
Nuances of Responses to Two Sources of Grapevine Leafroll Disease on Pinot Noir Grown in the Field for 17 Years

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
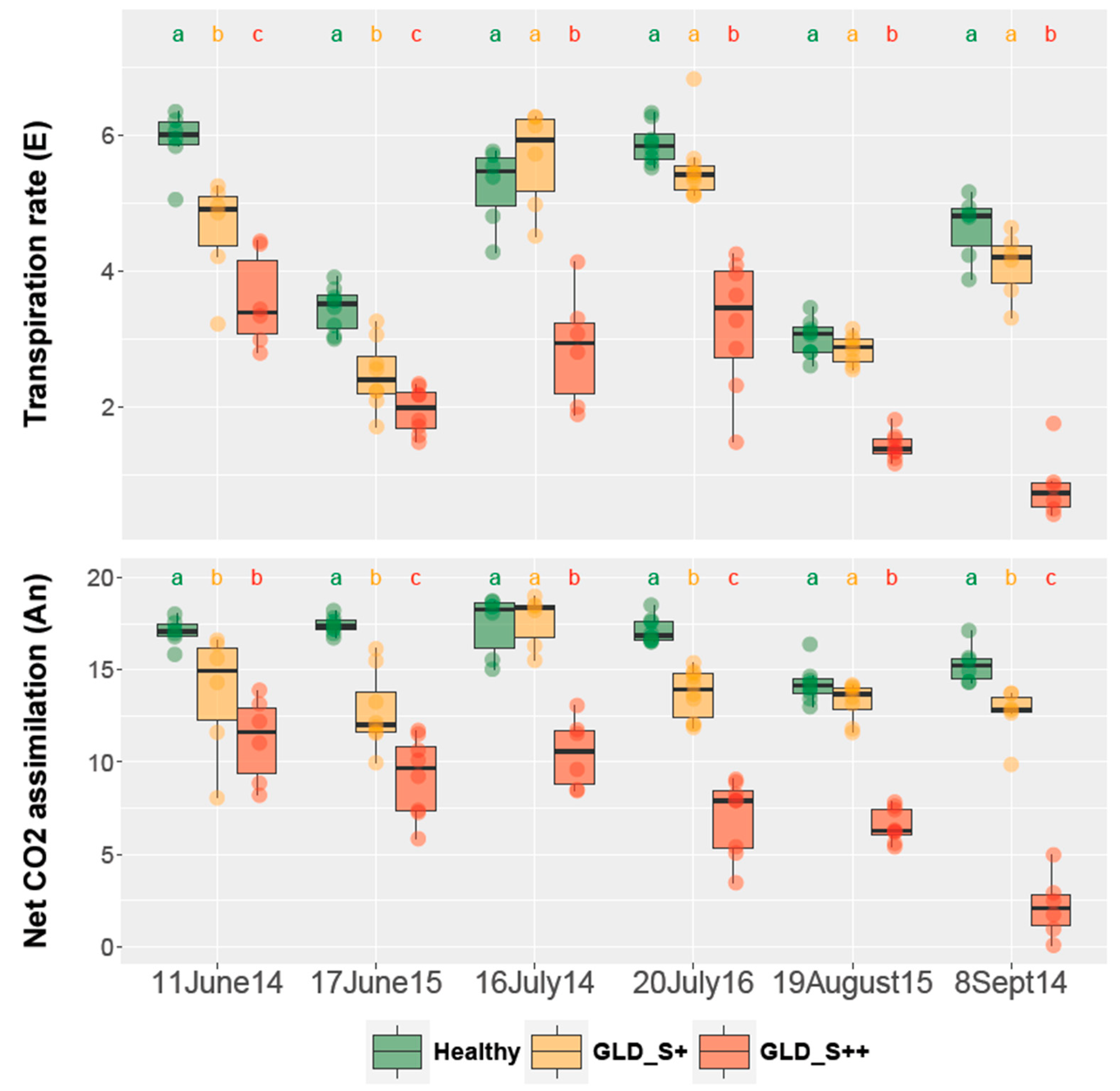
2.2. Vine Physiology and Fruit Composition
2.3. Virome Characterization
2.4. Mealybug Transmission Experiment
3. Results
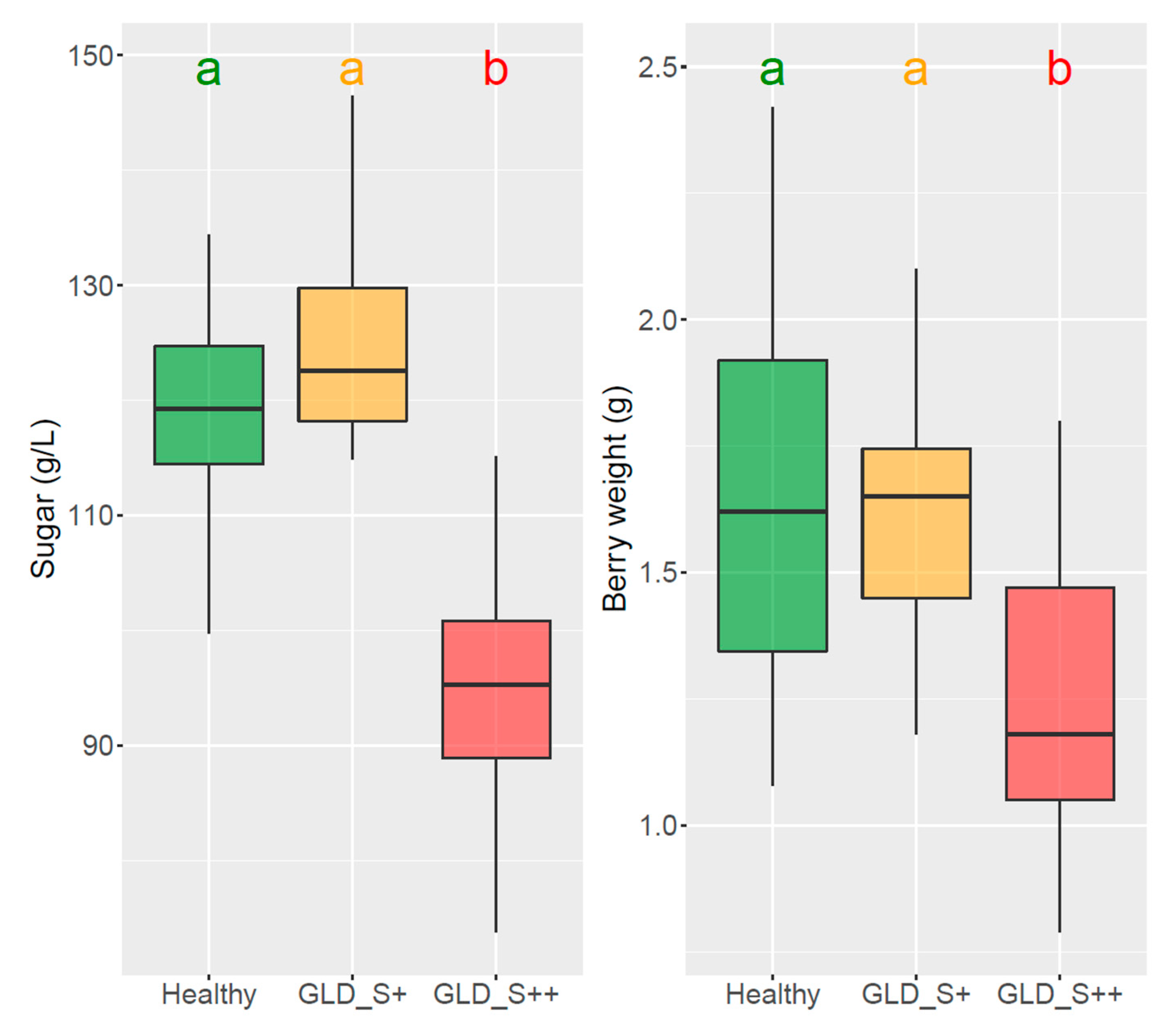
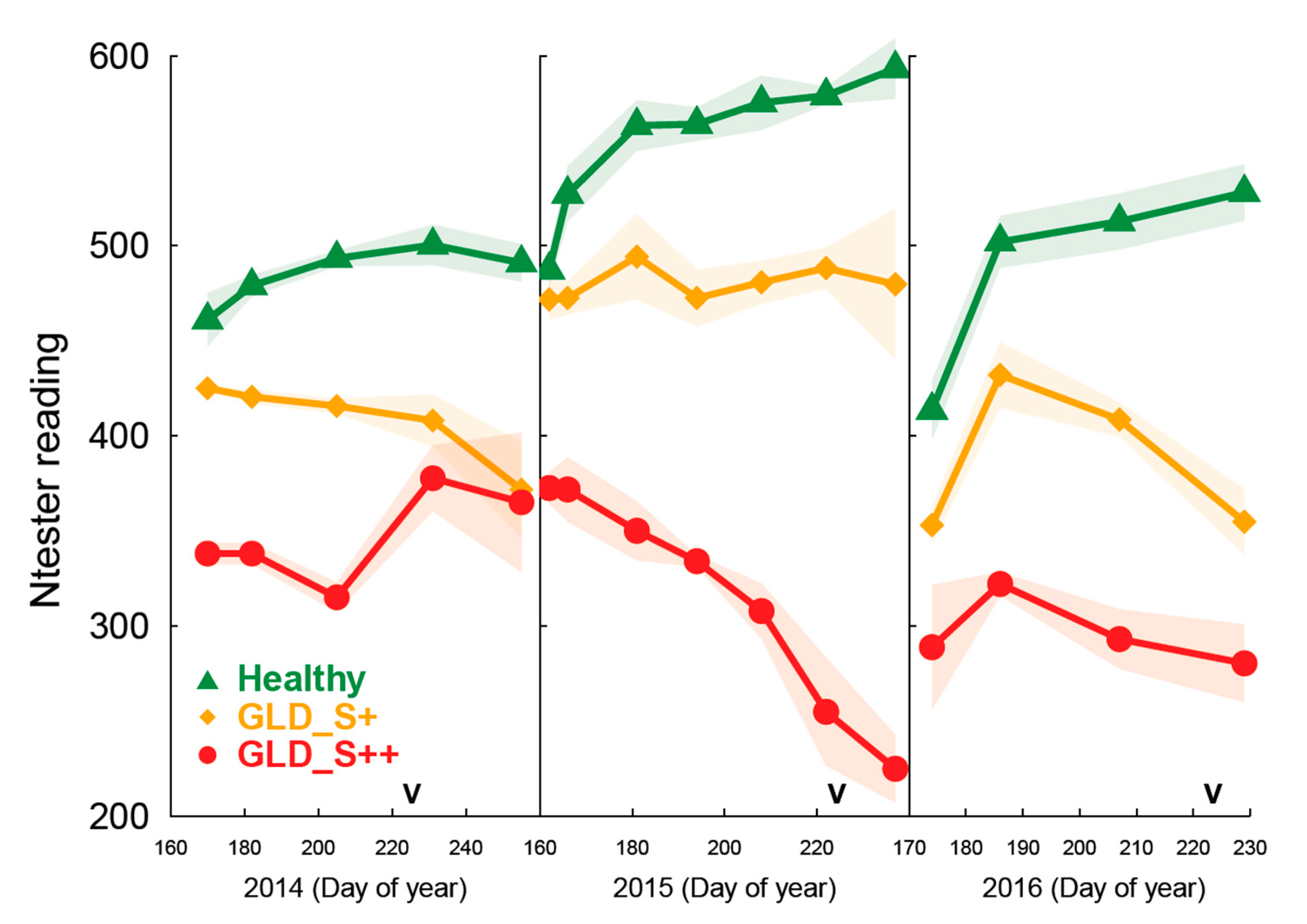
3.1. Symptoms Development, Agronomical Impact and Effects on Fruit Composition
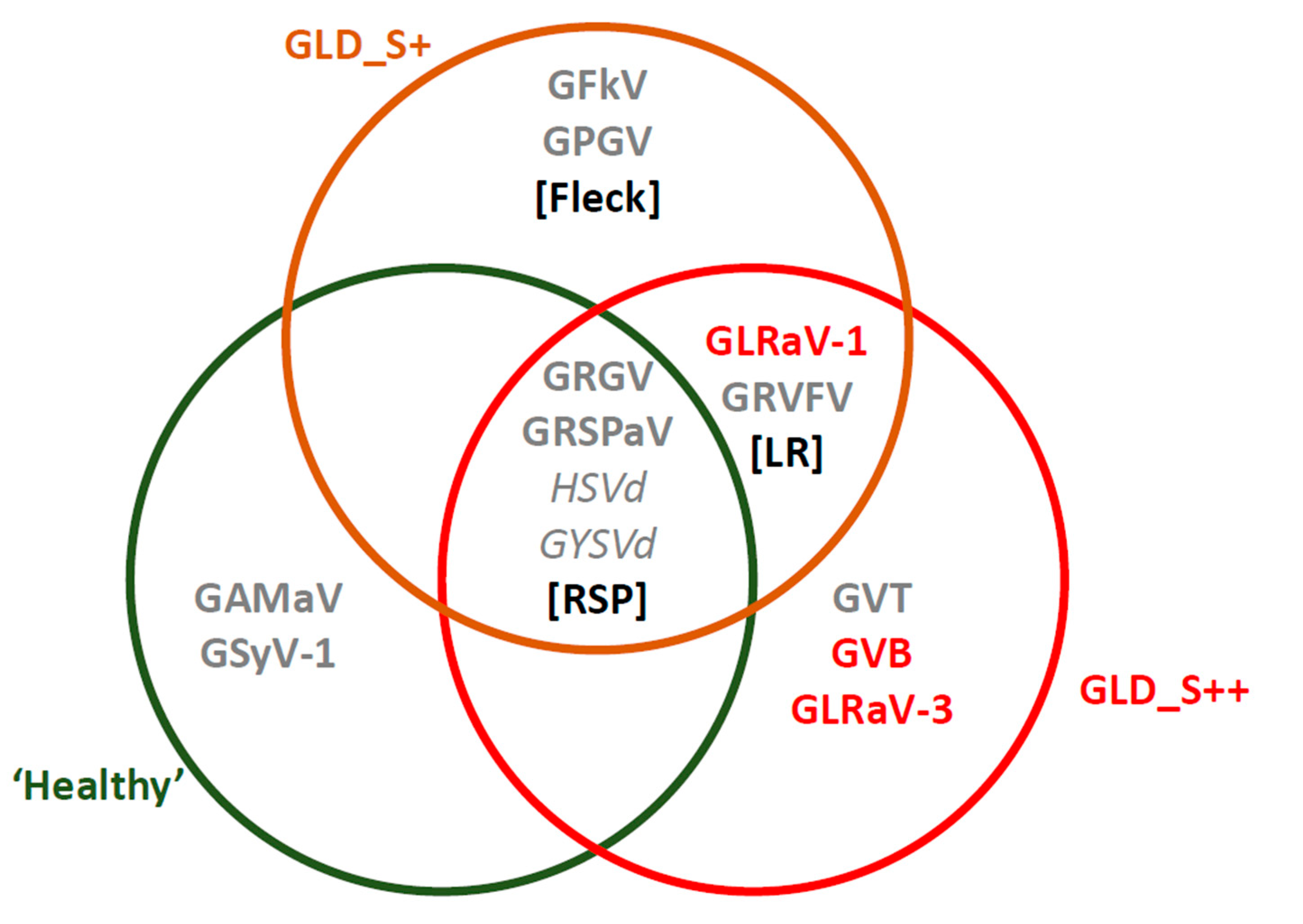
3.2. Virome Characterization
3.3. Mealybug Transmission Experiment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McGovern, P.; Jalabadze, M.; Batiuk, S.; Callahan, M.P.; Smith, K.E.; Hall, G.R.; Kvavadze, E.; Maghradze, D.; Rusishvili, N.; Bouby, L.; et al. Early Neolithic wine of Georgia in the South Caucasus. Proc. Natl. Acad. Sci. USA 2017, 114, E10309–E10318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, A. The grapevine, viticulture, and winemaking: A brief introduction. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Springer: Cham, Switzerland, 2017; pp. 3–29. [Google Scholar]
- Meng, B.; Rowhani, A. Grapevine rupestris stem pitting-associated virus. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Springer: Cham, Switzerland, 2017; pp. 257–287. [Google Scholar] [CrossRef]
- Hily, J.-M.; Beuve, M.; Vigne, E.; Demangeat, G.; Candresse, T.; Lemaire, O. A genome-wide diversity study of grapevine rupestris stem pitting-associated virus. Arch. Virol. 2018, 163, 3105–3111. [Google Scholar] [CrossRef] [PubMed]
- Naidu, R.A.; Maree, H.J.; Burger, J.T. Grapevine Leafroll Disease and Associated Viruses: A Unique Pathosystem. Annu. Rev. Phytopathol. 2015, 53, 613–634. [Google Scholar] [CrossRef] [PubMed]
- Maree, H.J.; Almeida, R.P.; Bester, R.; Chooi, K.M.; Cohen, D.; Dolja, V.V.; Fuchs, M.F.; Golino, D.A.; Jooste, A.E.; Martelli, G.P. Grapevine leafroll-associated virus 3. Front. Microbiol. 2013, 4, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alabi, O.; Casassa, L.F.; Gutha, L.R.; Larsen, R.C.; Henick-Kling, T.; Harbertson, J.F.; Naidu, R.A. Impacts of Grapevine Leafroll Disease on Fruit Yield and Grape and Wine Chemistry in a Wine Grape (Vitis vinifera L.) Cultivar. PLoS ONE 2016, 11, e0149666. [Google Scholar] [CrossRef] [Green Version]
