
Metagenomic Analysis Reveals High Abundance of Torque Teno Mini Virus in the Respiratory Tract of Children with Acute Respiratory Illness


 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples Selection
2.2. Metagenomic Workflow
2.3. Statistical Analysis
3. Results
3.1. Study Population
3.2. Composition of Metagenomic Sequences
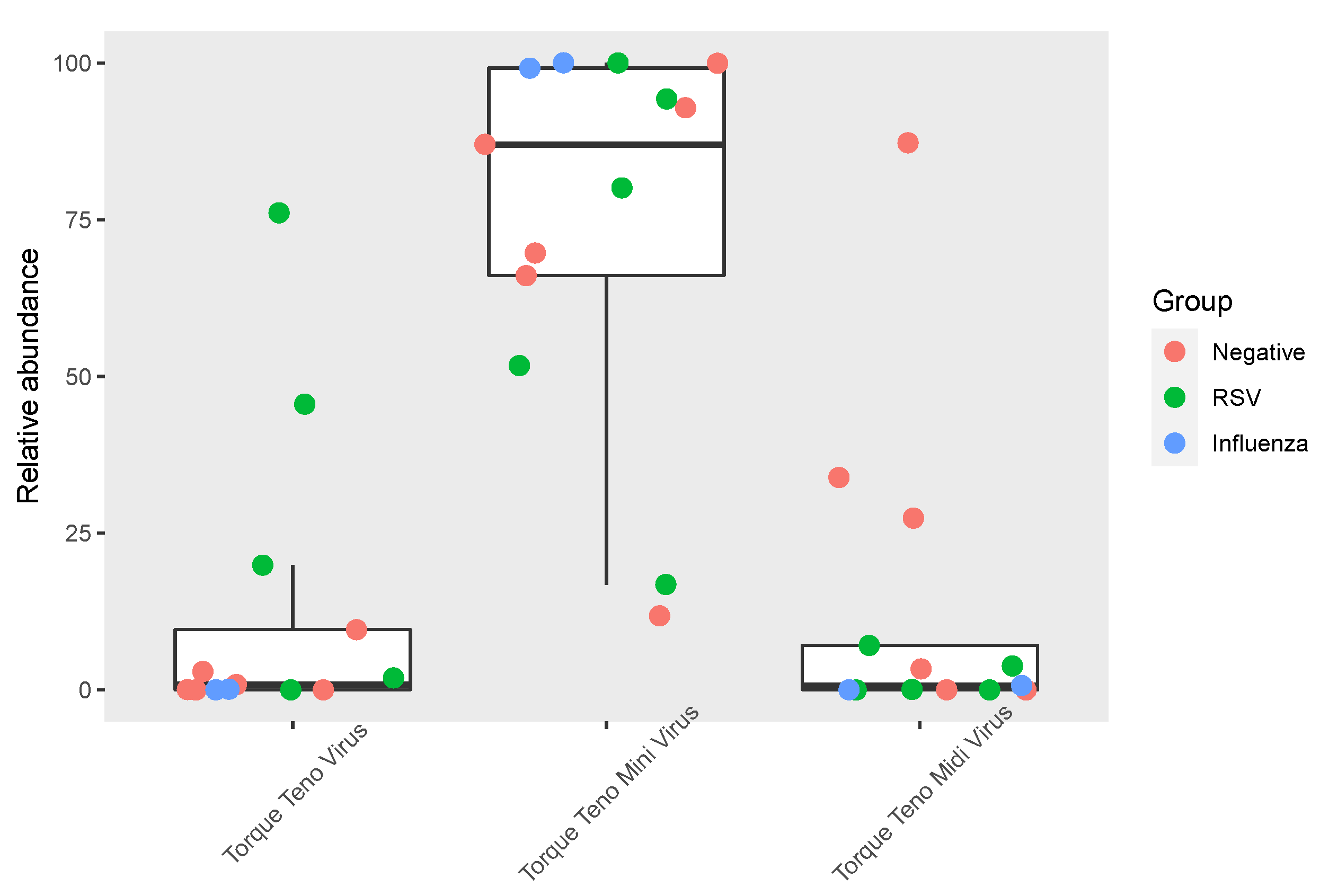
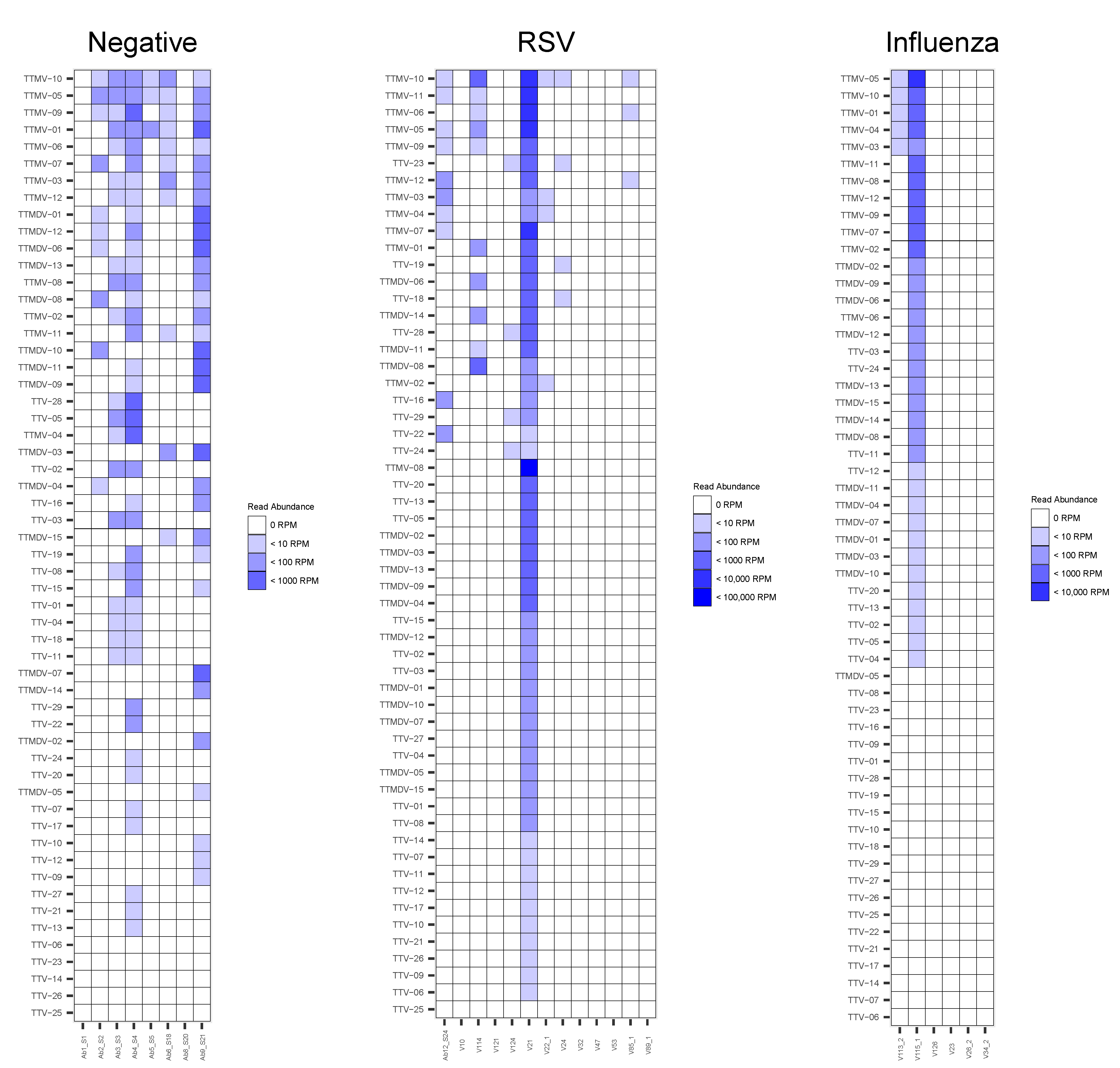
3.3. Anelloviridae Abundance and Composition within the Respiratory Virome
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Biagini, P. Classification of TTV and related viruses (anelloviruses). Curr. Top. Microbiol. Immunol. 2009, 331, 21–33. [Google Scholar] [CrossRef]
- Anelloviridae—ssDNA Viruses (2011)—ssDNA Viruses (2011)—ICTV. Available online: https://talk.ictvonline.org/ictv-reports/ictv_9th_report/ssdna-viruses-2011/w/ssdna_viruses/139/anelloviridae (accessed on 8 March 2022).
