
Outbreaks of Avipoxvirus Clade E in Vaccinated Broiler Breeders with Exacerbated Beak Injuries and Sex Differences in Severity

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of Outbreaks and the Clinical Samples
2.2. Statistical Analysis of the Zootechnical Performance
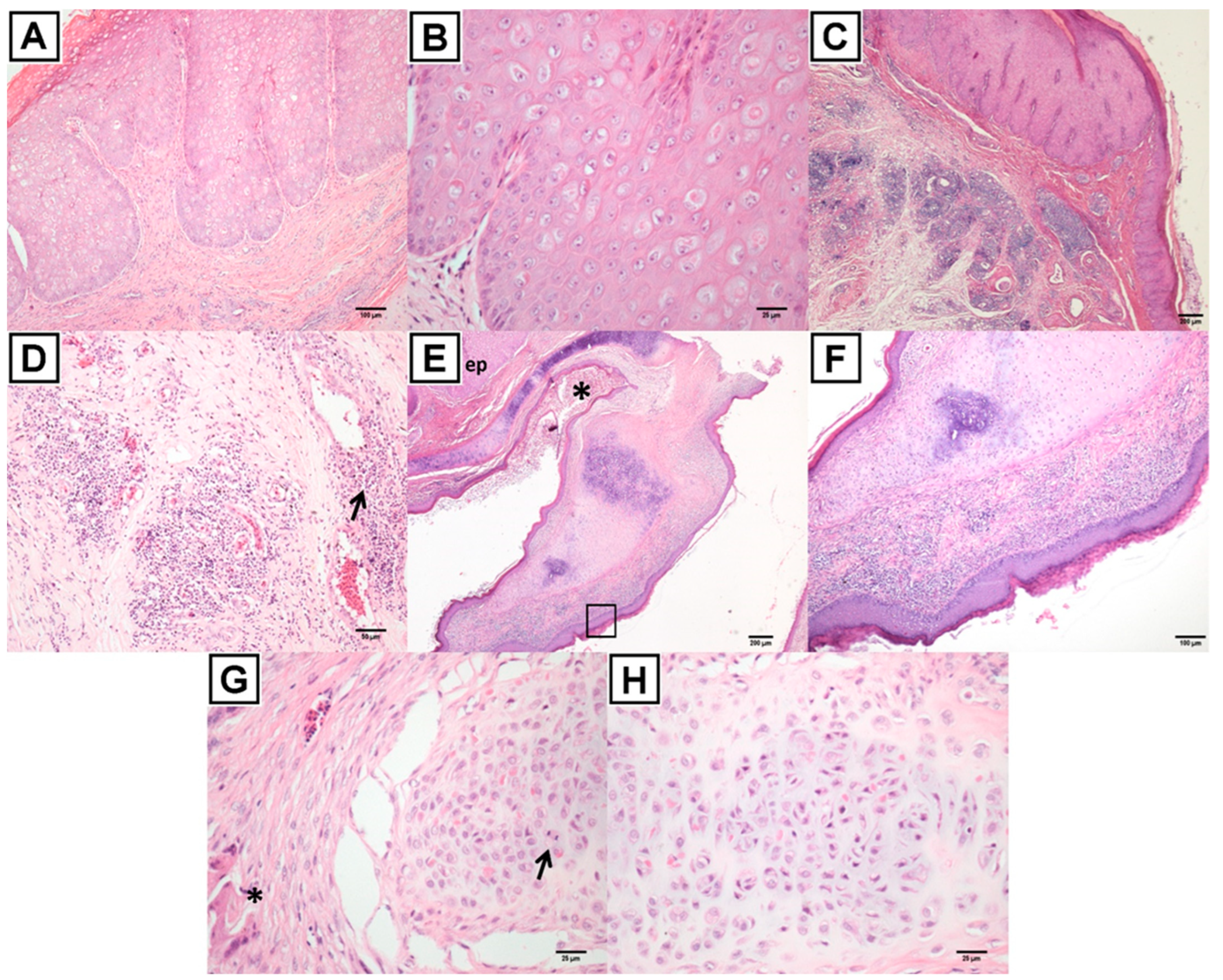
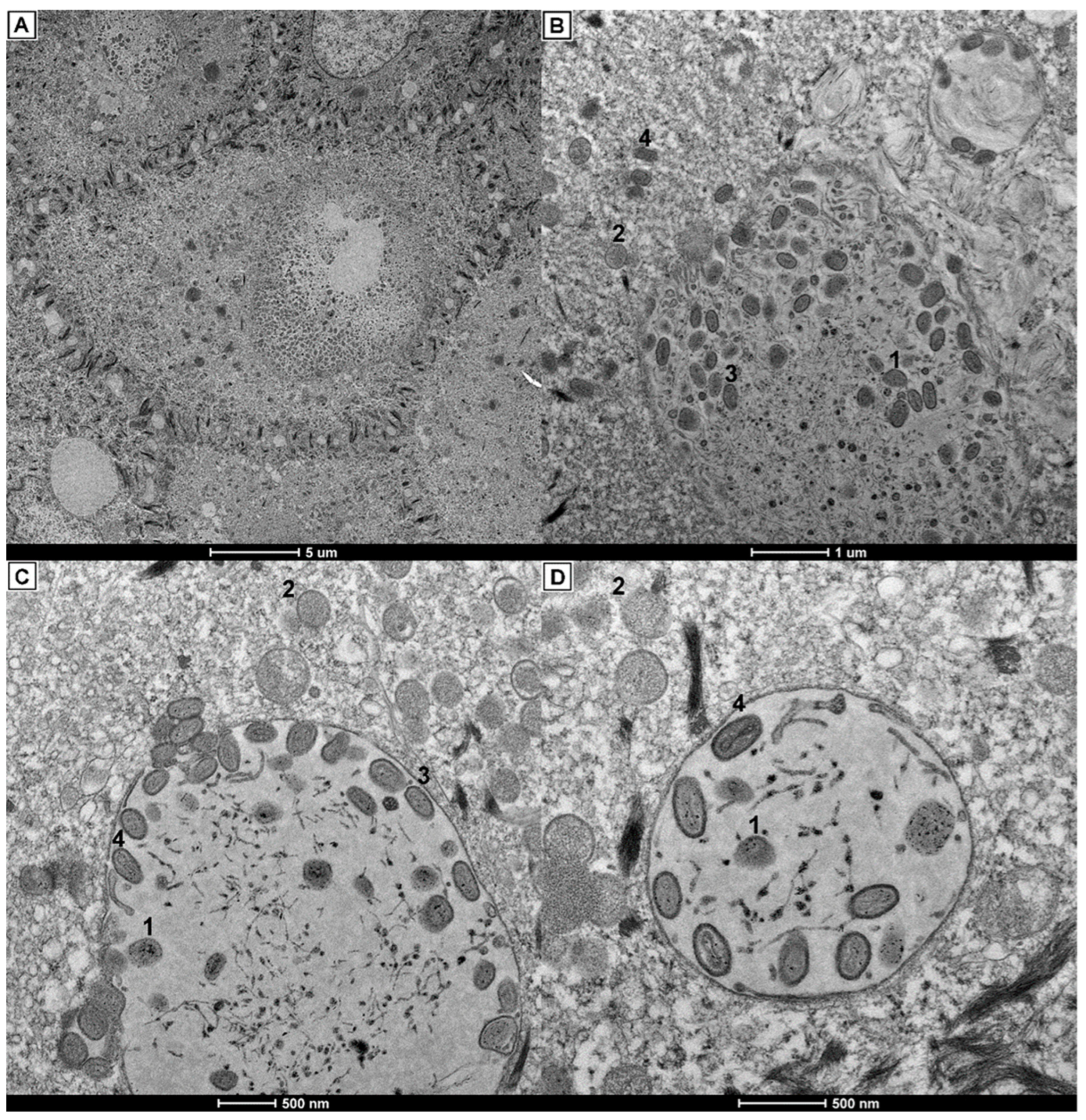
2.3. Histopathology and Electron Microscopy
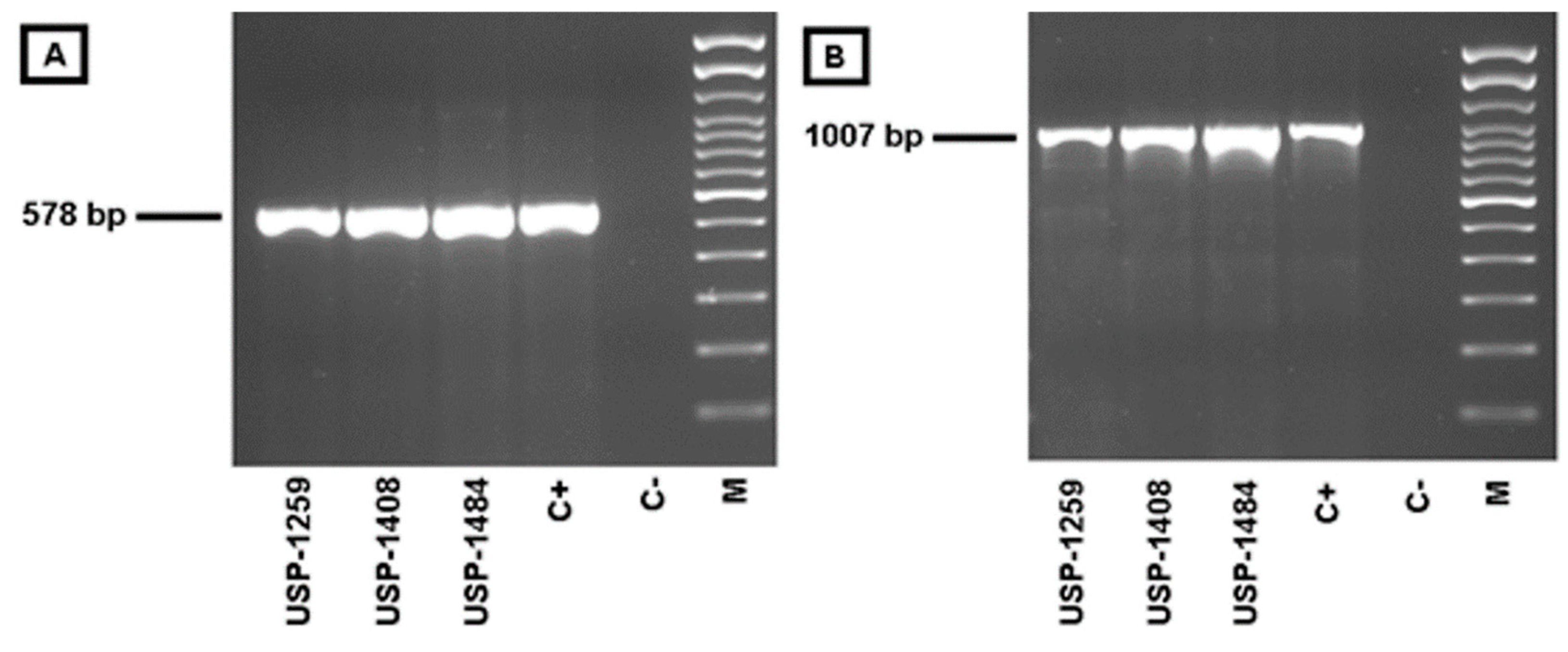
2.4. Nucleic Acid Extraction and PCR Detection of Avipoxvirus
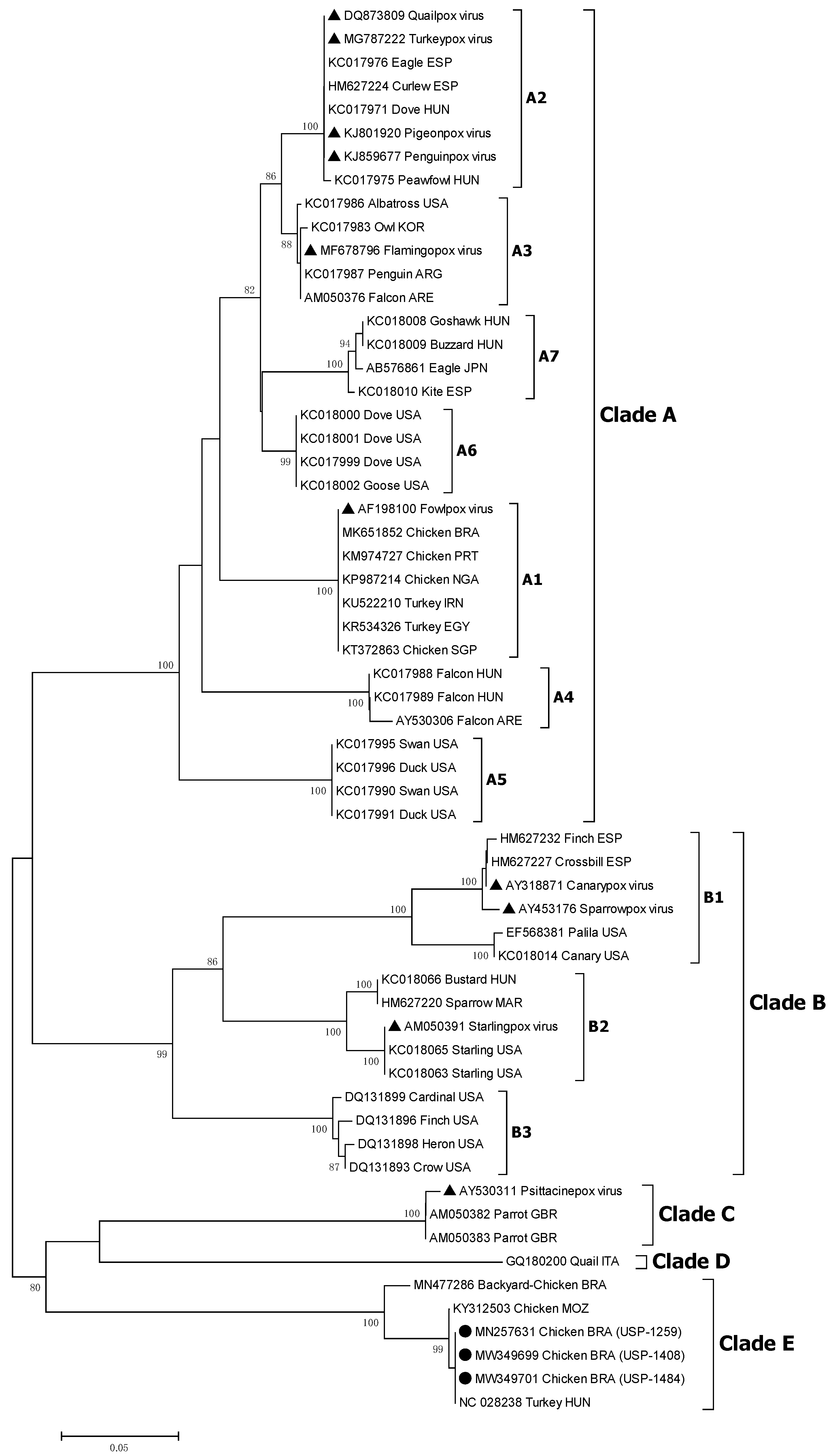
2.5. Sequencing and Phylogenetic Analysis
2.6. Development of qPCR Assays for the Specific Diagnosis and Quantification of Clade E Avipoxvirus
3. Results
3.1. Outbreaks and Clinical Samples
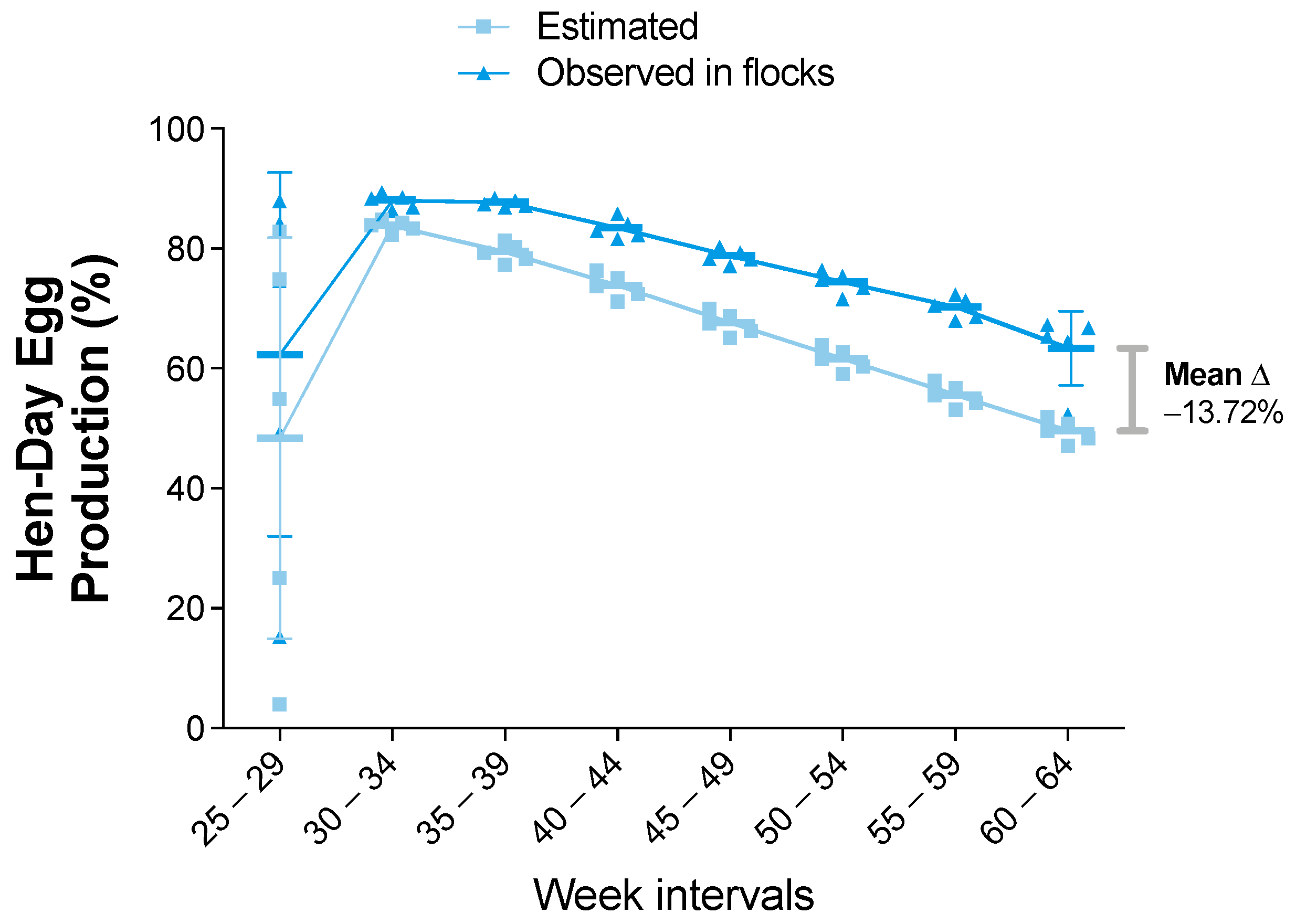
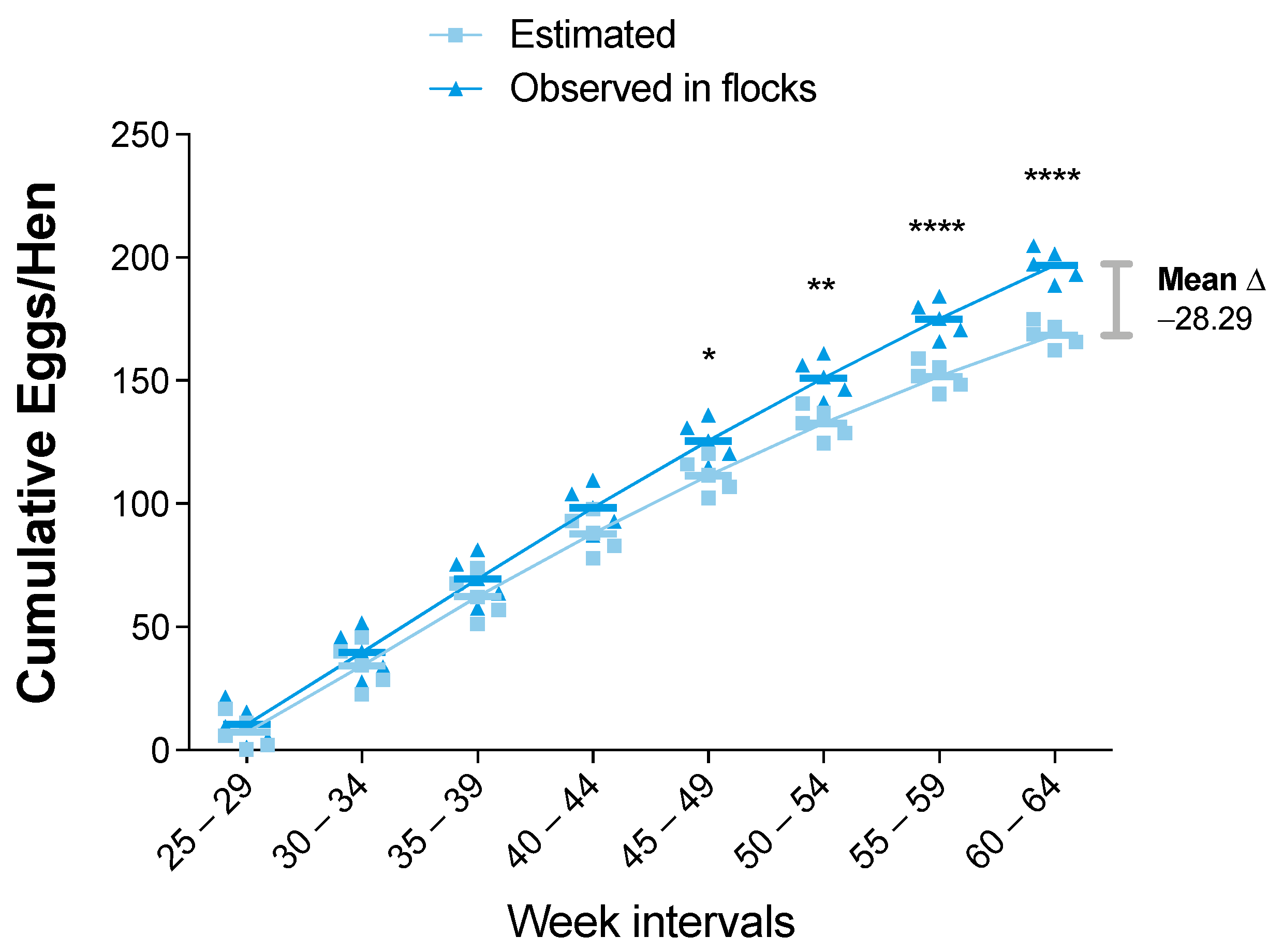
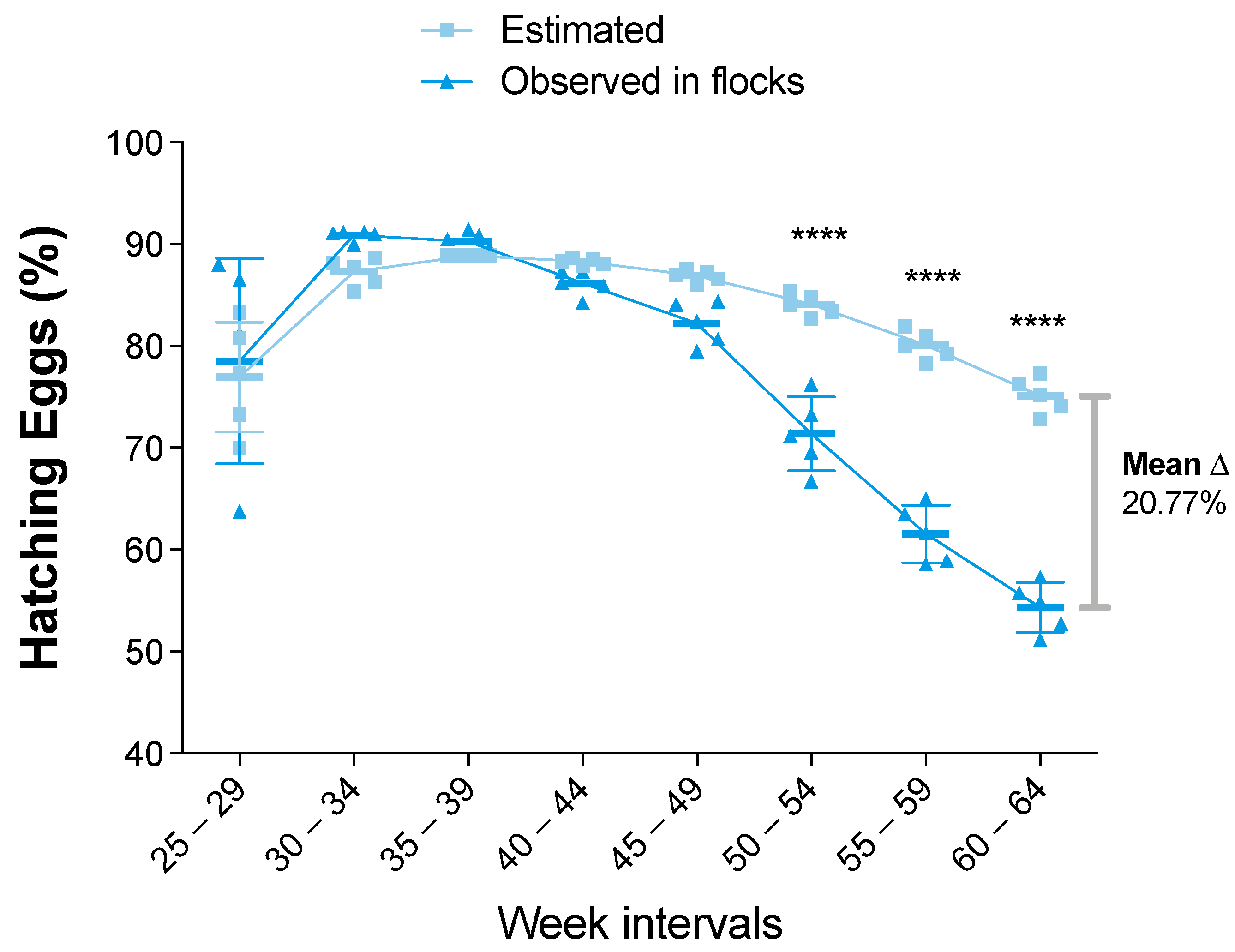
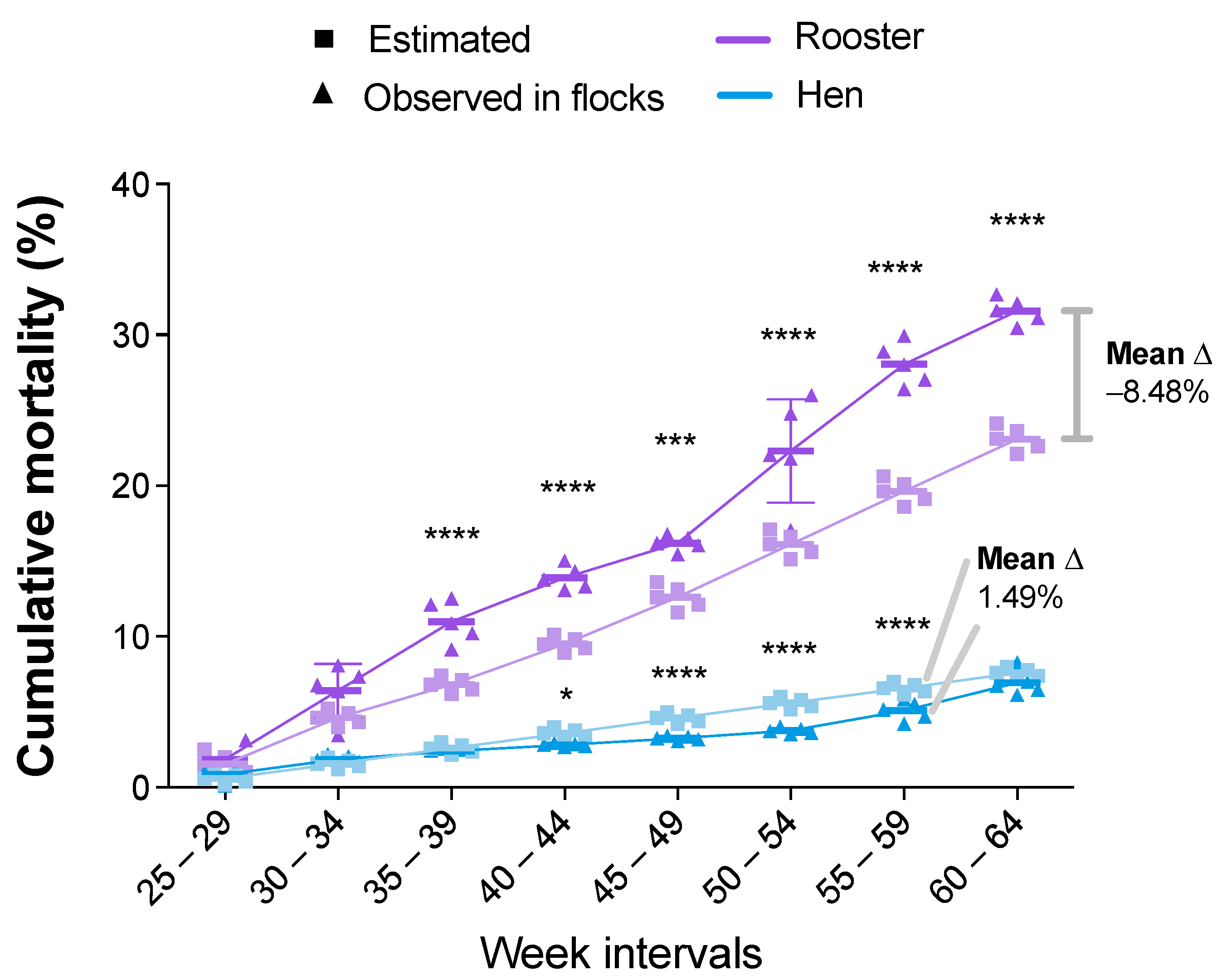
3.2. Statistical Analysis of the Zootechnical Performance
3.3. Histopathology and Electron Microscopy Findings
3.4. Molecular Detection and Characterization of Avipoxvirus
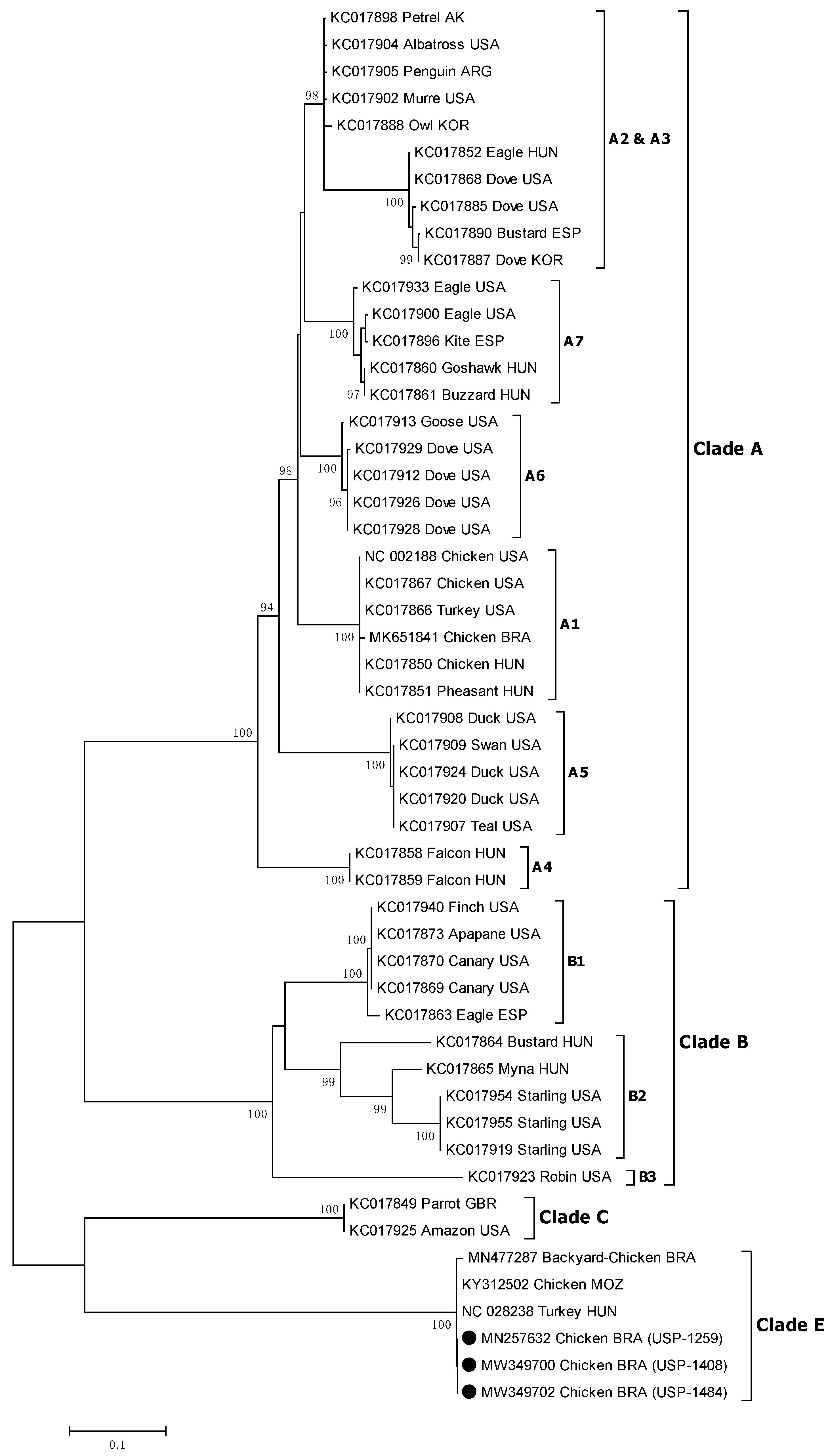
3.5. Sequencing and Phylogenetic Analysis
3.6. Development of qPCR Assays for the Detection and Quantification of Clade E Avipoxvirus
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tripathy, D.N.; Reed, W.M.P. Pox. In Diseases of Poultry; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2020; pp. 364–381. ISBN 978-1-119-37119-9. [Google Scholar]
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Adriaenssens, E.M.; Dempsey, D.M.; Dutilh, B.E.; Harrach, B.; Harrison, R.L.; Hendrickson, R.C.; et al. Changes to Virus Taxonomy and the Statutes Ratified by the International Committee on Taxonomy of Viruses. Arch. Virol. 2020, 165, 2737–2748. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.-J.; Schnitzlein, W.M.; McAloose, D.; Pessier, A.P.; Tripathy, D.N. Characterization of an Avianpox Virus Isolated from an Andean Condor (Vultur gryphus). Vet. Microbiol. 2003, 96, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Sarker, S.; Das, S.; Lavers, J.L.; Hutton, I.; Helbig, K.; Imbery, J.; Upton, C.; Raidal, S.R. Genomic Characterization of Two Novel Pathogenic Avipoxviruses Isolated from Pacific Shearwaters (Ardenna spp.). BMC Genom. 2017, 18, 298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, S.; Batinovic, S.; Talukder, S.; Das, S.; Park, F.; Petrovski, S.; Forwood, J.K.; Helbig, K.J.; Raidal, S.R. Molecular Characterisation of a Novel Pathogenic Avipoxvirus from the Australian Magpie (Gymnorhina tibicen). Virology 2020, 540, 1–16. [Google Scholar] [CrossRef]
