
Domestic Cat Hepadnavirus: Molecular Epidemiology and Phylogeny in Cats in Hong Kong

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Sample and Data Collection
2.3. Sample Processing
2.4. Real-Time Quantitative PCR (qPCR)
2.5. Data Analysis
2.6. Whole Genome Sequencing
2.7. Phylogenetic Analyses
2.8. Viral DNA Sequence Analyses
3. Results
3.1. Molecular Epidemiology of DCH in Cats in Hong Kong
3.2. Phylogenetic Analysis Positions Hong Kong DCH Isolates in Clade A1 with South-East Asian and Australian Strains
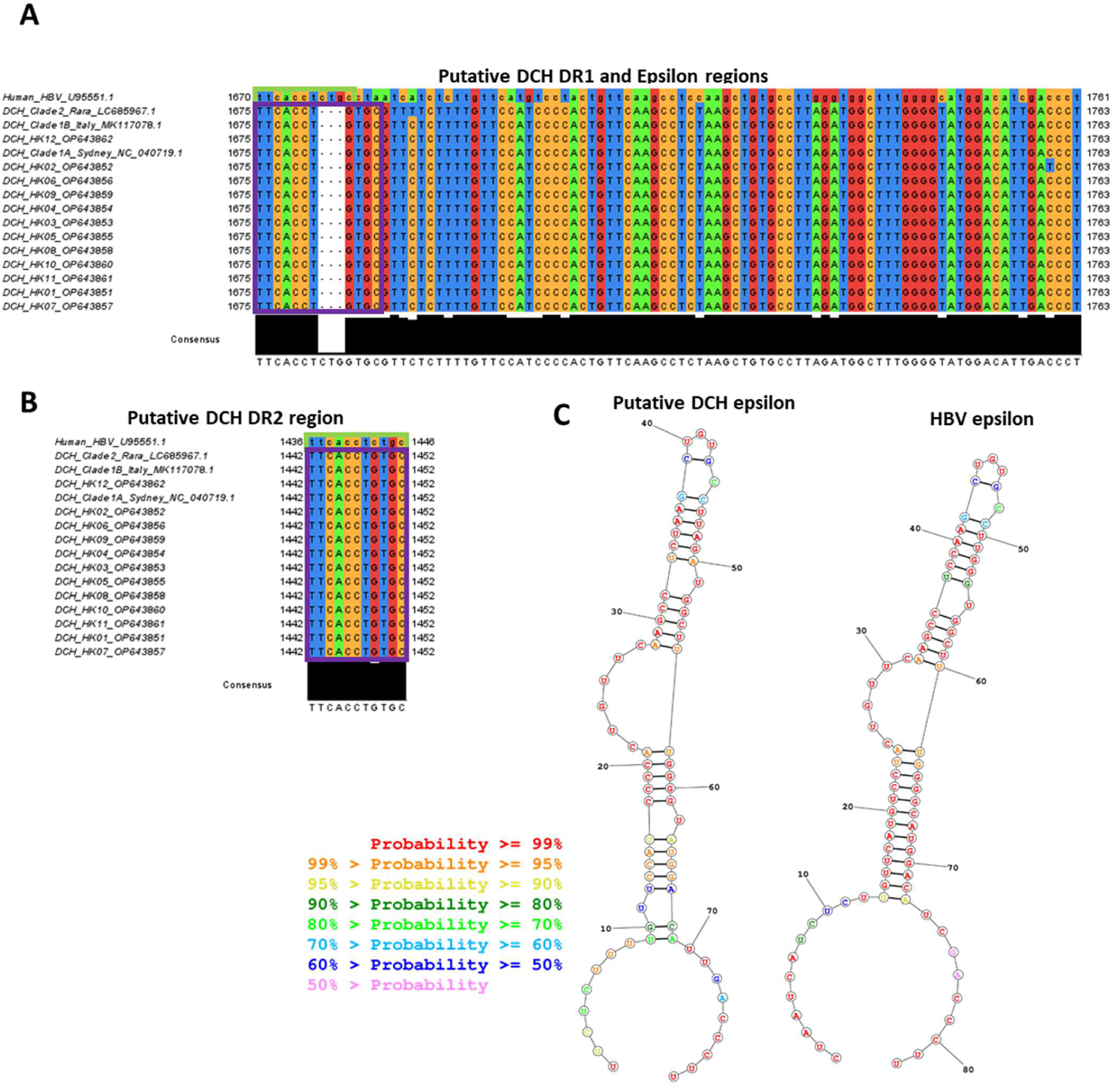
3.3. Sequence Analysis Shows Hong Kong DCH Isolates Have Conserved Features Common to Other DCH Isolates
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Magnius, L.; Mason, W.S.; Taylor, J.; Kann, M.; Glebe, D.; Dény, P.; Sureau, C.; Norder, H.; Consortium, I.R. ICTV Virus Taxonomy Profile: Hepadnaviridae. J. Gen. Virol. 2020, 101, 571–572. [Google Scholar] [CrossRef] [PubMed]
- Sheena, B.S.; Hiebert, L.; Han, H.; Ippolito, H.; Abbasi-Kangevari, M.; Abbasi-Kangevari, Z.; Abbastabar, H.; Abdoli, A.; Ali, H.A.; Adane, M.M.; et al. Global, regional, and national burden of hepatitis B, 1990–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet Gastroenterol. Hepatol. 2022, 7, 796–829. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Global Progress Report on HIV, Viral Hepatitis and Sexually Transmitted Infections, 2021: Accountability for the Global Health Sector Strategies 2016–2021: Actions for Impact: Web Annex 2: Data Methods; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- CDA Foundation Polaris Observatory. Available online: https://cdafound.org/polaris-regions-compare/ (accessed on 19 December 2022).
- Menne, S.; Cote, P.J. The woodchuck as an animal model for pathogenesis and therapy of chronic hepatitis B virus infection. World J. Gastroenterol. 2007, 13, 104–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aghazadeh, M.; Shi, M.; Barrs, V.R.; McLuckie, A.J.; Lindsay, S.A.; Jameson, B.; Hampson, B.; Holmes, E.C.; Beatty, J.A. A Novel Hepadnavirus Identified in an Immunocompromised Domestic Cat in Australia. Viruses 2018, 10, 269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanave, G.; Capozza, P.; Diakoudi, G.; Catella, C.; Catucci, L.; Ghergo, P.; Stasi, F.; Barrs, V.; Beatty, J.; Decaro, N.; et al. Identification of hepadnavirus in the sera of cats. Sci. Rep. 2019, 9, 10668. [Google Scholar] [CrossRef] [Green Version]