- Ghaffari, S.; Reynard, J.S.; Rienth, M. Single berry reconstitution prior to RNA-sequencing reveals novel insights into transcriptomic remodeling by leafroll virus infections in grapevines. Sci. Rep. 2020, 10, 12905. [Google Scholar] [CrossRef]
- Hou, W.; Li, S.; Massart, S. Is There a “Biological Desert” With the Discovery of New Plant Viruses? A Retrospective Analysis for New Fruit Tree Viruses. Front. Microbiol. 2020, 11, 2953. [Google Scholar] [CrossRef]
- Massart, S.; Candresse, T.; Gil, J.F.; Lacomme, C.; Predajna, L.; Ravnikar, M.; Reynard, J.-S.; Rumbou, A.; Saldarelli, P.; Škorić, D.; et al. A Framework for the Evaluation of Biosecurity, Commercial, Regulatory, and Scientific Impacts of Plant Viruses and Viroids Identified by NGS Technologies. Front. Microbiol. 2017, 8, 45. [Google Scholar] [CrossRef] [Green Version]
- Zufferey, V.; Cochard, H.; Ameglio, T.; Spring, J.L.; Viret, O. Diurnal cycles of embolism formation and repair in petioles of grapevine (Vitis vinifera cv. Chasselas). J. Exp. Bot. 2011, 62, 3885–3894. [Google Scholar] [CrossRef] [Green Version]
- Ribéreau-Gayon, P.; Glories, Y.; Maujean, A.; Dubourdieu, D. Handbook of Enology, Volume 2: The Chemistry of Wine Stabilization and Treatments; John Wiley & Sons: Hoboken, NJ, USA, 2006. [Google Scholar]
- de Mendiburu, F.; de Mendiburu, M.F. Package ‘agricolae’. R Package Version 2019, 1, 1–2. [Google Scholar]
- Reynard, J.-S.; Schaerer, S. Virus, bactéries et phytoplasmes de la vigne. Rev. Suisse De Vitic. Arboric. Et Hortic. 2019, 51, 168–172. [Google Scholar]
- Gugerli, P.; Brugger, J.-J.; Ramel, M.-E.; Besse, S. In Grapevine virus collection at Nyon: A contribution to a putative network of a worldwide grapevine virus reference collection. In Proceedings of the 16th Meeting of the International Council for the Stufy of Virus and Virus-Like Disease of Grapevine, Dijon, France, 31 August–4 September 2009; pp. 40–41. [Google Scholar]