- Bolte, A.L.; Meurer, J.; Kaleta, E.F. Avian Host Spectrum of Avipoxviruses. Avian Pathol. 1999, 28, 415–432. [Google Scholar] [CrossRef] [Green Version]
- Giddens, W.E.; Swango, L.J.; Henderson, J.D.; Lewis, R.A.; Farner, D.S.; Carlos, A.; Dolowy, W.C. Canary Pox in Sparrows and Canaries (Fringillidae and in Weavers (Ploceidae). Pathology and Host Specificity of the Virus. Vet. Pathol. 1971, 8, 260–280. [Google Scholar] [CrossRef] [Green Version]
- Giotis, E.S.; Skinner, M.A. Spotlight on Avian Pathology: Fowlpox Virus. Avian Pathol. 2019, 48, 87–90. [Google Scholar] [CrossRef]
- Chacón, R.D.; Astolfi-Ferreira, C.S.; De la Torre, D.I.; de Sá, L.R.M.; Piantino Ferreira, A.J. An Atypical Clinicopathological Manifestation of Fowlpox Virus Associated with Reticuloendotheliosis Virus in Commercial Laying Hen Flocks in Brazil. Transbound. Emerg. Dis. 2020, 67, 2923–2935. [Google Scholar] [CrossRef]
- Joshi, L.R.; Bauermann, F.V.; Hain, K.S.; Kutish, G.F.; Armién, A.G.; Lehman, C.P.; Neiger, R.; Afonso, C.L.; Tripathy, D.N.; Diel, D.G. Detection of Fowlpox Virus Carrying Distinct Genome Segments of Reticuloendotheliosis Virus. Virus Res. 2019, 260, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Schnitzlein, W.M.; Tripathy, D.N. Reticuloendotheliosis Virus Sequences within the Genomes of Field Strains of Fowlpox Virus Display Variability. J. Virol. 2003, 77, 5855–5862. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, V.; Tripathy, D.N. The DNA Repair Enzyme, CPD-Photolyase Restores the Infectivity of UV-Damaged Fowlpox Virus Isolated from Infected Scabs of Chickens. Vet. Microbiol. 2005, 108, 215–223. [Google Scholar] [CrossRef]
- Gyuranecz, M.; Foster, J.T.; Dán, Á.; Ip, H.S.; Egstad, K.F.; Parker, P.G.; Higashiguchi, J.M.; Skinner, M.A.; Höfle, U.; Kreizinger, Z.; et al. Worldwide Phylogenetic Relationship of Avian Poxviruses. J. Virol. 2013, 87, 4938–4951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, B.; Lachish, S.; Colvile, K.M.; Durrant, C.; Peck, K.M.; Toms, M.P.; Sheldon, B.C.; Cunningham, A.A. Emergence of a Novel Avian Pox Disease in British Tit Species. PLoS ONE 2012, 7, e40176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacDonald, A.M.; Gibson, D.J.; Barta, J.R.; Poulson, R.; Brown, J.D.; Allison, A.B.; Nemeth, N.M. Bayesian