
Viral Interference with Functions of the Cellular Receptor Tyrosine Phosphatase CD45

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Viral Interference with CD45 Affecting the Activity of Immune Cells

3. Utilization of CD45 to the Benefit of Viruses
4. Further Possible Interaction of Viruses with CD45 in Myeloid Cells

5. CD45 Evolution in the Light of Recently Identified Putative Viral Ligands

6. Conclusions and Further Directions
Acknowledgments
Conflicts of Interest
References
- Williams, A.F.; Barclay, A.N. Glycoprotein antigens of the lymphocyte surface and their purification by antibody affinity chromatography. In Handbook of Experimental Immunology; Weir, D.M., Herzenberg, L.A., Eds.; Blackwell Scientific Publications: Oxford, UK, 1986; pp. 1–24. [Google Scholar]
- Desai, D.M.; Sap, J.; Silvennoinen, O.; Schlessinger, J.; Weiss, A. The catalytic activity of the CD45 membrane-proximal phosphatase domain is required for TCR signaling and regulation. EMBO J. 1994, 13, 4002–4010. [Google Scholar] [PubMed]
- Streuli, M.; Hall, L.R.; Saga, Y.; Schlossman, S.F.; Saito, H. Differential usage of three exons generates at least five different mRNAs encoding human leukocyte common antigens. J. Exp. Med. 1987, 166, 1548–1566. [Google Scholar] [CrossRef] [PubMed]
- Trowbridge, I.S.; Thomas, M.L. CD45: An emerging role as a protein tyrosine phosphatase required for lymphocyte activation and development. Annu. Rev. Immunol. 1994, 12, 85–116. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, K.; Funakoshi, Y.; Autero, M.; Horejsi, V.; Kobata, A.; Gahmberg, C.G. Structural study of the O-linked sugar chains of human leukocyte tyrosine phosphatase CD45. Eur. J. Biochem. 1998, 251, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Furukawa, K.; Autero, M.; Gahmberg, C.G.; Kobata, A. Structural study of the sugar chains of human leukocyte common antigen CD45. Biochemistry 1993, 32, 12694–12704. [Google Scholar] [CrossRef] [PubMed]
- Dianzani, U.; Redoglia, V.; Malavasi, F.; Bragardo, M.; Pileri, A.; Janeway, C.A., Jr.; Bottomly, K. Isoform-specific associations of CD45 with accessory molecules in human T lymphocytes. Eur. J. Immunol. 1992, 22, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Hermiston, M.L.; Xu, Z.; Weiss, A. CD45: A critical regulator of signaling thresholds in immune cells. Annu. Rev. Immunol. 2003, 21, 107–137. [Google Scholar] [CrossRef] [PubMed]
