
Molecular Characterization and Pathogenicity of a Novel Soybean-Infecting Monopartite Geminivirus in China

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Small RNA-Based Deep Sequencing
2.3. DNA Extraction and Amplification of the Full-Length Viral Genome
2.4. Genome Characterization
2.5. Pairwise Distances, Phylogenetic, and Recombination Analysis
2.6. Infectivity Assays
2.7. Agroinoculation of Plants
2.8. Southern Blot Assay
3. Results
3.1. Discovery of a Novel Soybean-Infecting Monopartite Geminivirus
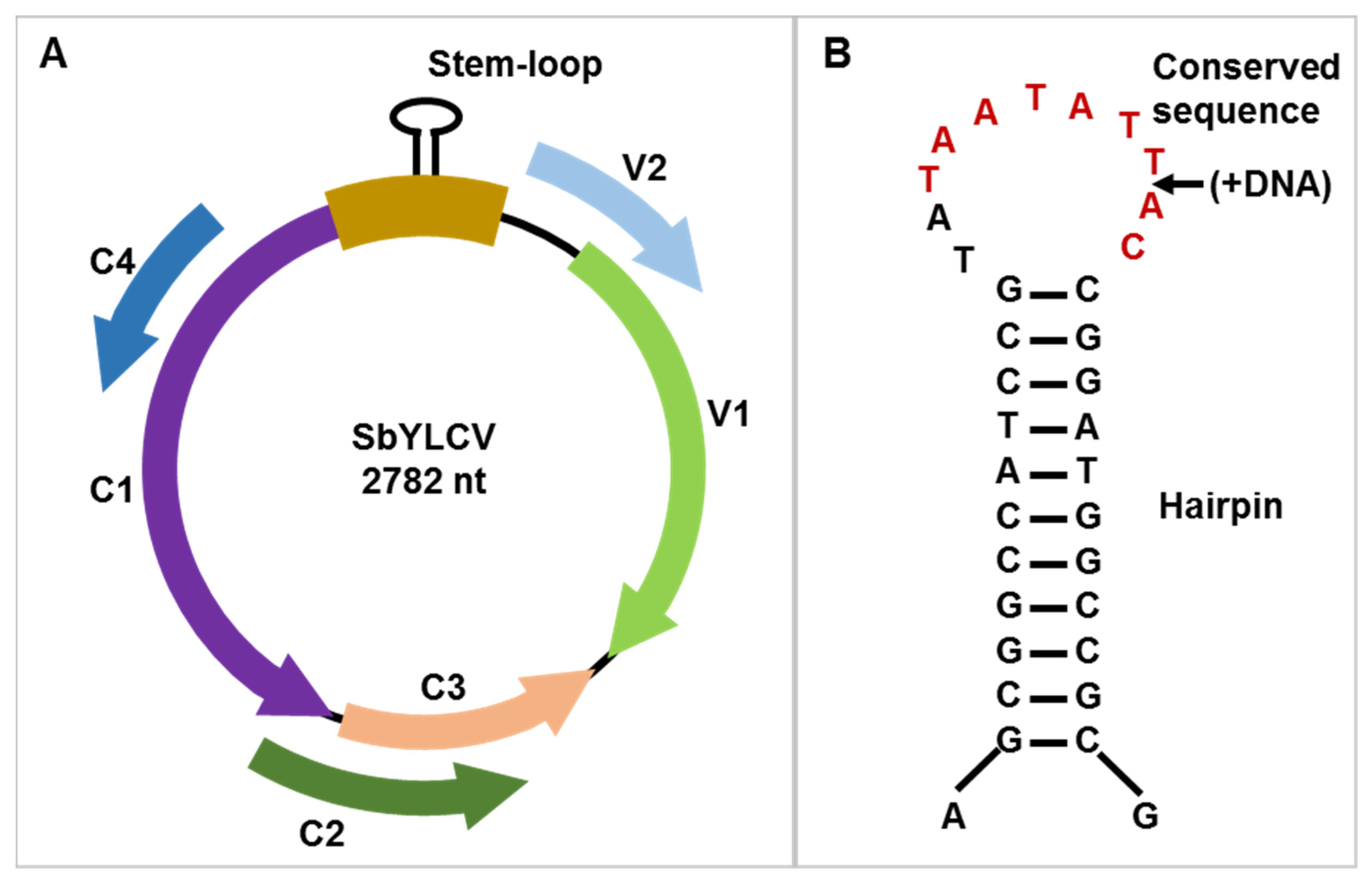
3.2. Genome Organization of SbYLCV
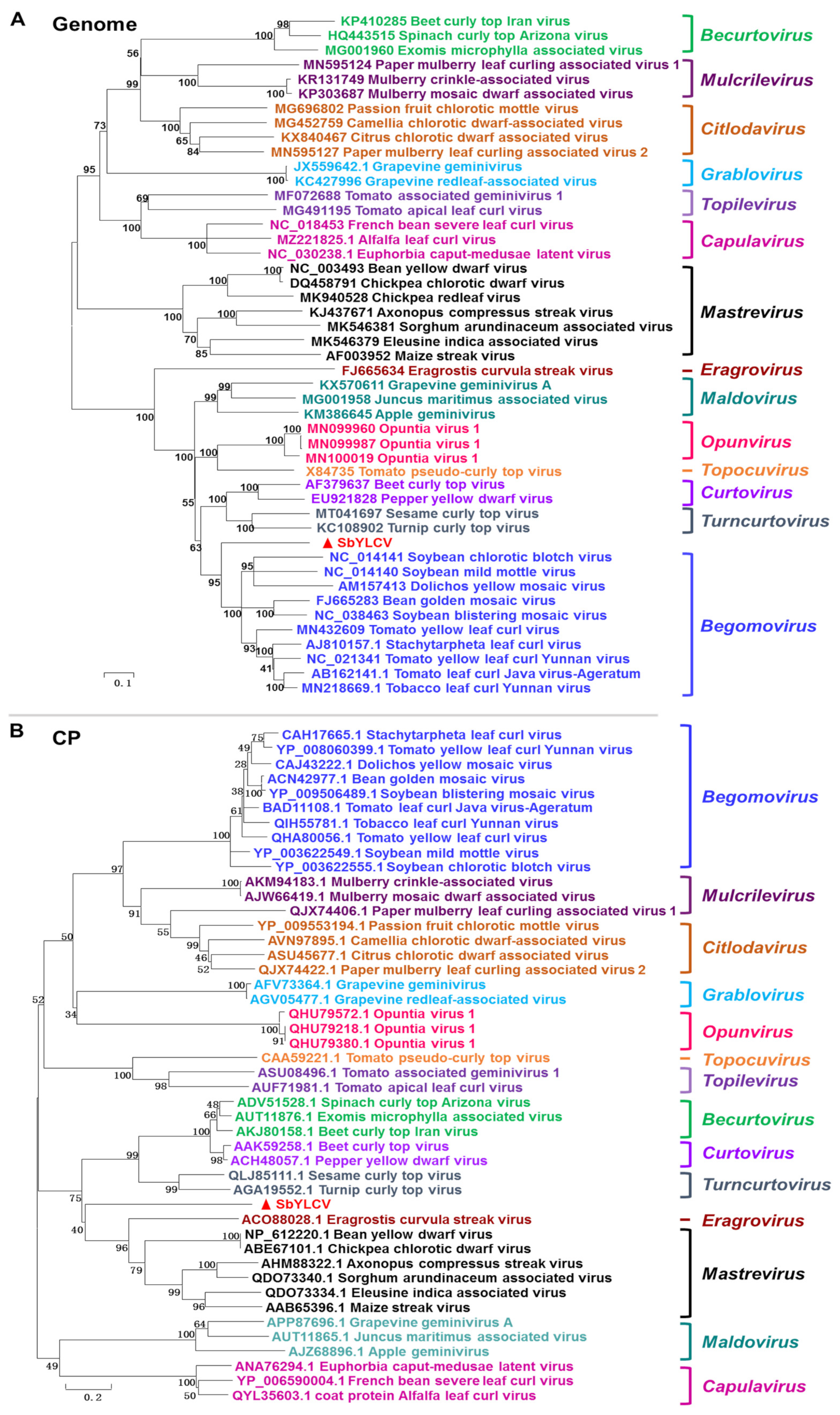
3.3. Phylogenetic Relationships
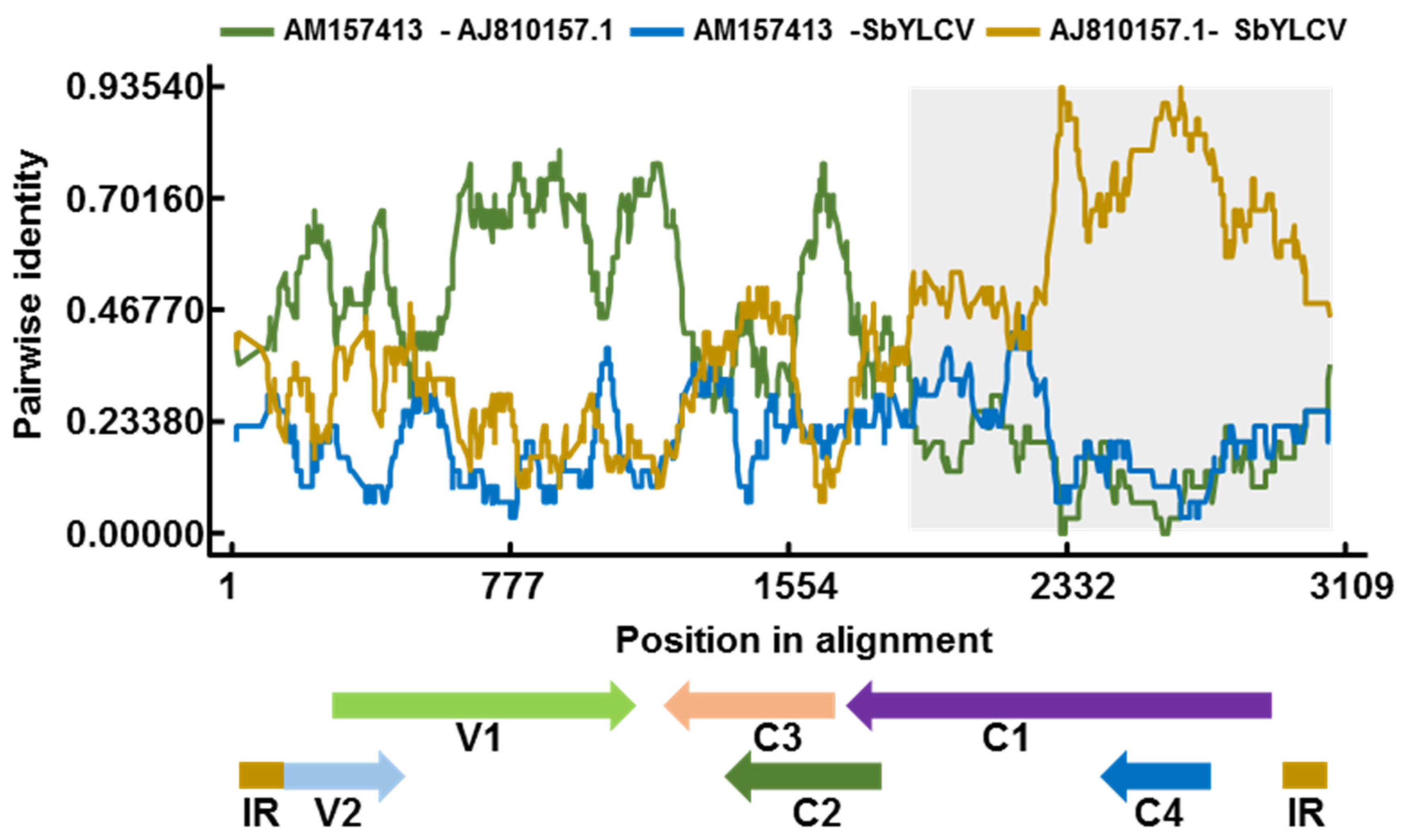
3.4. Recombination Analysis
3.5. Infectivity of the Infectious Clone of SbYLCV
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, X.; Guo, W.; Li, F.; Sunter, G.; Zhou, X. Geminivirus-associated betasatellites: Exploiting chinks in the antiviral arsenal of plants. Trends Plant Sci. 2019, 24, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Rojas, M.R.; Macedo, M.A.; Maliano, M.R.; Soto-Aguilar, M.; Souza, J.O.; Briddon, R.W.; Kenyon, L.; Rivera Bustamante, R.F.; Zerbini, F.M.; Adkins, S.; et al. World management of geminiviruses. Annu. Rev. Phytopathol. 2018, 56, 637–677. [Google Scholar] [CrossRef] [PubMed]
- Vaghi Medina, C.G.; Teppa, E.; Bornancini, V.A.; Flores, C.R.; Marino-Buslje, C.; Lopez Lambertini, P.M. Tomato apical leaf curl virus: A novel, monopartite geminivirus detected in tomatoes in Argentina. Front. Microbiol. 2017, 8, 2665. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Navarro, B.; Zhang, Z.; Lu, M.; Zhou, X.; Chi, S.; Di Serio, F.; Li, S. Identification and molecular characterization of a novel monopartite geminivirus associated with mulberry mosaic dwarf disease. J. Gen. Virol. 2015, 96, 2421–2434. [Google Scholar] [CrossRef]