- Correa, M.; Germain, J.; Malausa, T.; Zaviezo, T. Molecular and morphological characterization of mealybugs (Hemiptera: Pseudococcidae) from Chilean vineyards. Bull. Entomol. Res. 2012, 102, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Le Maguet, J.; Beuve, M.; Herrbach, E.; Lemaire, O. Transmission of Six Ampeloviruses and Two Vitiviruses to Grapevine by Phenacoccus aceris. Phytopathology 2012, 102, 717–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, V.A.; Chooi, K.M.; Blouin, A.G.; Pierersen, G.; MacDiarmid, R.M. In Timing of the symptoms influences the management strategies of Grapevine leafroll-associate virus 3 in New Zealand. In Proceedings of the 18th Congress of the International Council for the Study of Virus and Virus-like Diseases of the Grapevine (ICVG), Ankara, Turkey, 7–11 September 2015; pp. 40–41. [Google Scholar]
- Chooi, K.; Bell, B.A.; Blouin, A.; Cohen, D.; Mundy, D.; Henshall, W.; MacDiarmid, R.M. Grapevine leafroll-associated virus 3 genotype influences foliar symptom development in New Zealand vineyards. Viruses 2022, (in press). [Google Scholar]
- Diaz-Lara, A.; Klaassen, V.; Stevens, K.; Sudarshana, M.R.; Rowhani, A.; Maree, H.J.; Chooi, K.M.; Blouin, A.G.; Habili, N.; Song, Y. Characterization of grapevine leafroll-associated virus 3 genetic variants and application towards RT-qPCR assay design. PLoS ONE 2018, 13, e0208862. [Google Scholar] [CrossRef] [Green Version]
- Poojari, S.; Boulé, J.; Delury, N.; Lowery, D.T.; Rott, M.; Schmidt, A.-M.; Úrbez-Torres, J.R. Epidemiology and Genetic Diversity of Grapevine Leafroll-Associated Viruses in British Columbia. Plant Dis. 2017, 101, 2088–2097. [Google Scholar] [CrossRef] [Green Version]
- Gouveia, P.A.O.; Nolasco, G. The p19.7 RNA silencing suppressor from Grapevine leafroll-associated virus 3 shows different levels of activity across phylogenetic groups. Virus Genes 2012, 45, 333–339. [Google Scholar] [CrossRef]
- Pandey, N.; Ahmad, A.; Chooi, K.M.; Blouin, A.G.; Ziebell, H.; MacDiarmid, R.M. PP06: Characterising the suppressors of silencing encoded by Grapevine leafroll-associated virus 3 and their activity in New Zealand genetic variants. In Proceedings of the 18th Congress of ICVG, Ankara, Turkey, 7–11 September 2015; pp. 159–160. [Google Scholar]