- Piewbang, C.; Wardhani, S.W.; Chaiyasak, S.; Yostawonkul, J.; Chai-In, P.; Boonrungsiman, S.; Kasantikul, T.; Techangamsuwan, S. Insights into the genetic diversity, recombination, and systemic infections with evidence of intracellular maturation of hepadnavirus in cats. PLoS ONE 2020, 15, e0241212. [Google Scholar] [CrossRef]
- Anpuanandam, K.; Selvarajah, G.T.; Choy, M.M.K.; Ng, S.W.; Kumar, K.; Ali, R.M.; Rajendran, S.K.; Ho, K.L.; Tan, W.S. Molecular detection and characterisation of Domestic Cat Hepadnavirus (DCH) from blood and liver tissues of cats in Malaysia. BMC Vet. Res. 2021, 17, 9. [Google Scholar] [CrossRef]
- Jeanes, E.C.; Wegg, M.L.; Mitchell, J.A.; Priestnall, S.L.; Fleming, L.; Dawson, C. Comparison of the prevalence of Domestic Cat Hepadnavirus in a population of cats with uveitis and in a healthy blood donor cat population in the United Kingdom. Vet. Ophthalmol. 2022, 25, 165–172. [Google Scholar] [CrossRef]
- Stone, C.; Petch, R.; Gagne, R.B.; Nehring, M.; Tu, T.; Beatty, J.A.; VandeWoude, S. Prevalence and Genomic Sequence Analysis of Domestic Cat Hepadnavirus in the United States. Viruses 2022, 14, 2091. [Google Scholar] [CrossRef]
- Takahashi, K.; Kaneko, Y.; Shibanai, A.; Yamamoto, S.; Katagiri, A.; Osuga, T.; Inoue, Y.; Kuroda, K.; Tanabe, M.; Okabayashi, T.; et al. Identification of domestic cat hepadnavirus from a cat blood sample in Japan. J. Vet. Med. Sci. 2022, 84, 648–652. [Google Scholar] [CrossRef]
- Fruci, P.; Di Profio, F.; Palombieri, A.; Massirio, I.; Lanave, G.; Diakoudi, G.; Pellegrini, F.; Marsilio, F.; Martella, V.; Di Martino, B. Detection of antibodies against domestic cat hepadnavirus using baculovirus-expressed core protein. Transbound. Emerg. Dis. 2022, 69, 2980–2986. [Google Scholar] [CrossRef]
- Pesavento, P.A.; Jackson, K.; Hampson, T.; Munday, J.S.; Barrs, V.R.; Beatty, J.A. A Novel Hepadnavirus is Associated with Chronic Hepatitis and Hepatocellular Carcinoma in Cats. Viruses 2019, 11, 969. [Google Scholar] [CrossRef] [Green Version]
- Piewbang, C.; Dankaona, W.; Poonsin, P.; Yostawonkul, J.; Lacharoje, S.; Sirivisoot, S.; Kasantikul, T.; Tummaruk, P.; Techangamsuwan, S. Domestic cat hepadnavirus associated with hepatopathy in cats: A retrospective study. J. Vet. Intern. Med. 2022, 36, 1648–1659. [Google Scholar] [CrossRef]
- Peterson, M.E.; Ward, C.R. Etiopathologic Findings of Hyperthyroidism in Cats. Vet. Clin. N. Am. Small Anim. Pract. 2007, 37, 633–645. [Google Scholar] [CrossRef]