- Palacios, E.H.; Weiss, A. Function of the Src-family kinases, Lck and Fyn, in T-cell development and activation. Oncogene 2004, 23, 7990–8000. [Google Scholar] [CrossRef] [PubMed]
- Hermiston, M.L.; Zikherman, J.; Zhu, J.W. CD45, CD148, and Lyp/Pep: Critical phosphatases regulating Src family kinase signaling networks in immune cells. Immunol. Rev. 2009, 228, 288–311. [Google Scholar] [CrossRef] [PubMed]
- Mustelin, T.; Coggeshall, K.M.; Altman, A. Rapid activation of the T-cell tyrosine protein kinase pp56lck by the CD45 phosphotyrosine phosphatase. Proc. Natl. Acad. Sci. USA 1989, 86, 6302–6306. [Google Scholar] [CrossRef] [PubMed]
- Ostergaard, H.L.; Shackelford, D.A.; Hurley, T.R.; Johnson, P.; Hyman, R.; Sefton, B.M.; Trowbridge, I.S. Expression of CD45 alters phosphorylation of the lck-encoded tyrosine protein kinase in murine lymphoma T-cell lines. Proc. Natl. Acad. Sci. USA 1989, 86, 8959–8963. [Google Scholar] [CrossRef] [PubMed]
- Danielian, S.; Fagard, R.; Alcover, A.; Acuto, O.; Fischer, S. The lymphocyte-specific protein tyrosine kinase p56lck is hyperphosphorylated on serine and tyrosine residues within minutes after activation via T cell receptor or CD2. Eur. J. Immunol. 1989, 19, 2183–2189. [Google Scholar] [CrossRef] [PubMed]
- Rossy, J.; Williamson, D.J.; Gaus, K. How does the kinase Lck phosphorylate the T cell receptor? Spatial organization as a regulatory mechanism. Front Immunol. 2012, 3, 167. [Google Scholar] [PubMed]
- Bergman, M.; Mustelin, T.; Oetken, C.; Partanen, J.; Flint, N.A.; Amrein, K.E.; Autero, M.; Burn, P.; Alitalo, K. The human p50csk tyrosine kinase phosphorylates p56lck at Tyr-505 and down regulates its catalytic activity. EMBO J. 1992, 11, 2919–2924. [Google Scholar] [PubMed]
- Yamaguchi, H.; Hendrickson, W.A. Structural basis for activation of human lymphocyte kinase Lck upon tyrosine phosphorylation. Nature 1996, 384, 484–489. [Google Scholar] [CrossRef] [PubMed]
- D’Oro, U.; Sakaguchi, K.; Appella, E.; Ashwell, J.D. Mutational analysis of Lck in CD45-negative T cells: Dominant role of tyrosine 394 phosphorylation in kinase activity. Mol. Cell. Biol. 1996, 16, 4996–5003. [Google Scholar] [PubMed]
