
Oropharyngeal Shedding of Gammaherpesvirus DNA by Cats, and Natural Infection of Salivary Epithelium

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Samples
2.3. DNA Extraction and PCR Testing
2.4. In Situ Hybridization
2.5. Statistical Analysis
3. Results
3.1. Oropharyngeal Shedding of FcaGHV-1
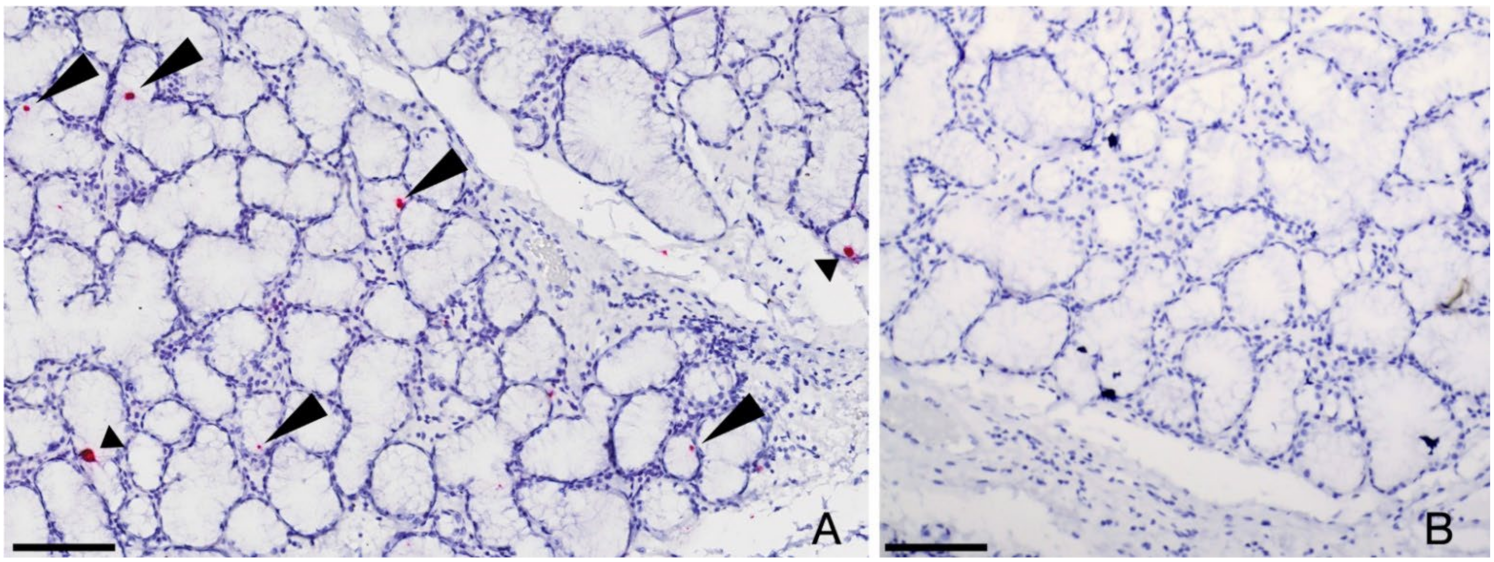
3.2. Persistence of FcaGHV-1 in Salivary Gland Epithelium
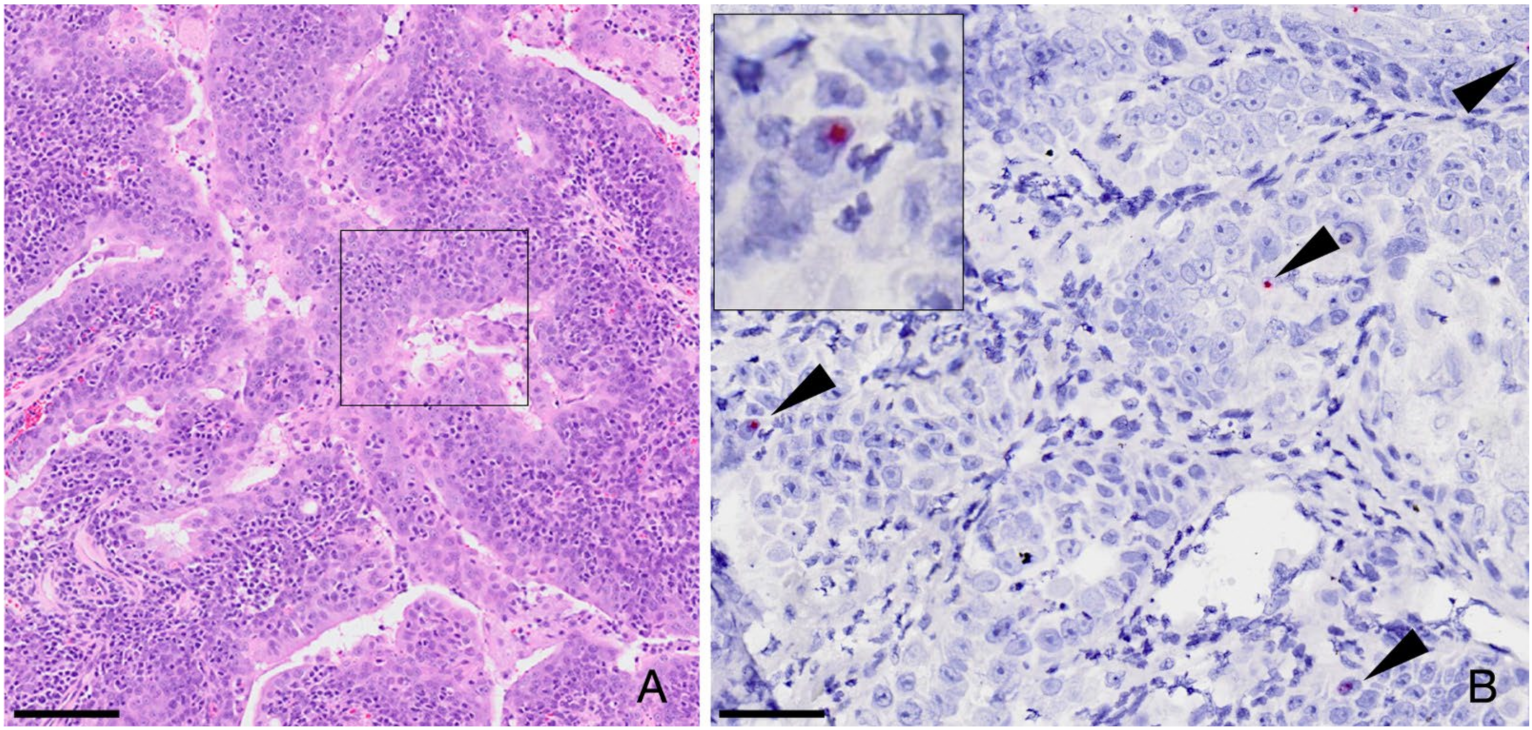
3.3. FcaGHV1 Sequence Not Detected in Neoplastic Cells from Oronasal Carcinomas
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Young, L.S.; Yap, L.F.; Murray, P.G. Epstein–Barr virus: More than 50 years old and still providing surprises. Nat. Rev. Cancer 2016, 16, 789–802. [Google Scholar] [CrossRef] [PubMed]
- Carbone, A.; Volpi, C.C.; Gualeni, A.V.; Gloghini, A. Epstein-Barr virus associated lymphomas in people with HIV. Curr. Opin. HIV AIDS 2017, 12, 39–46. [Google Scholar] [CrossRef]
- Li, H.; Cunha, C.W.; Taus, N.S.; Knowles, D.P. Malignant catarrhal fever: Inching toward understanding. Annu. Rev. Anim. Biosci. 2014, 2, 209–233. [Google Scholar] [CrossRef] [PubMed]
- Troyer, R.M.; Beatty, J.A.; Stutzman-Rodriguez, K.R.; Carver, S.; Lozano, C.C.; Lee, J.S.; Lappin, M.R.; Riley, S.P.D.; Serieys, L.E.K.; Logan, K.A.; et al. Novel Gammaherpesviruses in North American Domestic Cats, Bobcats, and Pumas: Identification, Prevalence, and Risk Factors. J. Virol. 2014, 88, 3914–3924. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Pérez, P.; Hyndman, T.H.; Fleming, P.A.; Vaz, P.K.; Ficorilli, N.P.; Wilks, C.R. A Widespread Novel Gammaherpesvirus in Apparently Healthy Wild Quokkas (Setonix brachyurus): A Threatened and Endemic Wallaby of Western Australia. J. Zoo Wildl. Med. 2021, 52, 592–603. [Google Scholar] [CrossRef]
- Lozano, C.C.; Sweanor, L.L.; Wilson-Henjum, G.; Kays, R.W.; Moreno, R.; VandeWoude, S.; Troyer, R.M. Identification of Novel Gammaherpesviruses in Ocelots (Leopardus pardalis) and Bobcats (Lynx rufus) in Panama and Colorado, USA. J. Wildl. Dis. 2015, 51, 911–915. [Google Scholar] [CrossRef]