- Liang, P.; Navarro, B.; Zhang, Z.; Wang, H.; Lu, M.; Xiao, H.; Wu, Q.; Zhou, X.; Di Serio, F.; Li, S. Identification and characterization of a novel geminivirus with a monopartite genome infecting apple trees. J. Gen. Virol. 2015, 96, 2411–2420. [Google Scholar] [CrossRef]
- Fontenele, R.S.; Abreu, R.A.; Lamas, N.S.; Alves-Freitas, D.M.T.; Vidal, A.H.; Poppiel, R.R.; Melo, F.L.; Lacorte, C.; Martin, D.P.; Campos, M.A.; et al. Passion fruit chlorotic mottle virus: Molecular characterization of a new divergent geminivirus in Brazil. Viruses 2018, 10, 169. [Google Scholar] [CrossRef] [Green Version]
- Zerbini, F.M.; Briddon, R.W.; Idris, A.; Martin, D.P.; Moriones, E.; Navas-Castillo, J.; Rivera-Bustamante, R.; Roumagnac, P.; Varsani, A.; ICTV Report, C. ICTV virus taxonomy profile: Geminiviridae. J. Gen. Virol. 2017, 98, 131–133. [Google Scholar] [CrossRef]
- Fiallo-Olivé, E.; Lett, J.M.; Martin, D.P.; Roumagnac, P.; Varsani, A.; Zerbini, F.M.; Navas-Castillo, J. ICTV Virus Taxonomy Profile: Geminiviridae 2021. J. Gen. Virol. 2021, 102, 001696. [Google Scholar] [CrossRef]
- Nawaz-ul-Rehman, M.S.; Fauquet, C.M. Evolution of geminiviruses and their satellites. FEBS Lett. 2009, 583, 1825–1832. [Google Scholar] [CrossRef] [Green Version]
- Strange, R.N.; Scott, P.R. Plant disease: A threat to global food security. Annu. Rev. Phytopathol. 2005, 43, 83–116. [Google Scholar] [CrossRef]
- Hill, J.H.; Whitham, S.A. Control of virus diseases in soybeans. Adv. Virus Res. 2014, 90, 355–390. [Google Scholar] [PubMed]