- Jaksch, P.; Kundi, M.; Görzer, I.; Muraközy, G.; Lambers, C.; Benazzo, A.; Hoetzenecker, K.; Klepetko, W.; Puchhammer-Stöckl, E. Torque teno virus as a novel biomarker targeting the efficacy of immunosuppression after lung transplantation. J. Infect. Dis. 2018, 218, 1922–1928. [Google Scholar] [CrossRef] [PubMed]
- De Vlaminck, I.; Khush, K.K.; Strehl, C.; Kohli, B.; Luikart, H.; Neff, N.F.; Okamoto, J.; Snyder, T.M.; Cornfield, D.N.; Nicolls, M.R.; et al. Temporal response of the human virome to immunosuppression and antiviral therapy. Cell 2013, 155, 1178–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spandole, S.; Cimponeriu, D.; Berca, L.M.; Mihăescu, G. Human anelloviruses: An update of molecular, epidemiological and clinical aspects. Arch. Virol. 2015, 160, 893–908. [Google Scholar] [CrossRef] [PubMed]
- Bal, A.; Sarkozy, C.; Josset, L.; Cheynet, V.; Oriol, G.; Becker, J.; Vilchez, G.; Sesques, P.; Mallet, F.; Pachot, A.; et al. Metagenomic next-generation sequencing reveals individual composition and dynamics of anelloviruses during autologous stem cell transplant recipient management. Viruses 2018, 10, 633. [Google Scholar] [CrossRef] [Green Version]
- Popgeorgiev, N.; Temmam, S.; Raoult, D.; Desnues, C. Describing the silent human virome with an emphasis on giant viruses. Intervirology 2013, 56, 395–412. [Google Scholar] [CrossRef]
- Arze, C.A.; Springer, S.; Dudas, G.; Patel, S.; Bhattacharyya, A.; Swaminathan, H.; Brugnara, C.; Delagrave, S.; Ong, T.; Kahvejian, A.; et al. Global genome analysis reveals a vast and dynamic anellovirus landscape within the human virome. Cell Host Microbe 2021, 29, 1305–1315.e6. [Google Scholar] [CrossRef]
- Wang, Y.; Zhu, N.; Li, Y.; Lu, R.; Wang, H.; Liu, G.; Zou, X.; Xie, Z.; Tan, W. Metagenomic analysis of viral genetic diversity in respiratory samples from children with severe acute respiratory infection in China. Clin. Microbiol. Infect. 2016, 22, 458.e1–458.e9. [Google Scholar] [CrossRef] [Green Version]
- Galmès, J.; Li, Y.; Rajoharison, A.; Ren, L.; Dollet, S.; Richard, N.; Vernet, G.; Javouhey, E.; Wang, J.; Telles, J.-N.; et al. Potential implication of new torque teno mini viruses in parapneumonic empyema in children. Eur. Respir. J. 2013, 42, 470–479. [Google Scholar] [CrossRef] [Green Version]
- Freer, G.; Maggi, F.; Pifferi, M.; Di Cicco, M.E.; Peroni, D.G.; Pistello, M. The virome and its major component, anellovirus, a convoluted system molding human immune defenses and possibly affecting the development of asthma and respiratory diseases in childhood. Front. Microbiol. 2018, 9, 686. [Google Scholar] [CrossRef] [Green Version]
- Espinoza, J.A.; Moreno-Valencia, Y.; Coronel-Tellez, R.H.; Castillejos-López, M.; Hernández, A.; Dominguez, A.; Miliar-García, A.; Barbachano-Guerrero, A.; Perez-Padilla, R.; Alejandre-Garcia, A.; et al. Virome and bacteriome characterization of children with pneumonia and asthma in Mexico City during winter seasons 2014 and 2015. PLoS ONE 2018, 13, e0192878. [Google Scholar] [CrossRef] [Green Version]