Phylogenetic Analysis of Avipoxviruses from North American Wild Birds Demonstrates New Insights into Host Specificity and Interspecies Transmission. Avian Dis. 2019, 63, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Manarolla, G.; Pisoni, G.; Sironi, G.; Rampin, T. Molecular Biological Characterization of Avian Poxvirus Strains Isolated from Different Avian Species. Vet. Microbiol. 2010, 140, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Bányai, K.; Palya, V.; Dénes, B.; Glávits, R.; Ivanics, É.; Horváth, B.; Farkas, S.L.; Marton, S.; Bálint, Á.; Gyuranecz, M.; et al. Unique Genomic Organization of a Novel Avipoxvirus Detected in Turkey (Meleagris gallopavo). Infect. Genet. Evol. 2015, 35, 221–229. [Google Scholar] [CrossRef]
- Mapaco, L.P.; Lacerda, Z.; Monjane, I.V.A.; Gelaye, E.; Sussuro, A.H.; Viljoen, G.J.; Dundon, W.G.; Achá, S.J. Identification of Clade E Avipoxvirus, Mozambique, 2016. Emerg. Infect. Dis. 2017, 23, 1602–1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, L.C.; Monteiro, F.L.; Chagas, D.B.; D’Ávila Vargas, G.; de Lima, M.; Fischer, G.; de Oliveira Hübner, S. Identification of Clade E Avipoxvirus in Brazil. Avian Dis. 2020, 64, 223–227. [Google Scholar] [CrossRef]
- Watanabe, I.; Yamada, E. The Fine Structure of Lamellated Nerve Endings Found in the Rat Gingiva. Arch. Histol. Jpn. 1983, 46, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Ciena, A.P.; de Almeida, S.R.Y.; Alves, P.H.d.M.; Bolina-Matos, R.d.S.; Dias, F.J.; Issa, J.P.M.; Iyomasa, M.M.; Watanabe, I. Histochemical and Ultrastructural Changes of Sternomastoid Muscle in Aged Wistar Rats. Micron 2011, 42, 871–876. [Google Scholar] [CrossRef]
- Spurr, A.R. A Low-Viscosity Epoxy Resin Embedding Medium for Electron Microscopy. J. Ultrastruct. Res. 1969, 26, 31–43. [Google Scholar] [CrossRef]
- Ciena, A.P.; de Sousa Bolina, C.; de Almeida, S.R.Y.; Rici, R.E.G.; de Oliveira, M.F.; da Silva, M.C.P.; Miglino, M.A.; Watanabe, I. Structural and Ultrastructural Features of the Agouti Tongue (Dasyprocta aguti linnaeus, 1766). J. Anat. 2013, 223, 152–158. [Google Scholar] [CrossRef] [Green Version]
- Huw Lee, L.; Hwa Lee, K. Application of the Polymerase Chain Reaction for the Diagnosis of Fowl Poxvirus Infection. J. Virol. Methods 1997, 63, 113–119. [Google Scholar] [CrossRef]