- McNeill, L.; Salmond, R.J.; Cooper, J.C.; Carret, C.K.; Cassady-Cain, R.L.; Roche-Molina, M.; Tandon, P.; Holmes, N.; Alexander, D.R. The differential regulation of Lck kinase phosphorylation sites by CD45 is critical for T cell receptor signaling responses. Immunity 2007, 27, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Brownlie, R.J.; Zamoyska, R. T cell receptor signalling networks: Branched, diversified and bounded. Nat. Rev. Immunol. 2013, 13, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Zikherman, J.; Jenne, C.; Watson, S.; Doan, K.; Raschke, W.; Goodnow, C.C.; Weiss, A. CD45-Csk phosphatase-kinase titration uncouples basal and inducible T cell receptor signaling during thymic development. Immunity 2010, 32, 342–354. [Google Scholar] [CrossRef] [PubMed]
- Schoenborn, J.R.; Tan, Y.X.; Zhang, C.; Shokat, K.M.; Weiss, A. Feedback circuits monitor and adjust basal Lck-dependent events in T cell receptor signaling. Sci. Signal. 2011, 4, ra59. [Google Scholar] [PubMed]
- Hui, E.; Vale, R.D. In vitro membrane reconstitution of the T-cell receptor proximal signaling network. Nat. Struct. Mol. Biol. 2014, 21, 133–142. [Google Scholar] [CrossRef] [PubMed]
- James, J.R.; Vale, R.D. Biophysical mechanism of T-cell receptor triggering in a reconstituted system. Nature 2012, 487, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Byth, K.F.; Conroy, L.A.; Howlett, S.; Smith, A.J.; May, J.; Alexander, D.R.; Holmes, N. CD45-null transgenic mice reveal a positive regulatory role for CD45 in early thymocyte development, in the selection of CD4+CD8+ thymocytes, and B cell maturation. J. Exp. Med. 1996, 183, 1707–1718. [Google Scholar] [CrossRef] [PubMed]
- Kishihara, K.; Penninger, J.; Wallace, V.A.; Kundig, T.M.; Kawai, K.; Wakeham, A.; Timms, E.; Pfeffer, K.; Ohashi, P.S.; Thomas, M.L.; et al. Normal B lymphocyte development but impaired T cell maturation in CD45-exon6 protein tyrosine phosphatase-deficient mice. Cell 1993, 74, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Mee, P.J.; Turner, M.; Basson, M.A.; Costello, P.S.; Zamoyska, R.; Tybulewicz, V.L. Greatly reduced efficiency of both positive and negative selection of thymocytes in CD45 tyrosine phosphatase-deficient mice. Eur. J. Immunol. 1999, 29, 2923–2933. [Google Scholar] [CrossRef] [PubMed]
- Cale, C.M.; Klein, N.J.; Novelli, V.; Veys, P.; Jones, A.M.; Morgan, G. Severe combined immunodeficiency with abnormalities in expression of the common leucocyte antigen, CD45. Arch. Dis. Child 1997, 76, 163–164. [Google Scholar] [CrossRef] [PubMed]
- Kung, C.; Pingel, J.T.; Heikinheimo, M.; Klemola, T.; Varkila, K.; Yoo, L.I.; Vuopala, K.; Poyhonen, M.; Uhari, M.; Rogers, M.; et al. Mutations in the tyrosine phosphatase CD45 gene in a child with severe combined immunodeficiency disease. Nat. Med. 2000, 6, 343–345. [Google Scholar] [CrossRef] [PubMed]