- Ghosh, D.; Chakraborty, S. Impact of viral silencing suppressors on plant viral synergism: A global agro-economic concern. Appl. Microbiol. Biotechnol. 2021, 105, 6301–6313. [Google Scholar] [CrossRef]
- Rowhani, A.; Daubert, S.; Arnold, K.; Al Rwahnih, M.; Klaassen, V.; Golino, D.; Uyemoto, J.K. Synergy between grapevine vitiviruses and grapevine leafroll viruses. Eur. J. Plant Pathol. 2018, 151, 919–925. [Google Scholar] [CrossRef]
- Minafra, A.; Mawassi, M.; Goszczynski, D.; Saldarelli, P. Grapevine vitiviruses. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Springer: Cham, Switzerland, 2017; pp. 229–256. [Google Scholar]
- Maree, H.J.; Blouin, A.G.; Diaz-Lara, A.; Mostert, I.; Al Rwahnih, M.; Candresse, T. Status of the current vitivirus taxonomy. Arch. Virol. 2019, 165, 451–458. [Google Scholar] [CrossRef]
- Blaisdell, G.K.; Zhang, S.; Rowhani, A.; Klaassen, V.; Cooper, M.L.; Daane, K.M.; Almeida, R.P.P. Trends in vector-borne transmission efficiency from coinfected hosts: Grapevine leafroll-associated virus-3 and Grapevine virus A. Eur. J. Plant Pathol. 2020, 156, 1163–1167. [Google Scholar] [CrossRef]
- Greenspan, M.D.; Schultz, H.R.; Matthews, M.A. Field evaluation of water transport in grape berries during water deficits. Physiol. Plant. 1996, 97, 55–62. [Google Scholar] [CrossRef]
- Giribaldi, M.; Purrotti, M.; Pacifico, D.; Santini, D.; Mannini, F.; Caciagli, P.; Rolle, L.; Cavallarin, L.; Giuffrida, M.G.; Marzachì, C. A multidisciplinary study on the effects of phloem-limited viruses on the agronomical performance and berry quality of Vitis vinifera cv. Nebbiolo. J. Proteom. 2011, 75, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Santini, D.; Rolle, L.; Cascio, P.; Mannini, F. Modifications in Chemical, Physical and Mechanical Properties of Nebbiolo (Vitis vinifera L.) Grape Berries Induced by Mixed Virus Infection. South Afr. J. Enol. Vitic. 2011, 32, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Rienth, M.; Vigneron, N.; Walker, R.P.; Castellarin, S.D.; Sweetman, C.; Burbidge, C.A.; Bonghi, C.; Famiani, F.; Darriet, P. Modifications of Grapevine Berry Composition Induced by Main Viral and Fungal Pathogens in a Climate Change Scenario. Front. Plant Sci. 2021, 12, 717223. [Google Scholar] [CrossRef]