- Dodi, G.; Attanasi, M.; Di Filippo, P.; Di Pillo, S.; Chiarelli, F. Virome in the lungs: The role of anelloviruses in childhood respiratory diseases. Microorganisms 2021, 9, 1357. [Google Scholar] [CrossRef] [PubMed]
- Del Rosal, T.; García-García, M.L.; Casas, I.; Alcolea, S.; Iglesias-Caballero, M.; Pozo, F.; Rodrigo-Muñoz, J.M.; del Pozo, V.; Calvo, C. Torque teno virus in nasopharyngeal aspirate of children with respiratory infections. Res. Square 2022, preprint. [Google Scholar] [CrossRef]
- Prasetyo, A.A.; Desyardi, M.N.; Tanamas, J.; Suradi; Reviono; Harsini; Kageyama, S.; Chikumi, H.; Shimizu, E. Respiratory viruses and torque teno virus in adults with acute respiratory infections. Intervirology 2015, 58, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Leijonhufvud, G.; Bajalan, A.; Soratto, T.A.T.; Gustafsson, B.; Bogdanovic, G.; Bjerkner, A.; Allander, T.; Ljungman, G.; Andersson, B. Better detection of Torque teno virus in children with leukemia by metagenomic sequencing than by quantitative PCR. J. Med Virol. 2022, 94, 634–641. [Google Scholar] [CrossRef] [PubMed]
- Mokili, J.L.; Rohwer, F.; Dutilh, B.E. Metagenomics and future perspectives in virus discovery. Curr. Opin. Virol. 2012, 2, 63–77. [Google Scholar] [CrossRef]
- Taboada, B.; Morán, P.; Serrano-Vázquez, A.; Iša, P.; Rojas-Velázquez, L.; Pérez-Juárez, H.; López, S.; Torres, J.; Ximenez, C.; Arias, C.F. The gut virome of healthy children during the first year of life is diverse and dynamic. PLoS ONE 2021, 16, e0240958. [Google Scholar] [CrossRef]
- Bal, A.; Pichon, M.; Picard, C.; Casalegno, J.S.; Valette, M.; Schuffenecker, I.; Billard, L.; Vallet, S.; Vilchez, G.; Cheynet, V.; et al. Quality control implementation for universal characterization of DNA and RNA viruses in clinical respiratory samples using single metagenomic next-generation sequencing workflow. BMC Infect. Dis. 2018, 18, 537. [Google Scholar] [CrossRef]
- Eibach, D.; Hogan, B.; Sarpong, N.; Winter, D.; Struck, N.S.; Adu-Sarkodie, Y.; Owusu-Dabo, E.; Schmidt-Chanasit, J.; May, J.; Cadar, D. Viral metagenomics revealed novel betatorquevirus species in pediatric inpatients with encephalitis/meningoencephalitis from Ghana. Sci. Rep. 2019, 9, 2360. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, F.; Shan, T.-L.; Deng, X.; Delwart, E.; Feng, X.-P. A novel species of torque teno mini virus (TTMV) in gingival tissue from chronic periodontitis patients. Sci. Rep. 2016, 6, 26739. [Google Scholar] [CrossRef] [Green Version]
- TeKippe, E.M.; Wylie, K.M.; Deych, E.; Sodergren, E.; Weinstock, G.; Storch, G.A. Increased prevalence of anellovirus in pediatric patients with fever. PLoS ONE 2012, 7, e50937. [Google Scholar] [CrossRef]
- Unger, S.A.; Bogaert, D. The respiratory microbiome and respiratory infections. J. Infect. 2017, 74, S84–S88. [Google Scholar] [CrossRef]