- Tchilian, E.Z.; Wallace, D.L.; Wells, R.S.; Flower, D.R.; Morgan, G.; Beverley, P.C. A deletion in the gene encoding the CD45 antigen in a patient with SCID. J. Immunol. 2001, 166, 1308–1313. [Google Scholar] [CrossRef] [PubMed]
- Yamada, H.; Kishihara, K.; Kong, Y.Y.; Nomoto, K. Enhanced generation of NK cells with intact cytotoxic function in CD45 exon 6-deficient mice. J. Immunol. 1996, 157, 1523–1528. [Google Scholar] [PubMed]
- Huntington, N.D.; Xu, Y.; Nutt, S.L.; Tarlinton, D.M. A requirement for CD45 distinguishes Ly49D-mediated cytokine and chemokine production from killing in primary natural killer cells. J. Exp. Med. 2005, 201, 1421–1433. [Google Scholar] [CrossRef] [PubMed]
- Mason, L.H.; Willette-Brown, J.; Taylor, L.S.; McVicar, D.W. Regulation of Ly49D/DAP12 signal transduction by Src-family kinases and CD45. J. Immunol. 2006, 176, 6615–6623. [Google Scholar] [CrossRef] [PubMed]
- Hesslein, D.G.; Takaki, R.; Hermiston, M.L.; Weiss, A.; Lanier, L.L. Dysregulation of signaling pathways in CD45-deficient NK cells leads to differentially regulated cytotoxicity and cytokine production. Proc. Natl. Acad. Sci. USA 2006, 103, 7012–7017. [Google Scholar] [CrossRef] [PubMed]
- Hesslein, D.G.; Palacios, E.H.; Sun, J.C.; Beilke, J.N.; Watson, S.R.; Weiss, A.; Lanier, L.L. Differential requirements for CD45 in NK-cell function reveal distinct roles for Syk-family kinases. Blood 2011, 117, 3087–3095. [Google Scholar] [CrossRef] [PubMed]
- Caignard, G.; Leiva-Torres, G.A.; Leney-Greene, M.; Charbonneau, B.; Dumaine, A.; Fodil-Cornu, N.; Pyzik, M.; Cingolani, P.; Schwartzentruber, J.; Dupaul-Chicoine, J.; et al. Genome-wide mouse mutagenesis reveals CD45-mediated T cell function as critical in protective immunity to HSV-1. PLoS. Pathog. 2013, 9, e1003637. [Google Scholar] [CrossRef] [PubMed]
- Panchal, R.G.; Bradfute, S.B.; Peyser, B.D.; Warfield, K.L.; Ruthel, G.; Lane, D.; Kenny, T.A.; Anderson, A.O.; Raschke, W.C.; Bavari, S. Reduced levels of protein tyrosine phosphatase CD45 protect mice from the lethal effects of Ebola virus infection. Cell Host. Microbe 2009, 6, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Panchal, R.G.; Ulrich, R.L.; Bradfute, S.B.; Lane, D.; Ruthel, G.; Kenny, T.A.; Iversen, P.L.; Anderson, A.O.; Gussio, R.; Raschke, W.C.; et al. Reduced expression of CD45 protein-tyrosine phosphatase provides protection against anthrax pathogenesis. J. Biol. Chem. 2009, 284, 12874–12885. [Google Scholar] [CrossRef] [PubMed]