- Nicolas de Francisco, O.; Esperón, F.; Juan-Sallés, C.; Ewbank, A.C.; das Neves, C.G.; Marco, A.; Neves, E.; Anderson, N.; Sacristán, C. Neoplasms and novel gammaherpesviruses in critically endangered captive European minks (Mustela lutreola). Transbound. Emerg. Dis. 2021, 68, 552–564. [Google Scholar] [CrossRef]
- McLuckie, A.; Tasker, S.; Dhand, N.K.; Spencer, S.; Beatty, J.A. High prevalence of Felis catus gammaherpesvirus 1 infection in haemoplasma-infected cats supports co-transmission. Vet. J. 2016, 214, 117–121. [Google Scholar] [CrossRef] [Green Version]
- Caringella, F.; Desario, C.; Lorusso, E.; Pallante, I.; Furlanello, T.; Lanave, G.; Elia, G.; Martella, V.; Iatta, R.; Barrs, V.R.; et al. Prevalence and risk factors for Felis catus gammaherpesvirus 1 detection in domestic cats in Italy. Vet. Microbiol. 2019, 238, 108426. [Google Scholar] [CrossRef]
- Novacco, M.; Ranjbar Kohan, N.; Stirn, M.; Meli, M.L.; Díaz-Sánchez, A.A.; Boretti, F.S.; Hofmann-Lehmann, R. Prevalence, Geographic Distribution, Risk Factors and Co-Infections of Feline Gammaherpesvirus Infections in Domestic Cats in Switzerland. Viruses 2019, 11, 721. [Google Scholar] [CrossRef] [Green Version]
- Beatty, J.A.; Troyer, R.M.; Carver, S.; Barrs, V.R.; Espinasse, F.; Conradi, O.; Stutzman-Rodriguez, K.; Chan, C.C.; Tasker, S.; Lappin, M.R.; et al. Felis catus gammaherpesvirus 1; a widely endemic potential pathogen of domestic cats. Virology 2014, 460, 100–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ertl, R.; Korb, M.; Langbein-Detsch, I.; Klein, D. Prevalence and risk factors of gammaherpesvirus infection in domestic cats in Central Europe. Virol. J. 2015, 12, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurissio, J.K.; Rodrigues, M.V.; Taniwaki, S.A.; Zanutto, M.D.; Filoni, C.; Galdino, M.V.; Araujo, J.P. Felis catus gammaherpesvirus 1 (FcaGHV1) and coinfections with feline viral pathogens in domestic cats in Brazil. Cienc. Rural 2018, 48, e20170480. [Google Scholar] [CrossRef] [Green Version]
- Stutzman-Rodriguez, K.; Rovnak, J.; VandeWoude, S.; Troyer, R.M. Domestic cats seropositive for Felis catus gammaherpesvirus 1 are often qPCR negative. Virology 2016, 498, 23–30. [Google Scholar] [CrossRef]