- Cao, W.; Mays, J.; Dunn, J.; Fulton, R.; Silva, R.; Fadly, A. Use of Polymerase Chain Reaction in Detection of Marek’s Disease and Reticuloendotheliosis Viruses in Formalin-Fixed, Paraffin-Embedded Tumorous Tissues. Avian Dis. 2013, 57, 785–789. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Afonso, C.L.; Tulman, E.R.; Lu, Z.; Zsak, L.; Kutish, G.F.; Rock, D.L. The Genome of Fowlpox Virus. J. Virol. 2000, 74, 3815–3831. [Google Scholar] [CrossRef] [Green Version]
- Laidlaw, S.M.; Skinner, M.A. Comparison of the Genome Sequence of FP9, an Attenuated, Tissue Culture-Adapted European Strain of Fowlpox Virus, with Those of Virulent American and European Viruses. J. Gen. Virol. 2004, 85, 305–322. [Google Scholar] [CrossRef]
- Offerman, K.; Carulei, O.; van der Walt, A.P.; Douglass, N.; Williamson, A.-L. The Complete Genome Sequences of Poxviruses Isolated from a Penguin and a Pigeon in South Africa and Comparison to Other Sequenced Avipoxviruses. BMC Genom. 2014, 15, 463. [Google Scholar] [CrossRef] [Green Version]
- Carulei, O.; Douglass, N.; Williamson, A.-L. Comparative Analysis of Avian Poxvirus Genomes, Including a Novel Poxvirus from Lesser Flamingos (Phoenicopterus minor), Highlights the Lack of Conservation of the Central Region. BMC Genom. 2017, 18, 947. [Google Scholar] [CrossRef]
- Croville, G.; Le Loc’h, G.; Zanchetta, C.; Manno, M.; Camus-Bouclainville, C.; Klopp, C.; Delverdier, M.; Lucas, M.-N.; Donnadieu, C.; Delpont, M.; et al. Rapid Whole-Genome Based Typing and Surveillance of Avipoxviruses Using Nanopore Sequencing. J. Virol. Methods 2018, 261, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Kost, T.A.; Theodorakis, N.; Hughes, S.H. The Nucleotide Sequence of the Chick Cytoplasmic Beta-Actin Gene. Nucleic Acids Res. 1983, 11, 8287–8301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chacón, R.D.; Astolfi-Ferreira, C.S.; Chacón, J.L.; Nuñez, L.F.N.; De la Torre, D.I.; Piantino Ferreira, A.J. A Seminested RT-PCR for Molecular Genotyping of the Brazilian BR-I Infectious Bronchitis Virus Strain (GI-11). Mol. Cell. Probes 2019, 47, 101426. [Google Scholar] [CrossRef] [PubMed]
- Karpińska, T.A.; Kozaczyński, W.; Niemczuk, K.; Jasik, A.; Kycko, A.; Reichert, M. Mixed Infection by Fowlpox Virus and Chlamydophila Psittaci in a Commercial Laying Hen Flock. Acta Vet. Hung. 2014, 62, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Zhao, K.; He, W.; Xie, S.; Song, D.; Lu, H.; Pan, W.; Zhou, P.; Liu, W.; Lu, R.; Zhou, J.; et al. Highly Pathogenic Fowlpox Virus in Cutaneously Infected Chickens, China. Emerg. Infect. Dis. 2014, 20, 1208–1210. [Google Scholar] [CrossRef]