- Yang, X.; Du, M.; Li, S.; Zhou, X. Coinfection of cotton plants with watermelon mosaic virus and a novel polerovirus in China. Viruses 2021, 13, 2210. [Google Scholar] [CrossRef] [PubMed]
- Zerbino, D.R.; Birney, E. Velvet: Algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef] [Green Version]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, e108277. [Google Scholar]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Martin, D.P.; Lemey, P.; Lott, M.; Moulton, V.; Posada, D.; Lefeuvre, P. RDP3: A flexible and fast computer program for analyzing recombination. Bioinformatics 2010, 26, 2462–2463. [Google Scholar] [CrossRef]
- Ma, Z.; Zhang, H.; Ding, M.; Zhang, Z.; Yang, X.; Zhou, X. Molecular characterization and pathogenicity of an infectious cDNA clone of tomato brown rugose fruit virus. Phytopathol. Res. 2021, 3, 14. [Google Scholar] [CrossRef]
- Yang, X.; Ren, Y.; Sun, S.; Wang, D.; Zhang, F.; Li, D.; Li, S.; Zhou, X. Identification of the potential virulence factors and RNA silencing suppressors of mulberry mosaic dwarf-associated geminivirus. Viruses 2018, 10, 472. [Google Scholar] [CrossRef] [Green Version]
- Deng, D.; McGrath, P.F.; Robinson, D.J.; Harrison, B.D. Detection and differentiation of whitefly-transmitted geminiviruses in plants and vector insects by the polymerase chain-reaction with degenerate primers. Ann. Appl. Biol. 1994, 125, 327–336. [Google Scholar] [CrossRef]
- Bull, S.E.; Briddon, R.W.; Markham, P.G. Universal primers for the PCR-mediated amplification of DNA 1: A satellite-like molecule associated with begomovirus-DNAß complexes. Mol. Biotechnol. 2003, 20, 83–86. [Google Scholar] [CrossRef]
- Briddon, R.W.; Bull, S.E.; Mansoor, S.; Amin, I.; Markham, P.G. Universal primers for the PCR-mediated amplification of DNAß. Mol. Biotechnol. 2002, 20, 315–318. [Google Scholar] [CrossRef]