- Hadinoto, V.; Shapiro, M.; Sun, C.C.; Thorley-Lawson, D.A. The dynamics of EBV shedding implicate a central role for epithelial cells in amplifying viral output. PLoS Pathog. 2009, 5, e1000496. [Google Scholar] [CrossRef] [Green Version]
- Tsao, S.W.; Tsang, C.M.; Lo, K.W. Epstein-Barr virus infection and nasopharyngeal carcinoma. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160270. [Google Scholar] [CrossRef]
- Hwang, S.; Wu, T.-T.; Tong, L.M.; Kim, K.S.; Martinez-Guzman, D.; Colantonio, A.D.; Uittenbogaart, C.H.; Sun, R. Persistent Gammaherpesvirus Replication and Dynamic Interaction with the Host In Vivo. J. Virol. 2008, 82, 12498–12509. [Google Scholar] [CrossRef] [Green Version]
- Ambrosio, M.R.; Mastrogiulio, M.G.; Barone, A.; Rocca, B.J.; Gallo, C.; Lazzi, S.; Leoncini, L.; Bellan, C. Lymphoepithelial-like carcinoma of the parotid gland: A case report and a brief review of the western literature. Diagn. Pathol. 2013, 8, 115. [Google Scholar] [CrossRef] [Green Version]
- Pathmanathan, R.; Prasad, U.; Sadler, R.; Flynn, K.; Raab-Traub, N. Clonal proliferations of cells infected with Epstein-Barr virus in preinvasive lesions related to nasopharyngeal carcinoma. N. Engl. J. Med. 1995, 333, 693–698. [Google Scholar] [CrossRef]
- McLuckie, A.; Barrs, V.; Wilson, B.; Westman, M.; Beatty, J. Felis Catus Gammaherpesvirus 1 DNAemia in Whole Blood from Therapeutically Immunosuppressed or Retrovirus-Infected Cats. Vet. Sci. 2017, 4, 16. [Google Scholar] [CrossRef] [Green Version]
- McLuckie, A.; Barrs, V.; Lindsay, S.; Aghazadeh, M.; Sangster, C.; Beatty, J. Molecular Diagnosis of Felis catus Gammaherpesvirus 1 (FcaGHV1) Infection in Cats of Known Retrovirus Status with and without Lymphoma. Viruses 2018, 10, 128. [Google Scholar] [CrossRef] [Green Version]
- Marenzoni, M.L.; Stefanetti, V.; Danzetta, M.L.; Timoney, P.J. Gammaherpesvirus infections in equids: A review. Vet. Med. 2015, 6, 91–101. [Google Scholar] [CrossRef] [Green Version]