- Van Vliet, S.J.; Gringhuis, S.I.; Geijtenbeek, T.B.; van Kooyk, Y. Regulation of effector T cells by antigen-presenting cells via interaction of the C-type lectin MGL with CD45. Nat. Immunol. 2006, 7, 1200–1208. [Google Scholar] [CrossRef] [PubMed]
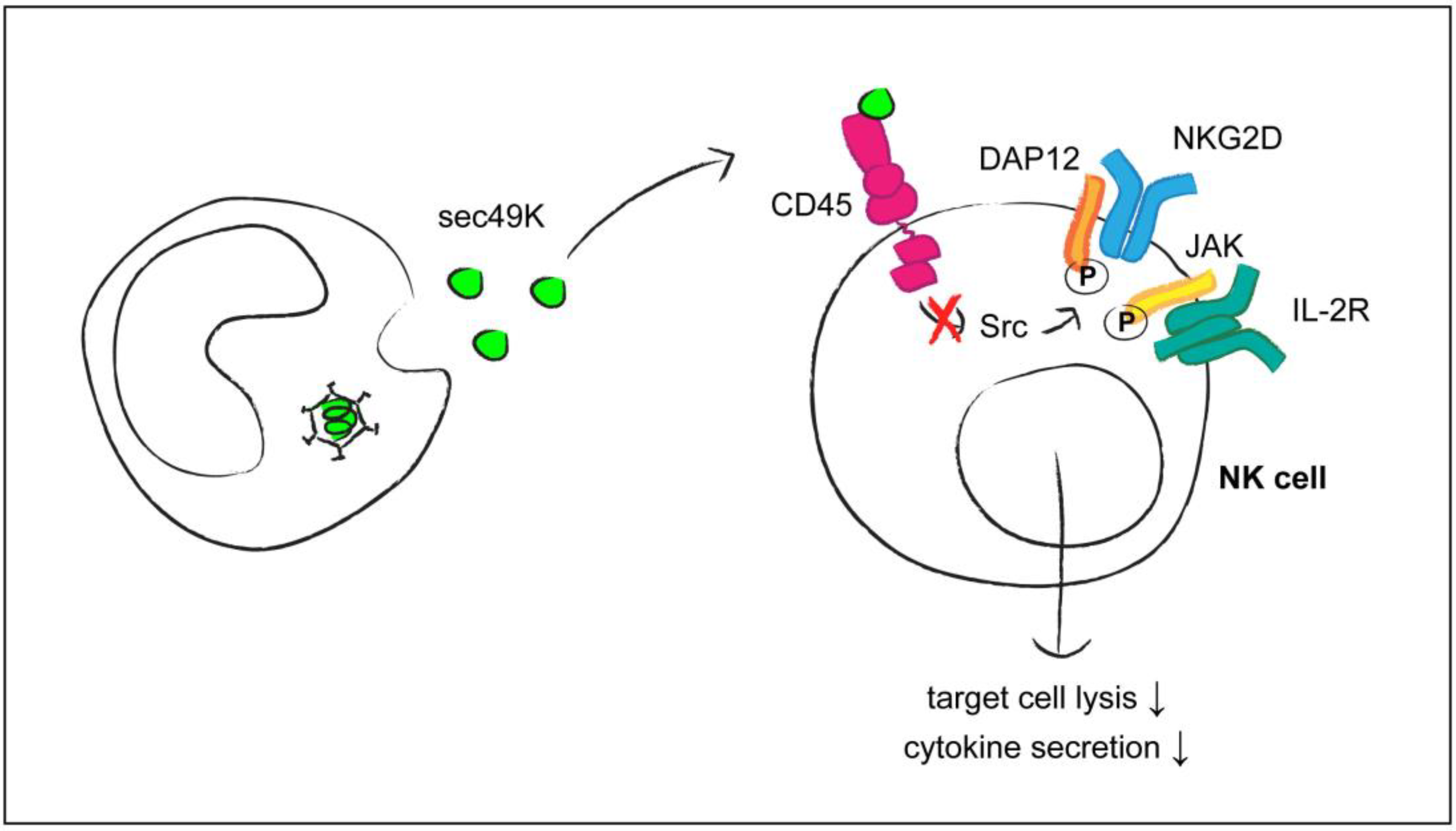
- Gabaev, I.; Steinbrück, L.; Pokoyski, C.; Pich, A.; Stanton, R.J.; Schwinzer, R.; Schulz, T.F.; Jacobs, R.; Messerle, M.; Kay-Fedorov, P.C. The human cytomegalovirus UL11 protein interacts with the receptor tyrosine phosphatase CD45, resulting in functional paralysis of T cells. PLoS. Pathog. 2011, 7, e1002432. [Google Scholar] [CrossRef] [PubMed]
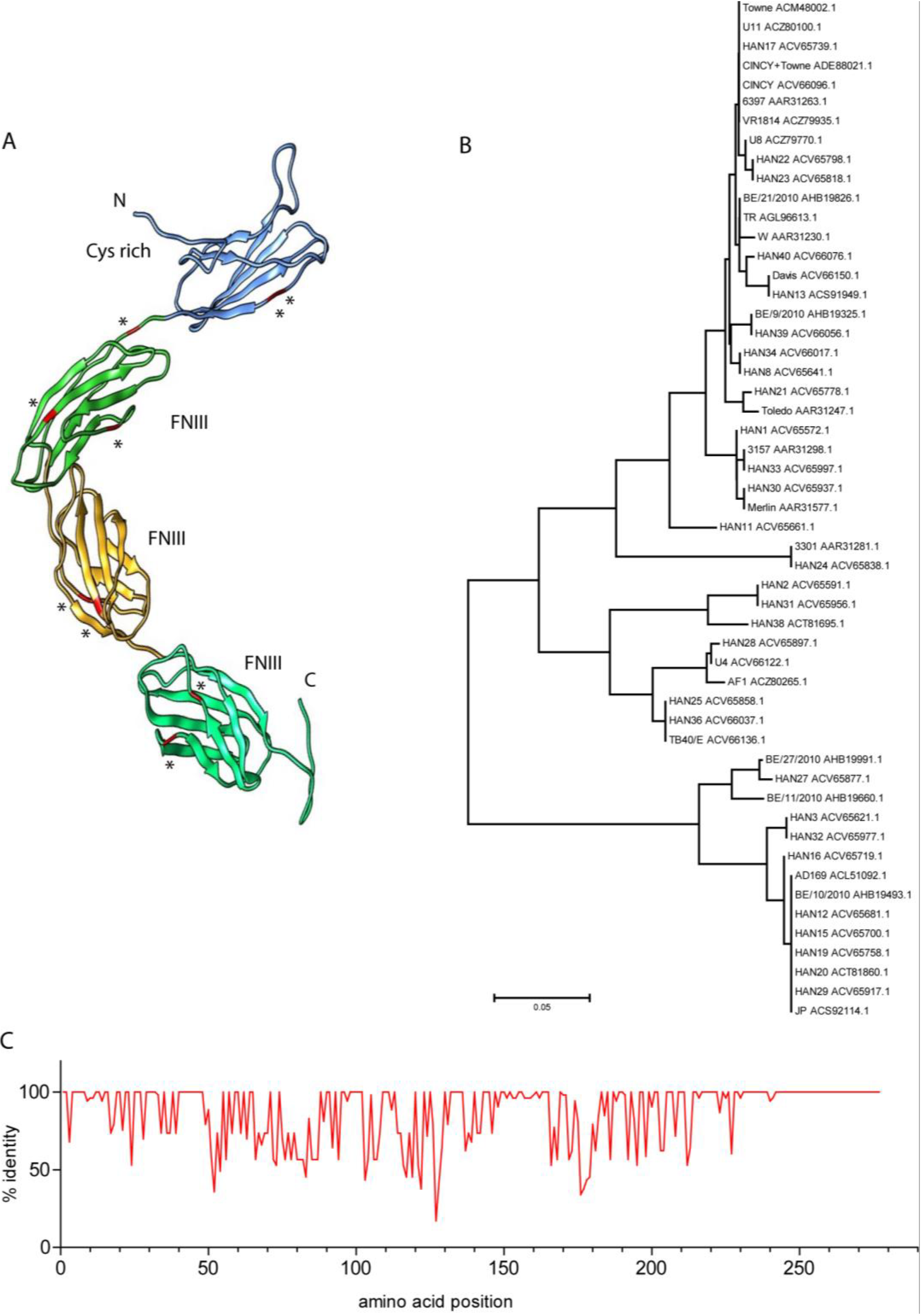
- Windheim, M.; Southcombe, J.H.; Kremmer, E.; Chaplin, L.; Urlaub, D.; Falk, C.S.; Claus, M.; Mihm, J.; Braithwaite, M.; Dennehy, K.; et al. A unique secreted adenovirus E3 protein binds to the leukocyte common antigen CD45 and modulates leukocyte functions. Proc. Natl. Acad. Sci. USA 2013, 110, e4884–e4893. [Google Scholar] [CrossRef] [PubMed]
- Davison, A.J.; Akter, P.; Cunningham, C.; Dolan, A.; Addison, C.; Dargan, D.J.; Hassan-Walker, A.F.; Emery, V.C.; Griffiths, P.D.; Wilkinson, G.W. Homology between the human cytomegalovirus RL11 gene family and human adenovirus E3 genes. J. Gen. Virol. 2003, 84, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Dolan, A.; Cunningham, C.; Hector, R.D.; Hassan-Walker, A.F.; Lee, L.; Addison, C.; Dargan, D.J.; McGeoch, D.J.; Gatherer, D.; Emery, V.C.; et al. Genetic content of wild-type human cytomegalovirus. J. Gen. Virol. 2004, 85, 1301–1312. [Google Scholar] [CrossRef] [PubMed]
- Sekulin, K.; Görzer, I.; Heiss-Czedik, D.; Puchhammer-Stöckl, E. Analysis of the variability of CMV strains in the RL11D domain of the RL11 multigene family. Virus Genes 2007, 35, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Hitomi, S.; Kozuka-Hata, H.; Chen, Z.; Sugano, S.; Yamaguchi, N.; Watanabe, S. Human cytomegalovirus open reading frame UL11 encodes a highly polymorphic protein expressed on the infected cell surface. Arch. Virol. 1997, 142, 1407–1427. [Google Scholar] [CrossRef] [PubMed]
- Gabaev, I.; Elbasani, E.; Ameres, S.; Steinbrück, L.; Stanton, R.; Döring, M.; Lenac-Rovis, T.; Kalinke, U.; Jonjic, S.; Moosmann, A.; et al. Expression of the Human Cytomegalovirus UL11 Glycoprotein in Viral Infection and Evaluation of Its Effect on Virus-Specific CD8 T Cells. J. Virol. 2014, 88, 14326–14339. [Google Scholar] [CrossRef] [PubMed]