- Mannini, F.; Mollo, A.; Credi, R. Field Performance and Wine Quality Modification in a Clone of Vitis vinifera cv. Dolcetto after GLRaV-3 Elimination. Am. J. Enol. Vitic. 2012, 63, 144–147. [Google Scholar] [CrossRef] [Green Version]
- Moutinho-Pereira, J.; Correia, C.; Gonçalves, B.; Bacelar, E.; Coutinho, J.; Ferreira, H.M.F.; Lousada, J.; Cortez, I. Impacts of leafroll-associated viruses (GLRaV-1 and -3) on the physiology of the Portuguese grapevine cultivar ‘Touriga Nacional’ growing under field conditions. Ann. Appl. Biol. 2012, 160, 237–249. [Google Scholar] [CrossRef]
- Montero, R.; Mundy, D.; Albright, A.; Grose, C.; Trought, M.; Cohen, D.; Chooi, K.; MacDiarmid, R.; Flexas, J.; Bota, J. Effects of Grapevine Leafroll associated Virus 3 (GLRaV-3) and duration of infection on fruit composition and wine chemical profile of Vitis vinifera L. cv. Sauvignon blanc. Food Chem. 2016, 197, 1177–1183. [Google Scholar] [CrossRef]
- Bell, S.J.; Henschke, P.A. Implications of nitrogen nutrition for grapes, fermentation and wine. Aust. J. Grape Wine Res. 2005, 11, 242–295. [Google Scholar] [CrossRef]
- Lee, J.; Keller, K.E.; Rennaker, C.; Martin, R.R. Influence of grapevine leafroll associated viruses (GLRaV-2 and-3) on the fruit composition of Oregon Vitis vinifera L. cv. Pinot noir: Free amino acids, sugars, and organic acids. Food Chem. 2009, 117, 99–105. [Google Scholar] [CrossRef]
- Cabaleiro, C.; Pesqueira, A.; García-Berrios, J. Influence of Grapevine Leafroll-associated Virus-3 in Mature Plants of Vitis vinifera L. cv Albariño on 110R and 196.17C Rootstocks. South Afr. J. Enol. Vitic. 2021, 42, 165–174. [Google Scholar] [CrossRef]
- Golino, D.A.; Wolpert, J.; Sim, B.J.; Aderson, R.A. Virus effects on vine growth and fruit components of Cabernet Sauvignon on six rootstocks. In Proceedings of the Second Annual National Viticulture Research Conference, University of California, Davis, CA, USA, 9–11 July 2008; pp. 9–11. [Google Scholar]