- Niederman, J.C.; Miller, G.; Pearson, H.A.; Pagano, J.S.; Dowaliby, J.M. Infectious mononucleosis. Epstein-Barr-virus shedding in saliva and the oropharynx. N. Engl. J. Med. 1976, 294, 1355–1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casper, C.; Krantz, E.; Selke, S.; Kuntz, S.R.; Wang, J.; Huang, M.L.; Pauk, J.S.; Corey, L.; Wald, A. Frequent and asymptomatic oropharyngeal shedding of human herpesvirus 8 among immunocompetent men. J. Infect. Dis. 2007, 195, 30–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLuckie, A.J.; Barrs, V.R.; Smith, A.L.; Beatty, J.A. Detection of Felis catus gammaherpesvirus 1 (FcaGHV1) in peripheral blood B- and T-lymphocytes in asymptomatic, naturally-infected domestic cats. Virology 2016, 497, 211–216. [Google Scholar] [CrossRef]
- Tizard, I. Veterinary Immunology: An Introduction; WB Saunders, Co.: Philadelphia, PA, USA, 1992. [Google Scholar]
- Chan, K.H.; Tam, J.S.; Peiris, J.S.; Seto, W.H.; Ng, M.H. Epstein-Barr virus (EBV) infection in infancy. J. Clin. Virol. 2001, 21, 57–62. [Google Scholar] [CrossRef]
- Fourcade, G.; Germi, R.; Guerber, F.; Lupo, J.; Baccard, M.; Seigneurin, A.; Semenova, T.; Morand, P.; Epaulard, O. Evolution of EBV seroprevalence and primary infection age in a French hospital and a city laboratory network, 2000–2016. PLoS ONE 2017, 12, e0175574. [Google Scholar] [CrossRef]
- Balfour, H.H., Jr.; Sifakis, F.; Sliman, J.A.; Knight, J.A.; Schmeling, D.O.; Thomas, W. Age-specific prevalence of Epstein-Barr virus infection among individuals aged 6–19 years in the United States and factors affecting its acquisition. J. Infect. Dis. 2013, 208, 1286–1293. [Google Scholar] [CrossRef] [Green Version]
- Kuri, A.; Jacobs, B.M.; Vickaryous, N.; Pakpoor, J.; Middeldorp, J.; Giovannoni, G.; Dobson, R. Epidemiology of Epstein-Barr virus infection and infectious mononucleosis in the United Kingdom. BMC Public Health 2020, 20, 912. [Google Scholar] [CrossRef]
- Lankester, F.; Lugelo, A.; Mnyambwa, N.; Ndabigaye, A.; Keyyu, J.; Kazwala, R.; Grant, D.M.; Relf, V.; Haig, D.M.; Cleaveland, S.; et al. Alcelaphine Herpesvirus-1 (Malignant Catarrhal Fever Virus) in Wildebeest Placenta: Genetic Variation of ORF50 and A9.5 Alleles. PLoS ONE 2015, 10, e0124121. [Google Scholar] [CrossRef] [Green Version]