- Jahn, G.; Stenglein, S.; Riegler, S.; Einsele, H.; Sinzger, C. Human cytomegalovirus infection of immature dendritic cells and macrophages. Intervirology 1999, 42, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Deryckere, F.; Burgert, H.G. Early region 3 of adenovirus type 19 (subgroup D) encodes an HLA-binding protein distinct from that of subgroups B and C. J. Virol. 1996, 70, 2832–2841. [Google Scholar] [PubMed]
- Anand, A.R.; Ganju, R.K. HIV-1 gp120-mediated apoptosis of T cells is regulated by the membrane tyrosine phosphatase CD45. J. Biol. Chem. 2006, 281, 12289–12299. [Google Scholar] [CrossRef] [PubMed]
- Okoye, A.A.; Picker, L.J. CD4(+) T-cell depletion in HIV infection: Mechanisms of immunological failure. Immunol. Rev. 2013, 254, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Dupere-Minier, G.; Desharnais, P.; Bernier, J. Involvement of tyrosine phosphatase CD45 in apoptosis. Apoptosis. 2010, 15, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Earl, L.A.; Baum, L.G. CD45 glycosylation controls T-cell life and death. Immunol. Cell Biol. 2008, 86, 608–615. [Google Scholar] [CrossRef] [PubMed]
- Grossman, Z.; Meier-Schellersheim, M.; Sousa, A.E.; Victorino, R.M.; Paul, W.E. CD4+ T-cell depletion in HIV infection: Are we closer to understanding the cause? Nat. Med. 2002, 8, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Tchilian, E.Z.; Beverley, P.C. Altered CD45 expression and disease. Trends Immunol. 2006, 27, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Tchilian, E.Z.; Wallace, D.L.; Dawes, R.; Imami, N.; Burton, C.; Gotch, F.; Beverley, P.C. A point mutation in CD45 may be associated with an increased risk of HIV-1 infection. AIDS 2001, 15, 1892–1894. [Google Scholar] [CrossRef] [PubMed]
- Stanton, T.; Boxall, S.; Bennett, A.; Kaleebu, P.; Watera, C.; Whitworth, J.; French, N.; Dawes, R.; Hill, A.V.; Bodmer, W.; et al. CD45 variant alleles: Possibly increased frequency of a novel exon 4 CD45 polymorphism in HIV seropositive Ugandans. Immunogenetics 2004, 56, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Dawes, R.; Hennig, B.; Irving, W.; Petrova, S.; Boxall, S.; Ward, V.; Wallace, D.; Macallan, D.C.; Thursz, M.; Hill, A.; et al. Altered CD45 expression in C77G carriers influences immune function and outcome of hepatitis C infection. J. Med. Genet. 2006, 43, 678–684. [Google Scholar] [CrossRef] [PubMed]