- Komar, V.; Vigne, E.; Demangeat, G.; Lemaire, O.; Fuchs, M. Comparative performance of virus-infected Vitis vinifera cv. Savagnin rose grafted onto three rootstocks. Am. J. Enol. Vitic. 2010, 61, 68–73. [Google Scholar]
- Rosa, C.; Jimenez, J.F.; Margaria, P.; Rowhani, A. Symptomatology and Effects of Viruses Associated with Rugose Wood Complex on the Growth of Four Different Rootstocks. Am. J. Enol. Vitic. 2011, 62, 207–213. [Google Scholar] [CrossRef]
- Vondras, A.M.; Lerno, L.; Massonnet, M.; Minio, A.; Rowhani, A.; Liang, D.; Garcia, J.; Quiroz, D.; Figueroa-Balderas, R.; Golino, D.A.; et al. Rootstock influences the effect of grapevine leafroll-associated viruses on berry development and metabolism via abscisic acid signalling. Mol. Plant Pathol. 2021, 22, 984–1005. [Google Scholar] [CrossRef] [PubMed]
- Rowhani, A.; Golino, D.A.; Klaassen, V.; Sim, S.T.; Gouran, M.; Al Rwahnih, M. In Grapevine Leafroll associated virus 3: Effects on rootstocks, vine performance, yield and berries. In Proceedings of the 18th Congress of ICVG, Ankara, Turkey, 7–11 September 2015; pp. 161–162. [Google Scholar]
- Golino, D.A.; Rowhani, A.; Klaassen, V.; Sim, S.T.; Al Rwahnih, M. Grapevine leafroll associated virus 1 effects on different grapevine rootstocks. In Proceedings of the 18th Congress of ICVG, Ankara, Turkey, 7–11 September 2015; pp. 46–47. [Google Scholar]
- Spring, J.-L.; Ferretti, M.; Zufferey, V.; Verdenal, T.; Viret, O. Influence du porte-greffe sur le comportement du cépage Gamaret dans le vignoble tessinois. Rev. Suisse De Vitic. Arboric. Et Hortic. 2013, 45, 158–164. [Google Scholar]
- Anonymous. Ordonnance du DFE (916.151.3) sur la Production et la Mise en Circulation de Matériel de Multiplication de la Vigne (Ordonnance du DFE sur les Plants de Vigne). In L’économie; le Département Fédéral de L’économie, de la Formation et de la Recherche (DEFR). 2006. Available online: https://fedlex.data.admin.ch/filestore/fedlex.data.admin.ch/eli/cc/2006/789/20120701/fr/pdf-a/fedlex-data-admin-ch-eli-cc-2006-789-20120701-fr-pdf-a.pdf (accessed on 17 June 2022).