- Yeo, G.; Wang, Y.; Chong, S.M.; Humaidi, M.; Lim, X.F.; Mailepessov, D.; Chan, S.; How, C.B.; Lin, Y.N.; Huangfu, T.; et al. Characterization of Fowlpox Virus in Chickens and Bird-Biting Mosquitoes: A Molecular Approach to Investigating Avipoxvirus Transmission. J. Gen. Virol. 2019, 100, 838–850. [Google Scholar] [CrossRef]
- Fukui, D.; Nakamura, M.; Yamaguchi, T.; Takenaka, M.; Murakami, M.; Yanai, T.; Fukushi, H.; Yanagida, K.; Bando, G.; Matsuno, K.; et al. An Epizootic of Emerging Novel Avian Pox in Carrion Crows (Corvus corone) and Large-Billed Crows (Corvus macrorhynchos) in Japan. J. Wildl. Dis. 2016, 52, 230–241. [Google Scholar] [CrossRef]
- González-Hein, G.; González, C.; Hidalgo, H. Case Report: An Avian Pox Outbreak in Captive Psittacine Birds in Chile. J. Exot. Pet Med. 2008, 17, 210–215. [Google Scholar] [CrossRef]
- Murer, L.; Westenhofen, M.; Kommers, G.D.; Furian, T.Q.; Borges, K.A.; Kunert-Filho, H.C.; Streck, A.F.; Lovato, M. Identification and Phylogenetic Analysis of Clade C Avipoxvirus in a Fowlpox Outbreak in Exotic Psittacines in Southern Brazil. J. Vet. Diagn. Investig. 2018, 30, 946–950. [Google Scholar] [CrossRef] [Green Version]
- Robinson, F.E.; Hardin, R.T.; Robblee, A.R. Reproductive Senescence in Domestic Fowl: Effects on Egg Production, Sequence Length and Inter-Sequence Pause Length. Br. Poult. Sci. 1990, 31, 871–879. [Google Scholar] [CrossRef]
- Zakaria, A.H.; Omar, O.H. Egg Laying Pattern, Egg Weight, Body Weight at Hatch, and Sex Ratio Bias Relative to Oviposition Time of Young-and Mid-Age Broiler Breeders. Anim. Reprod. Sci. 2013, 141, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.S.H.; Abdul-Careem, M.F. Avian Viruses That Impact Table Egg Production. Animals 2020, 10, 1747. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.L.; Cotter, P.F.; Wing, T.L.; Briles, W.E. Major Histocompatibility (B) Complex and Sex Effects on the Phytohaemagglutinin Wattle Response. Anim. Genet. 1987, 18, 343–350. [Google Scholar] [CrossRef]
- Barbour, E.K.; Hamadeh, S.K.; Hilan, C.; Abbas, S.S. Comparison of Immunity and Resistance to Diseases in Male and Female Poultry Breeders in Lebanon. Trop. Anim. Health Prod. 1995, 27, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Barreda, C.B. Relationship Between Values of Fowlpox ELISA and the Presence of “Takes” After Vaccination. Avian Dis. 2016, 60, 67–69. [Google Scholar] [CrossRef] [PubMed]