- Ilyina, T.V.; Koonin, E.V. Conserved sequence motifs in the initiator proteins for rolling circle DNA replication encoded by diverse replicons from eubacteria, eucaryotes, and archaebacteria. Nucleic Acids Res. 1992, 20, 3279–3285. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V.; Ilyina, T.V. Geminivirus replication proteins are related to prokaryotic plasmid rolling circle DNA replication initiator proteins. J. Gen. Virol. 1992, 73, 2763–2766. [Google Scholar] [CrossRef] [PubMed]
- Nash, T.E.; Dallas, M.B.; Reyes, M.I.; Buhrman, G.K.; Ascencio-Ibanez, J.T.; Hanley-Bowdoin, L. Functional analysis of a novel motif conserved across geminivirus Rep proteins. J. Virol. 2011, 85, 1182–1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefeuvre, P.; Moriones, E. Recombination as a motor of host switches and virus emergence: Geminiviruses as case studies. Curr. Opin. Virol. 2015, 10, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Zhao, L.; Jiao, X.; Jiang, T.; Gong, H.; Wang, B.; Briddon, R.W.; Zhou, X. A recombinant begomovirus resulting from exchange of the C4 gene. J. Gen. Virol. 2013, 94, 1896–1907. [Google Scholar] [CrossRef]
- Tao, X.; Zhou, X. A modified viral satellite DNA that suppresses gene expression in plants. Plant J. Cell Mol. Biol. 2004, 38, 850–860. [Google Scholar] [CrossRef]
- Xu, X.; Qian, Y.; Wang, Y.; Li, Z.; Zhou, X. Iterons homologous to helper geminiviruses are essential for efficient replication of betasatellites. J. Virol. 2019, 93, e01532-18. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X. Advances in understanding begomovirus satellites. Annu. Rev. Phytopathol. 2013, 51, 357–381. [Google Scholar] [CrossRef]