- EPPO. Pathogen-tested material of grapevine varieties and rootstocks. EPPO Bull. 2008, 38, 422–429. [Google Scholar] [CrossRef]
- Golino, D.A.; Fuchs, M.; Sim, S.; Farrar, K.; Martelli, G.P. Improvement of Grapevine Planting Stock through Sanitary Selection and Pathogen Elimination. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Springer: Cham, Switzerland, 2017; pp. 561–579. [Google Scholar] [CrossRef]
- Golino, D.A.; Fuchs, M.; Al Rwahnih, M.; Farrar, K.; Schmidt, A.; Martelli, G.P. Regulatory Aspects of Grape Viruses and Virus Diseases: Certification, Quarantine, and Harmonization. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Springer: Cham, Switzerland, 2017; pp. 581–598. [Google Scholar] [CrossRef]
- Refatti, E. Grapevine asteroid mosaic. In Proceedings of the International Conference on Virus and Vector on Perennial Hosts with Special Referemceto Vitis, Davis, CA, USA, 6–10 September 1965; pp. 157–164. [Google Scholar]
- Sabanadzovic, S.; Abou-Ghanem, N.; Castellano, M.A.; Digiaro, M.; Martelli, G.P. Grapevine fleck virus-like viruses in Vitis. Arch. Virol. 2000, 145, 553–565. [Google Scholar] [CrossRef]
- Goszczynski, D.E. Divergent molecular variants of Grapevine virus B (GVB) from corky bark (CB)-affected and CB-negative LN33 hybrid grapevines. Virus Genes 2010, 41, 273–281. [Google Scholar] [CrossRef]
- Rowhani, A.; Uyemoto, J.K.; Golino, D.A.; Daubert, S.D.; Al Rwahnih, M. Viruses Involved in Graft Incompatibility and Decline. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Springer: Cham, Switzerland, 2017; pp. 289–302. [Google Scholar] [CrossRef]
- Al Rwahnih, M.; Daubert, S.; Golino, D.; Islas, C.; Rowhani, A. Comparison of Next-Generation Sequencing Versus Biological Indexing for the Optimal Detection of Viral Pathogens in Grapevine. Phytopathology 2015, 105, 758–763. [Google Scholar] [CrossRef] [Green Version]
- Rott, M.; Xiang, Y.; Boyes, I.; Belton, M.; Saeed, H.; Kesanakurti, P.; Hayes, S.; Lawrence, T.; Birch, C.; Bhagwat, B. Application of next generation sequencing for diagnostic testing of tree fruit viruses and viroids. Plant Dis. 2017, 101, 1489–1499. [Google Scholar] [CrossRef] [Green Version]
- Sebastien, M.; Adams, I.; Al Rwahnih, M.; Baeyen, S.; Bilodeau, G.J.; Blouin, A.; Boonham, N.; Candresse, T.; Chandelier, A.; De Jonghe, K.; et al. Guidelines for the reliable use of high throughput sequencing technologies to detect plant pathogens and pests. Peer Community J. 2022. submitted. [Google Scholar] [CrossRef]
- Lebas, B.S.M.; Adams, I.; Al Rwahnih, M.; Baeyen, S.; Bilodeau, G.; Blouin, A.G.; Boonham, N.; Candresse, T.; Chandelier, A.; De Jonghe, K.; et al. Guidelines for preparing a laboratory to use high-throughput sequencing as routine diagnostic test: General and technical recommendations. EPPO Bull. 2022. submitted. [Google Scholar]
- Osman, F.; Rowhani, A. Application of a spotting sample preparation technique for the detection of pathogens in woody plants by RT-PCR and real-time PCR (TaqMan). J. Virol. Methods 2006, 133, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Beuve, M.; Moury, B.; Spilmont, A.-S.; Sempé-Ignatovic, L.; Hemmer, C.; Lemaire, O. Viral sanitary status of declining grapevine Syrah clones and genetic diversity of Grapevine Rupestris stem pitting-associated virus. Eur. J. Plant Pathol. 2013, 135, 439–452. [Google Scholar] [CrossRef]
- Levy, L.; Lee, M.; Hadidi, A. Simple and rapid preparation of infected plant tissue extracts for PCR amplification of virus, viroid, and MLO nucleic acids. J. Virol. Methods 1994, 49, 295–304. [Google Scholar] [CrossRef]
- Saldarelli, P.; Giampetruzzi, A.; Morelli, M.; Malossini, U.; Pirolo, C.; Bianchedi, P.; Gualandri, V. Genetic variability of Grapevine Pinot gris virus and its association with grapevine leaf mottling and deformation. Phytopathology 2015, 105, 555–563. [Google Scholar] [CrossRef] [Green Version]
