anti-SARS-CoV-2 IgA/IgM/IgG n-specific antibodies | 20 (100%) * | 30 (100%) | 0 |
| History of SARS-CoV-2-compatible symptoms | 20 (100%) * | 19 (63.3%) | 0 |
| SARS-CoV-2 IgG S-specific antibodies at inclusion | 16 (80%) | 30 (100%) | 0 |
| Sepsis-related organ failure assessment (SOFA) score | |||
| 1 | 15 (75%) | - | - |
| 2–3 | 5 (25%) | - | - |
| Time since SARS-CoV-2 diagnosis (days) | 7 (5–9) | 60 (26–75) | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Casado, J.L.; Moraga, E.; Vizcarra, P.; Velasco, H.; Martín-Hondarza, A.; Haemmerle, J.; Gómez, S.; Quereda, C.; Vallejo, A. Expansion of CD56dimCD16neg NK Cell Subset and Increased Inhibitory KIRs in Hospitalized COVID-19 Patients. Viruses 2022, 14, 46. https://0-doi-org.brum.beds.ac.uk/10.3390/v14010046
Casado JL, Moraga E, Vizcarra P, Velasco H, Martín-Hondarza A, Haemmerle J, Gómez S, Quereda C, Vallejo A. Expansion of CD56dimCD16neg NK Cell Subset and Increased Inhibitory KIRs in Hospitalized COVID-19 Patients. Viruses. 2022; 14(1):46. https://0-doi-org.brum.beds.ac.uk/10.3390/v14010046
Chicago/Turabian StyleCasado, José L., Elisa Moraga, Pilar Vizcarra, Héctor Velasco, Adrián Martín-Hondarza, Johannes Haemmerle, Sandra Gómez, Carmen Quereda, and Alejandro Vallejo. 2022. "Expansion of CD56dimCD16neg NK Cell Subset and Increased Inhibitory KIRs in Hospitalized COVID-19 Patients" Viruses 14, no. 1: 46. https://0-doi-org.brum.beds.ac.uk/10.3390/v14010046