- Vereide, D.; Sugden, B. Proof for EBV’s sustaining role in Burkitt’s lymphomas. Semin. Cancer Biol. 2009, 19, 389–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon-Lowe, C.; Rickinson, A. The Global Landscape of EBV-Associated Tumors. Front. Oncol. 2019, 9, 713. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Age Group | Oropharyngeal Swab PCR Result | Comparison of Shedding between Age Groups p-Value, Odds Ratio (OR), 95% Confidence Interval (CI) | |||
|---|---|---|---|---|---|
| PCR Positive/Number Tested | Frequency of Shedding (%) | >3 Months and ≤1 Year | >1 Year and ≤6 Years | >6 Years | |
| ≤3 months | 5/94 | 5.3 | p = 0.001 * OR 7.91 95% CI (2.320, 26.979) | p < 0.001 * OR 17.8 95% CI (5.065, 62.557) | p = 0.078 |
| >3 months and ≤1 year | 8/26 | 30.8 | - | p = 0.231 | p = 0.716 |
| >1 year and ≤6 years | 10/20 | 50 | p = 0.231 | - | p = 0.089 |
| >6 years | 3/15 | 33.3 | p = 0.716 | p = 0.089 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rose, E.C.; Tse, T.Y.; Oates, A.W.; Jackson, K.; Pfeiffer, S.; Donahoe, S.L.; Setyo, L.; Barrs, V.R.; Beatty, J.A.; Pesavento, P.A. Oropharyngeal Shedding of Gammaherpesvirus DNA by Cats, and Natural Infection of Salivary Epithelium. Viruses 2022, 14, 566. https://0-doi-org.brum.beds.ac.uk/10.3390/v14030566
Rose EC, Tse TY, Oates AW, Jackson K, Pfeiffer S, Donahoe SL, Setyo L, Barrs VR, Beatty JA, Pesavento PA. Oropharyngeal Shedding of Gammaherpesvirus DNA by Cats, and Natural Infection of Salivary Epithelium. Viruses. 2022; 14(3):566. https://0-doi-org.brum.beds.ac.uk/10.3390/v14030566
Chicago/Turabian StyleRose, Elizabeth C., Tiffany Y. Tse, Andrew W. Oates, Ken Jackson, Susanne Pfeiffer, Shannon L. Donahoe, Laura Setyo, Vanessa R. Barrs, Julia A. Beatty, and Patricia A. Pesavento. 2022. "Oropharyngeal Shedding of Gammaherpesvirus DNA by Cats, and Natural Infection of Salivary Epithelium" Viruses 14, no. 3: 566. https://0-doi-org.brum.beds.ac.uk/10.3390/v14030566