- Luna, A.P.; Lozano-Duran, R. Geminivirus-encoded proteins: Not all positional homologs are made equal. Front. Microbiol. 2020, 11, 878. [Google Scholar] [CrossRef]
- Zhou, X. Hijack to escape: A geminivirus seizes a host imprinted E3 ligase to escape epigenetic repression. Sci. China Life Sci. 2021, 64, 323–325. [Google Scholar] [CrossRef] [PubMed]
- Medina-Puche, L.; Orilio, A.F.; Zerbini, F.M.; Lozano-Duran, R. Small but mighty: Functional landscape of the versatile geminivirus-encoded C4 protein. PLoS Pathog. 2021, 17, e1009915. [Google Scholar] [CrossRef]
- Gong, P.; Tan, H.; Zhao, S.; Li, H.; Liu, H.; Ma, Y.; Zhang, X.; Rong, J.; Fu, X.; Lozano-Duran, R.; et al. Geminiviruses encode additional small proteins with specific subcellular localizations and virulence function. Nat. Commun. 2021, 12, 4278. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Song, Y.; Wang, Y.; Zhou, X. Functional analysis of a novel ßV1 gene identified in a geminivirus betasatellite. Sci. China Life Sci. 2020, 63, 688–696. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ORF | Nucleotide Coordinates | Orientation | No. Amino Acids | Mr (kDa) | BLASTP (E-Value) | Predicted Domains (SMART) | Amino Acid Coordinates of Predicted Domain |
|---|---|---|---|---|---|---|---|
| V1 | 263–1048 | Sense | 262 | 29.4 | Coat protein of sweet potato symptomless virus 1 | Geminivirus coat protein/nuclear export factor BR1 family (1.04E-10) | 5–259 |
| V2 | 139–450 | Sense | 129 | 11.9 | V2 of Juncus maritimus associated virus | - | - |
| C1 | 2637–1552 | Complementary | 362 | 41.3 | Rep of ageratum yellow vein China virus | Geminivirus Rep catalytic domain (3.04E-62) | 7–119 |
| Geminivirus rep protein central domain (5.73E-35) | 126–230 | ||||||