- McLuckie, A.; Barrs, V.; Wilson, B.; Westman, M.; Beatty, J. Felis Catus Gammaherpesvirus 1 DNAemia in Whole Blood from Therapeutically Immunosuppressed or Retrovirus-Infected Cats. Vet. Sci. 2017, 4, 16. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, M.; Kishino, H.; Yano, T. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef]
- Madeira, F.; Pearce, M.; Tivey, A.R.N.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and sequence analysis tools services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.M.; Procter, J.B.; Martin DM, A.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reuter, J.S.; Mathews, D.H. RNA structure: Software for RNA secondary structure prediction and analysis. BMC Bioinform. 2010, 11, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scavone, D.; Lauzi, S.; Stranieri, A.; Tramontano, G.; Ratti, G.; Paltrinieri, S. Evaluating the presence of domestic cat hepadnavirus viraemia in cats with biochemical alterations suggestive of liver disease. Vet. Rec. 2022, 191, e1626. [Google Scholar] [CrossRef] [PubMed]
- Alter, M.J. Epidemiology of viral hepatitis and HIV co-infection. J. Hepatol. 2006, 44, S6–S9. [Google Scholar] [CrossRef]
- Perrillo, R.P.; Chad, K.H.; Overby, L.R.; Decker, R.H. Anti-Hepatitis B Core Immunoglobulin M in the Serologic Evaluation of Hepatitis B Virus Infection and Simultaneous Infection with Type B, Delta Agent, and Non-A, Non-B Viruses. Gastroenterology 1983, 85, 163–167. [Google Scholar] [CrossRef]
- Yap, W.B.; Tey, B.T.; Ng, M.Y.; Ong, S.T.; Tan, W.S. N-terminally His-tagged hepatitis B core antigens: Construction, expression, purification and antigenicity. J. Virol. Methods 2009, 160, 125–131. [Google Scholar] [CrossRef]
- Hartmann, K. Efficacy of antiviral chemotherapy for retrovirus-infected cats: What does the current literature tell us? J. Feline Med. Surg. 2015, 17, 925–939. [Google Scholar] [CrossRef]
- Sunbul, M. Hepatitis B virus genotypes: Global distribution and clinical importance. World J. Gastroenterol. 2014, 20, 5427–5434. [Google Scholar] [CrossRef]
- Yuen, L.K.W.; Littlejohn, M.; Duchêne, S.; Edwards, R.; Bukulatjpi, S.; Binks, P.; Jackson, K.; Davies, J.; Davis, J.S.; Tong, S.Y.C.; et al. Tracing Ancient Human Migrations into Sahul Using Hepatitis B Virus Genomes. Mol. Biol. Evol. 2019, 36, 942–954. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Primer Set | Name | Primer Binding Site | Sequence | Product Size | Cycling Conditions | ||
|---|---|---|---|---|---|---|---|
| (Numbering Based on MH307930) | Denaturation | Annealing | Extension | ||||