- Maggi, F.; Pifferi, M.; Fornai, C.; Andreoli, E.; Tempestini, E.; Vatteroni, M.; Presciuttini, S.; Marchi, S.; Pietrobelli, A.; Boner, A.; et al. TT Virus in the nasal secretions of children with acute respiratory diseases: Relations to viremia and disease severity. J. Virol. 2003, 77, 2418–2425. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Group | Sample ID | Total Number of Reads | Number of Reads for PCR-Positive Virus | % Human | % Viruses | % Bacteria |
|---|---|---|---|---|---|---|
| Negative samples (n = 8) | Ab1_S1 | 13,242,872 | NA | 68.1 | 0.2 | 39.8 |
| Ab2_S2 | 21,124,748 | NA | 53.7 | 1.0 | 36.1 | |
| Ab3_S3 | 18,613,428 | NA | 88.5 | 2.7 | 32.7 | |
| Ab4_S4 | 5,729,468 | NA | 40.9 | 2.1 | 38.3 | |
| Ab5_S5 | 6,506,898 | NA | 74.6 | 0.4 | 38.6 | |
| Ab6_S18 | 9,719,338 | NA | 58.1 | 0.2 | 36.8 | |
| Ab8_S20 | 12,453,484 | NA | 58.9 | 0.2 | 50.2 | |
| Ab9_S21 | 5,124,554 | NA | 42.2 | 1.7 | 42.1 | |
| RSV samples (n = 13) | Ab12_S24 | 10,533,108 | 232,827 | 67.6 | 1.6 | 35.2 |
| V10 | 4,547,410 | 102 | 87.0 | 0.0 | 3.7 | |
| V32 | 14,911,096 | 57,606 | 88.5 | 0.4 | 2.1 | |
| V53 | 10,826,666 | 361,365 | 88.8 | 3.4 | 1.5 | |
| V89_1 | 6,622,864 | 58,284 | 87.5 | 0.9 | 2.2 | |
| V47 | 11,747,124 | 182,776 | 85.9 | 1.6 | 3.1 | |
| V22_1 | 16,819,624 | 77 | 87.6 | 0.0 | 2.8 | |
| V124 | 19,598,588 | 540,934 | 84.3 | 2.8 | 3.0 | |
| V121 | 14,490,590 | 62,279 | 87.9 | 0.4 | 2.1 | |
| V24 | 8,452,260 | 81,253 | 8.6 | 24.0 | 7.0 | |
| V85_1 | 19,138,760 | 892 | 83.8 | 0.0 | 4.2 | |
| V114 | 12,516,284 | 1,673,602 | 52.0 | 13.6 | 9.7 | |
| V21 | 16,928,282 | 406,594 | 64.9 | 6.6 | 6.0 | |
| Influenza samples (n = 6) | V126 | 7,910,142 | 1125 | 87.2 | 0.01 | 3.6 |
| V23 | 18,131,472 | 103 | 90.7 | 0.001 | 0.6 | |
| V26_2 | 13,672,726 | 53,556 | 82.7 | 0.5 | 4.8 | |
| V34_2 | 11,880,612 | 8058 | 90.0 | 0.1 | 1.4 | |
| V113_2 | 10,944,276 | 78,193 | 49.6 | 0.7 | 16.0 | |
| V115_1 | 11,403,046 | 2108 | 3.0 | 59.9 | 6.7 | |
| NTC | NTC | 7,210,744 | NA | 8.5 | 0.04 | 46.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bal, A.; Destras, G.; Sabatier, M.; Pichon, M.; Regue, H.; Oriol, G.; Gillet, Y.; Lina, B.; Brengel-Pesce, K.; Josset, L.; et al. Metagenomic Analysis Reveals High Abundance of Torque Teno Mini Virus in the Respiratory Tract of Children with Acute Respiratory Illness. Viruses 2022, 14, 955. https://0-doi-org.brum.beds.ac.uk/10.3390/v14050955
Bal A, Destras G, Sabatier M, Pichon M, Regue H, Oriol G, Gillet Y, Lina B, Brengel-Pesce K, Josset L, et al. Metagenomic Analysis Reveals High Abundance of Torque Teno Mini Virus in the Respiratory Tract of Children with Acute Respiratory Illness. Viruses. 2022; 14(5):955. https://0-doi-org.brum.beds.ac.uk/10.3390/v14050955
Chicago/Turabian StyleBal, Antonin, Gregory Destras, Marina Sabatier, Maxime Pichon, Hadrien Regue, Guy Oriol, Yves Gillet, Bruno Lina, Karen Brengel-Pesce, Laurence Josset, and et al. 2022. "Metagenomic Analysis Reveals High Abundance of Torque Teno Mini Virus in the Respiratory Tract of Children with Acute Respiratory Illness" Viruses 14, no. 5: 955. https://0-doi-org.brum.beds.ac.uk/10.3390/v14050955