- Weli, S.C.; Tryland, M. Avipoxviruses: Infection Biology and Their Use as Vaccine Vectors. Virol. J. 2011, 8, 49. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Yousaf, A.; Khan, M.Z.; Siddique, M.; Gul, S.T.; Mahmood, F. Cutaneous Form of Pox Infection among Captive Peafowl (Pavo cristatus) Chicks. Avian Pathol. 2009, 38, 65–70. [Google Scholar] [CrossRef]
- Tripathy, D.N.; Schnitzlein, W.M.; Morris, P.J.; Janssen, D.L.; Zuba, J.K.; Massey, G.; Atkinson, C.T. Characterization of Poxviruses from Forest Birds in Hawaii. J. Wildl. Dis. 2000, 36, 225–230. [Google Scholar] [CrossRef] [Green Version]
- Groupe, V.; Oskay, J.; Rake, G. Electron Micrographs of the Elementary Bodies of Fowl Pox and Canary Pox. Proc. Soc. Exp. Biol. Med. 1946, 63, 477. [Google Scholar] [CrossRef]
- Tudor, D.C.; Rue, J.W.; Woodward, H.L. Electron Scanning Microscope Studies in Pigeon Pox Virus. Poult. Sci. 1975, 54, 814–824. [Google Scholar] [CrossRef]
- Chacón, R.D.; Astolfi-Ferreira, C.S.; Guimarães, M.B.; Torres, L.N.; De la Torre, D.I.; de Sá, L.R.M.; Piantino Ferreira, A.J. Detection and Molecular Characterization of a Natural Coinfection of Marek’s Disease Virus and Reticuloendotheliosis Virus in Brazilian Backyard Chicken Flock. Vet. Sci. 2019, 6, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoerr, F.J. Clinical Aspects of Immunosuppression in Poultry. Avian Dis. 2010, 54, 2–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gimeno, I.M.; Schat, K.A. Virus-Induced Immunosuppression in Chickens. Avian Dis. 2018, 62, 272–285. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, F.; Yamamoto, Y.; Sato, Y.; Fukunari, K.; Murata, K.-I.; Yaegashi, G.; Goto, M.; Murakami, R. Concurrent Fowlpox and Candidiasis Diseases in Backyard Chickens with Unusual Pox Lesions in the Bursa of Fabricius. Avian Dis. 2016, 60, 705–708. [Google Scholar] [CrossRef] [PubMed]
- Shivaprasad, H.L.; Kim, T.; Tripathy, D.; Woolcock, P.R.; Uzal, F. Unusual Pathology of Canary Poxvirus Infection Associated with High Mortality in Young and Adult Breeder Canaries (Serinus canaria). Avian Pathol. 2009, 38, 311–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landolt, M.; Kocan, R.M. Transmission of Avian Pox from Starlings to Rothchild’s Mynahs. J. Wildl. Dis. 1976, 12, 353–356. [Google Scholar] [CrossRef]
- Donnelly, T.M.; Crane, L.A. An Epornitic of Avian Pox in a Research Aviary. Avian Dis. 1984, 28, 517–525. [Google Scholar] [CrossRef]