- Terlizzi, F.; Li, C.; Ratti, C.; Qiu, W.; Credi, R.; Meng, B. Detection of multiple sequence variants of Grapevine rupestris stem pitting-associated virus using primers targeting the polymerase domain and partial genome sequencing of a novel variant. Ann. Appl. Biol. 2011, 159, 478–490. [Google Scholar] [CrossRef]
- Beuve, M.; Candresse, T.; Tannières, M.; Lemaire, O. First report of grapevine redglobe virus (GRGV) in grapevine in France. Plant Dis. 2015, 99, 422. [Google Scholar] [CrossRef]
- Glasa, M.; Predajňa, L.; Komínek, P. Grapevine fleck virus isolates split into two distinct molecular groups. J. Phytopathol. 2011, 159, 805–807. [Google Scholar]
- Xiao, H.; Meng, B. First report of Grapevine asteroid mosaic-associated virus and Grapevine rupestris vein feathering virus in Grapevines in Canada. Plant Dis. 2016, 100, 2175. [Google Scholar] [CrossRef]
- Al Rwahnih, M.; Daubert, S.; Golino, D.; Rowhani, A. Deep sequencing analysis of RNAs from a grapevine showing Syrah decline symptoms reveals a multiple virus infection that includes a novel virus. Virology 2009, 387, 395–401. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2015 | 2016 | 2017 | 2018 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rootsock | ||||||||||||||||
| Treatment | 5BB | % * | 3309C | % * | 5BB | % * | 3309C | % * | 5BB | % * | 3309C | % * | 5BB | % * | 3309C | % * |
| Symptomless | 460 a | 338 a | 480 a | 305 a | 391 a | 220 a | 540 a | 399 a | ||||||||
| GLD_+ | 403 a | −12 | 296 b | −12 | 380 b | −20 | 263 a | −13 | 295 b | −20 | 195 a | −12 | 490 a | −9 | 349 a | −12 |
| GLD_S++ | 199 b | −43 | 64 c | −81 | 211 c | −56 | 54 b | −83 | 159 c | −59 | 64 b | −62 | 205 b | −62 | 88 b | −78 |
| Rootstock | ||||||
|---|---|---|---|---|---|---|
| Treatment | 5BB | 3309C | ||||
| Mortality | Trunk Diameter (cm) | % * | Mortality | Trunk Diameter (cm) | % * | |
| Symptomless | 0/10 | 3.5 a | 0/10 | 3.1 a | ||
| GLD_S+ | 0/10 | 3.2 b | −9 | 0/10 | 2.8 b | −10 |
| GLD_S++ | 0/10 | 2.4 c | −31 | 4/10 | 1.8 c | −42 |
| Yield (kg/vine) | Soluble Solids (°Brix) | pH | Total Acidity | Tartaric Acid (g/L) | YAN | |
|---|---|---|---|---|---|---|
| 2015 | ||||||
| Symptomless | 0.6 a | 22.2 a | 3.12 a | 8.6 a | 7.9 a | 178 a |
| GLD_S+ | 0.5 a | 22.5 a | 3.10 a | 8.3 a | 7.5 a | 132 a |
| GLD_S++ | 0.1 b | 22.4 a | 3.33 b | 6.8 b | 6.4 b | 65 b |
| 2016 | ||||||
| Symptomless | 1.0 a | 24.1 a | 3.23 a | 8.0 a | 6.4 a | 151 a |
| GLD_S+ | 1.1 a | 23.3 a | 3.12 b | 8.4 a | 6.7 a | 111 ab |
| GLD_S++ | 0.2 b | 19.4 b | 3.25 a | 7.8 a | 6.1 a | 66 b |
| 2017 | ||||||
| Symptomless | 0.6 a | 22.5 a | 3.23 a | 8.7 a | 6.5 a | 245 a |
| GLD_S+ | 0.3 b | 22.4 a | 3.23 a | 8.8 a | 6.5 a | 178 b |
| GLD_S++ | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reynard, J.-S.; Brodard, J.; Zufferey, V.; Rienth, M.; Gugerli, P.; Schumpp, O.; Blouin, A.G. Nuances of Responses to Two Sources of Grapevine Leafroll Disease on Pinot Noir Grown in the Field for 17 Years. Viruses 2022, 14, 1333. https://0-doi-org.brum.beds.ac.uk/10.3390/v14061333
Reynard J-S, Brodard J, Zufferey V, Rienth M, Gugerli P, Schumpp O, Blouin AG. Nuances of Responses to Two Sources of Grapevine Leafroll Disease on Pinot Noir Grown in the Field for 17 Years. Viruses. 2022; 14(6):1333. https://0-doi-org.brum.beds.ac.uk/10.3390/v14061333
Chicago/Turabian StyleReynard, Jean-Sébastien, Justine Brodard, Vivian Zufferey, Markus Rienth, Paul Gugerli, Olivier Schumpp, and Arnaud G. Blouin. 2022. "Nuances of Responses to Two Sources of Grapevine Leafroll Disease on Pinot Noir Grown in the Field for 17 Years" Viruses 14, no. 6: 1333. https://0-doi-org.brum.beds.ac.uk/10.3390/v14061333