| Pair 1 | 184F_1 | 184–203 | CCAACTTCCTGTCCTCCGAC | 776 | 95 °C, 1 min | 56 °C, 30 s | 72 °C, 1 min |
| 959R_1 | 959–940 | TCGATAAGCGGGGACAAAGG | |||||
| Pair 2 | 687F_2 | 687–706 | ACAAAAACCAAACGCTGGGG | 600 | 95 °C, 30 s | 56 °C, 30 s | 72 °C, 1 min |
| 1286R_2 | 1286–1267 | AGGGACGTAGACGAAGGACA | |||||
| Pair 3 * | 1107F_3 | 1107–1126 | CCCCTTCCTATCCATGTCGC | 802 | 95 °C, 1 min | 58 °C, 30 s | 72 °C, 1 min |
| 1908R_3 | 1908–1889 | CCGTATGGTGAGGGGAACAG | |||||
| Pair 3b * | 1107F_3 | 1107–1126 | CCCCTTCCTATCCATGTCGC | 1024 | 95 °C, 1 min | 58 °C, 30 s | 72 °C, 1 min |
| Cir5R | 2130–2113 | TGCGAATCCAGGTGCCAA | |||||
| P3Euro * | P3Euro2F | 1099–1118 | TCCATTCACCCCTTCCTATC | 790 | 95 °C, 1 min | 55 °C, 30 s | 72 °C, 1 min |
| P3Euro3R | 1888–1868 | TGTTCCCTACCTGTAAGTTCC | |||||
| Pair 4 | 1729F_4 | 1729–1748 | GCCTTAGATGGCTTTGGGGT | 768 | 95 °C, 30 s | 56 °C, 30 s | 72 °C, 1 min |
| 2496R_4 | 2496–2477 | CTTCAGACGGCGGAGTTCAT | |||||
| Pair 5 | 2110F_5 | 2110–2129 | CTTTTGGCACCTGGATTCGC | 852 | 95 °C, 30 s | 56 °C, 30 s | 72 °C, 1 min |
| 2952R_5 | 2961–2942 | TTGGTGATTGGTTGAGCGGA | |||||
| Pair 6 | 2752F_6 | 2752–2771 | GGGGTTTTTCCCTCGTCACA | 1061 | 95 °C, 30 s | 56 °C, 30 s | 72 °C, 1 min |
| Cir4R | 625–605 | AGATGTTCCACACTCTTAGCC | |||||
| Variable | Categories | PCR Positive | PCR Negative | % PCR Positive | Total | OR | 95% CI | p-Value |
|---|---|---|---|---|---|---|---|---|
| Age (months) | <6 | 3 | 17 | 15.0 | 20 | - | - | 0.246 * |
| 6–24 | 2 | 50 | 3.8 | 52 | 0.23 | 0.03–1.47 | 0.120 | |
| >24–84 | 13 | 103 | 11.2 | 116 | 0.71 | 0.18–2.78 | 0.628 | |
| >84 | 39 | 286 | 12.0 | 325 | 0.77 | 0.22–2.76 | 0.691 | |
| Sex | Female | 24 | 206 | 10.4 | 230 | - | - | - |
| Male | 33 | 250 | 11.7 | 283 | 1.13 | 0.65–1.98 | 0.661 | |
| Neutered | Yes | 48 | 379 | 11.2 | 427 | - | - | - |
| No | 8 | 56 | 12.5 | 64 | 1.13 | 0.51–2.51 | 0.768 | |
| Unknown | - | - | - | 22 | - | - | - | |
| Breed | Mixed | 26 | 216 | 10.7 | 242 | - | - | - |
| Pure | 31 | 240 | 11.4 | 271 | 1.07 | 0.62–1.86 | 0.802 | |
| Serum ALT (U/L) | Normal (12–130) | 42 | 285 | 12.8 | 327 | - | - | - |
| High (>130) | 5 | 59 | 7.8 | 64 | 0.57 | 0.22–1.51 | 0.263 | |
| Unknown | - | - | - | 122 | - | - | - |
| Sample ID | Accession | Size (nt) | L Region | S Region | X Region | C Region | Identity to Reference Sequences * | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| nt | aa | nt | aa | nt | aa | nt | aa | DCH Strain ** (Accession nr.) | nt Identity % | |||