- Fatunmbi, O.O.; Reed, W.M. Evaluation of a Commercial Modified Live Virus Fowl Pox Vaccine for the Control of “Variant” Fowl Poxvirus Infections. Avian Dis. 1996, 40, 582–587. [Google Scholar] [CrossRef]
- Fallavena, L.C.B.; Canal, C.W.; Salle, C.T.P.; Moraes, H.L.S.; Rocha, S.L.S.; Pereira, R.A.; da Silva, A.B. Presence of Avipoxvirus DNA in Avian Dermal Squamous Cell Carcinoma. Avian Pathol. 2002, 31, 241–246. [Google Scholar] [CrossRef]
- Schnitzlein, W.M.; Ghildyal, N.; Tripathy, D.N. Genomic and Antigenic Characterization of Avipoxviruses. Virus Res. 1988, 10, 65–75. [Google Scholar] [CrossRef]
- Singh, P.; Kim, T.J.; Tripathy, D.N. Re-Emerging Fowlpox: Evaluation of Isolates from Vaccinated Flocks. Avian Pathol. 2000, 29, 449–455. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer | Sequence (5’ > 3’) | Location * | Product Length |
|---|---|---|---|---|
| ORF49 | qEPox49-F | GCTGATTACGGGATCTATGTTG | 52,912–52,933 A | 107 bp |
| qEPox49-R | TAGTTGCCTTTGTATCTGCG | 52,999–53,018 A | ||
| E49-Fex | CTGCCAATGCTATCGATACC | 52,730–52,749 A | 449 bp | |
| E49-Rex | GCAACAAAACGAGAGGTTTC | 53,159–53,178 A | ||
| ORF114 | qEPox114-F | GGACTCAACAAACGTGCTAT | 118,713–118,732 A | 177 bp |
| qEPox114-R | CTGTTCATTAGACGTCGTGC | 118,870–118,889 A | ||
| E114-Fex | GGCTAGATTGATAACAGCTATGG | 118,494–118,516 A | 434 bp | |
| E114-Rex | TGATAGTCGTCTTTATACGAGCAC | 118,904–118,927 A | ||
| ORF159 | qEPox159-F | AGATGGTGATGATTACGGATG | 168,643–168,663 A | 104 bp |
| qEPox159-R | CAGATACGCTAGACCAATCAG | 168,726–168,746 A | ||
| E159-Fex | CGTATATGCCTCTACTTGTAATTG | 168,451–168,474 A | 511 bp | |
| E159-Rex | ACTTTCTCCCTTATCAGCAAC | 168,941–168,961 A | ||
| B actin | qβactin-F | TCAGACTCTGGAGTGCCTTG | 3913–3932 B | 101 bp |
| qβactin-R | GGTCAGATGCAGTGTGATGG | 3994–4013 B | ||
| βactin-Fex | GCTGCTCTTAGCACACCTCTT | 3694–3714 B | 346 bp | |
| βactin-Rex | GCAATGATCTGCAGGAGAGA | 4020–4039 B |
| CLADE | SUBCLADE | Comparative Identity with Strains from This Study | |||
|---|---|---|---|---|---|
| Partial P4b Gene | Partial Pol Gene | ||||
| Nucleotides | Amino Acids | Nucleotides | Amino Acids | ||
| A | A1 | 72.5–73.9 | 75.9–76.6 | 72.3–73.6 | 77.4–78.1 |
| A2 | 73.2–74.9 | 75.6–80.3 | 72.7–74.3 | 75.7–77.0 | |
| A3 | 72.7–74.1 | 75.4–76.8 | 73.8–74.2 | 76.4–77.2 | |
| A4 | 72.2–73.5 | 74.6–75.1 | 74.6–74.7 | 77.7–78.2 | |
| A5 | 73.3–73.5 | 74.5–74.6 | 72.5–72.6 | 76.4–76.8 | |