| C2 | 1649–1233 | Complementary | 139 | 15.4 | C2 of Juncus maritimus associated virus | Geminivirus AL2 protein (3.18E-20) | 1–135 |
| C3 | 1537–1082 | Complementary | 152 | 17.6 | C3 of Juncus maritimus associated virus | Geminivirus AL3 protein (4.13E-35) | 18–132 |
| C4 | 2480–2190 | Complementary | 97 | 10.5 | C4 of ageratum yellow vein virus | Geminivirus C4 protein (1.72E-35) | 1–84 |
| Plants | No. of Plants Infiltrated | No. of Plants Systemically Infected | Infection Efficiency (%) | Symptoms |
|---|---|---|---|---|
| Nicotiana benthamiana | 34 | 34 | 100% | Leaf curling |
| Glycine max | 52 | 5 | 9.6% | Leaf curling, yellowing |
| Solanum lycopersicum | 15 | 0 | 0 | NA |
| Nicotiana tabacum | 16 | 13 | 81.3% | Leaf curling |
| Nicotiana tabacum cv. Samsun NN | 10 | 7 | 70% | Leaf curling |
| Nicotiana tabacum cv. Samsun nn | 10 | 3 | 33% | Leaf curling |
| Nicotiana glutinosa | 10 | 2 | 20% | Leaf curling |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, M.; Wang, Y.; Chen, C.; Li, X.; Feng, R.; Zhou, X.; Yang, X. Molecular Characterization and Pathogenicity of a Novel Soybean-Infecting Monopartite Geminivirus in China. Viruses 2022, 14, 341. https://0-doi-org.brum.beds.ac.uk/10.3390/v14020341
Du M, Wang Y, Chen C, Li X, Feng R, Zhou X, Yang X. Molecular Characterization and Pathogenicity of a Novel Soybean-Infecting Monopartite Geminivirus in China. Viruses. 2022; 14(2):341. https://0-doi-org.brum.beds.ac.uk/10.3390/v14020341
Chicago/Turabian StyleDu, Min, Yongzhi Wang, Cheng Chen, Xiaoyu Li, Runzi Feng, Xueping Zhou, and Xiuling Yang. 2022. "Molecular Characterization and Pathogenicity of a Novel Soybean-Infecting Monopartite Geminivirus in China" Viruses 14, no. 2: 341. https://0-doi-org.brum.beds.ac.uk/10.3390/v14020341