| HK_01 | OP643851 | 3184 | 2514 | 838 | 1146 | 382 | 438 | 146 | 657 | 219 | MYS/2019/UPM CHV04 (MK902920) | 98.9 |
| HK_02 | OP643852 | 3184 | 2514 | 838 | 1146 | 382 | 438 | 146 | 654 | 218 | MYS/2019/UPM CHV04 (MK902920) | 98.9 |
| HK_03 | OP643853 | 3184 | 2514 | 838 | 1146 | 382 | 438 | 146 | 657 | 219 | MYS/2019/UPM CHV04 (MK902920) | 98.6 |
| HK_04 | OP643854 | 3184 | 2514 | 838 | 1146 | 382 | 438 | 146 | 657 | 219 | MYS/2019/UPM CHV04 (MK902920) | 99.1 |
| HK_05 | OP643855 | 3184 | 2514 | 838 | 1146 | 382 | 438 | 146 | 657 | 219 | MYS/2019/UPM CHV04 (MK902920) | 99.0 |
| HK_06 | OP643856 | 3184 | 2514 | 838 | 1146 | 382 | 438 | 146 | 657 | 219 | MYS/2019/UPM CHV04 (MK902920) | 98.8 |
| HK_07 | OP643857 | 3184 | 2514 | 838 | 1146 | 382 | 438 | 146 | 657 | 219 | MYS/2019/UPM CHV04 (MK902920) | 99.1 |
| HK_08 | OP643858 | 3184 | 2514 | 838 | 1146 | 382 | 438 | 146 | 657 | 219 | MYS/2019/UPM CHV04 (MK902920) | 99.1 |
| HK_09 | OP643859 | 3184 | 2514 | 838 | 1146 | 382 | 438 | 146 | 657 | 219 | MYS/2019/UPM CHV04 (MK902920) | 98.8 |
| HK_10 | OP643860 | 3184 | 2514 | 838 | 1146 | 382 | 438 | 146 | 657 | 219 | MYS/2019/UPM CHV04 (MK902920) | 99.1 |
| HK_11 | OP643861 | 3184 | 2514 | 838 | 1146 | 382 | 438 | 146 | 657 | 219 | MYS/2019/UPM CHV04 (MK902920) | 99.1 |
| HK_12 | OP643862 | 3184 | 2514 | 838 | 1146 | 382 | 438 | 146 | 657 | 219 | AUS/2016/Sydney (MH307930) | 98.8 |
| Genotype A | Genotype B | |||
|---|---|---|---|---|
| clade A1 | clade A2 | |||
| Genotype A | clade A1 | 100 | 95.8–97.5 | 88.7–89.2 |
| clade A2 | 95.8–97.5 | 100 | 89.0–89.5 | |
| Genotype B | 88.7–89.2 | 89.0–89.5 | 100 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capozza, P.; Carrai, M.; Choi, Y.R.; Tu, T.; Nekouei, O.; Lanave, G.; Martella, V.; Beatty, J.A.; Barrs, V.R. Domestic Cat Hepadnavirus: Molecular Epidemiology and Phylogeny in Cats in Hong Kong. Viruses 2023, 15, 150. https://0-doi-org.brum.beds.ac.uk/10.3390/v15010150
Capozza P, Carrai M, Choi YR, Tu T, Nekouei O, Lanave G, Martella V, Beatty JA, Barrs VR. Domestic Cat Hepadnavirus: Molecular Epidemiology and Phylogeny in Cats in Hong Kong. Viruses. 2023; 15(1):150. https://0-doi-org.brum.beds.ac.uk/10.3390/v15010150
Chicago/Turabian StyleCapozza, Paolo, Maura Carrai, Yan Ru Choi, Thomas Tu, Omid Nekouei, Gianvito Lanave, Vito Martella, Julia A. Beatty, and Vanessa R. Barrs. 2023. "Domestic Cat Hepadnavirus: Molecular Epidemiology and Phylogeny in Cats in Hong Kong" Viruses 15, no. 1: 150. https://0-doi-org.brum.beds.ac.uk/10.3390/v15010150