| A6 | 72.8–73.0 | 75.7–75.8 | 74.3–75.0 | 78.3–79.3 | |
| A7 | 72.0–73.6 | 75.0–77-8 | 72.4–74.5 | 77.1–78.2 | |
| B | B1 | 71.1–71.9 | 73.9–74.3 | 73.8–74.0 | 76.9–77.4 |
| B2 | 74.6–75.3 | 76.3–77.3 | 72.2–74.3 | 74.8–77.2 | |
| B3 | 72.8–73.4 | 76.6–77.2 | 74.0 | 77.2–77.4 | |
| C | 74.5–75.2 | 82.7–83.7 | 73.3–73.4 | 78.7–79.3 | |
| D | 69.0 | 76.9 | – | – | |
| E | 95.8–100.0 | 92.5–100.0 | 99.4–99.9 | 98.6–99.7 | |
| Target | Standard Curve | USP-1259 | USP-1408 | USP-1484 | ||||
|---|---|---|---|---|---|---|---|---|
| E A | Tm | GC | GC/Cell | GC | GC/Cell | GC | GC/Cell | |
| ORF49 | 96.44% | 72.34 °C ± 0.14 | 3.95 × 105 ± 0.24 | 1.07 × 102 | 7.66 × 104 ± 1.49 | 1.94 × 100 | 9.83 × 105 ± 2.11 | 1.81 × 101 |
| ORF114 | 99.14% | 75.05 °C ± 0.15 | 2.51 × 106 ± 0.65 | 6.87 × 102 | 4.38 × 105 ± 1.41 | 1.11 × 101 | 3.30 × 106 ± 0.85 | 6.08 × 101 |
| ORF159 | 96.49% | 73.86 °C ± 0.15 | 4.00 × 105 ± 0.08 | 1.09 × 102 | 1.08 × 105 ± 0.04 | 2.73 × 100 | 6.63 × 105 ± 0.35 | 1.22 × 101 |
| B-actin | 95.03% | 84.10 °C ± 0.16 | 3.65 × 103 ± 0.10 | ─ | 3.95 × 104 ± 2.97 | ─ | 1.64 × 105 ± 0.27 | ─ |
| GC Range | 1.07–6.87 × 102 GC/cell | 0.19–1.11 × 101 GC/cell | 1.22–6.08 × 101 GC/cell | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chacón, R.D.; Astolfi-Ferreira, C.S.; Pereira, P.C.; Assayag, M.S., Jr.; Campos-Salazar, A.B.; De la Torre, D.; Sá, L.R.M.d.; Almeida, S.R.Y.d.; Rici, R.E.G.; Ferreira, A.J.P. Outbreaks of Avipoxvirus Clade E in Vaccinated Broiler Breeders with Exacerbated Beak Injuries and Sex Differences in Severity. Viruses 2022, 14, 773. https://0-doi-org.brum.beds.ac.uk/10.3390/v14040773
Chacón RD, Astolfi-Ferreira CS, Pereira PC, Assayag MS Jr., Campos-Salazar AB, De la Torre D, Sá LRMd, Almeida SRYd, Rici REG, Ferreira AJP. Outbreaks of Avipoxvirus Clade E in Vaccinated Broiler Breeders with Exacerbated Beak Injuries and Sex Differences in Severity. Viruses. 2022; 14(4):773. https://0-doi-org.brum.beds.ac.uk/10.3390/v14040773
Chicago/Turabian StyleChacón, Ruy D., Claudete S. Astolfi-Ferreira, Patrícia C. Pereira, Mario S. Assayag, Jr., Antony B. Campos-Salazar, David De la Torre, Lilian R. M. de Sá, Sonia R. Yokomizo de Almeida, Rose Elí Grassi Rici, and Antonio J. Piantino Ferreira. 2022. "Outbreaks of Avipoxvirus Clade E in Vaccinated Broiler Breeders with Exacerbated Beak Injuries and Sex Differences in Severity" Viruses 14, no. 4: 773. https://0-doi-org.brum.beds.ac.uk/10.3390/v14040773