- Boxall, S.; Stanton, T.; Hirai, K.; Ward, V.; Yasui, T.; Tahara, H.; Tamori, A.; Nishiguchi, S.; Shiomi, S.; Ishiko, O.; et al. Disease associations and altered immune function in CD45 138G variant carriers. Hum. Mol. Genet. 2004, 13, 2377–2384. [Google Scholar] [CrossRef] [PubMed]
- Barcellos, L.F.; Caillier, S.; Dragone, L.; Elder, M.; Vittinghoff, E.; Bucher, P.; Lincoln, R.R.; Pericak-Vance, M.; Haines, J.L.; Weiss, A.; et al. PTPRC (CD45) is not associated with the development of multiple sclerosis in U.S. patients. Nat. Genet. 2001, 29, 23–24. [Google Scholar] [CrossRef] [PubMed]
- Mazurov, D.; Ilinskaya, A.; Heidecker, G.; Filatov, A. Role of O-glycosylation and expression of CD43 and CD45 on the surfaces of effector T cells in human T cell leukemia virus type 1 cell-to-cell infection. J. Virol. 2012, 86, 2447–2458. [Google Scholar] [CrossRef] [PubMed]
- Graf, B.; Bushnell, T.; Miller, J. LFA-1-mediated T cell costimulation through increased localization of TCR/class II complexes to the central supramolecular activation cluster and exclusion of CD45 from the immunological synapse. J. Immunol. 2007, 179, 1616–1624. [Google Scholar] [CrossRef] [PubMed]
- Dustin, M.L. Signaling at neuro/immune synapses. J. Clin. Investig. 2012, 122, 1149–1155. [Google Scholar] [CrossRef] [PubMed]
- Jolly, C. T cell polarization at the virological synapse. Viruses 2010, 2, 1261–1278. [Google Scholar] [CrossRef] [PubMed]
- Igakura, T.; Stinchcombe, J.C.; Goon, P.K.; Taylor, G.P.; Weber, J.N.; Griffiths, G.M.; Tanaka, Y.; Osame, M.; Bangham, C.R. Spread of HTLV-I between lymphocytes by virus-induced polarization of the cytoskeleton. Science 2003, 299, 1713–1716. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Cox, J.; Wang, P.; Krishnan, M.N.; Dai, J.; Qian, F.; Anderson, J.F.; Fikrig, E. A C-type lectin collaborates with a CD45 phosphatase homolog to facilitate West Nile virus infection of mosquitoes. Cell 2010, 142, 714–725. [Google Scholar] [CrossRef] [PubMed]
- Daley-Bauer, L.P.; Roback, L.J.; Wynn, G.M.; Mocarski, E.S. Cytomegalovirus hijacks CX3CR1(hi) patrolling monocytes as immune-privileged vehicles for dissemination in mice. Cell Host. Microbe 2014, 15, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, J.; Reeves, M. The intimate relationship between human cytomegalovirus and the dendritic cell lineage. Front Microbiol. 2014, 5, 389. [Google Scholar] [PubMed]
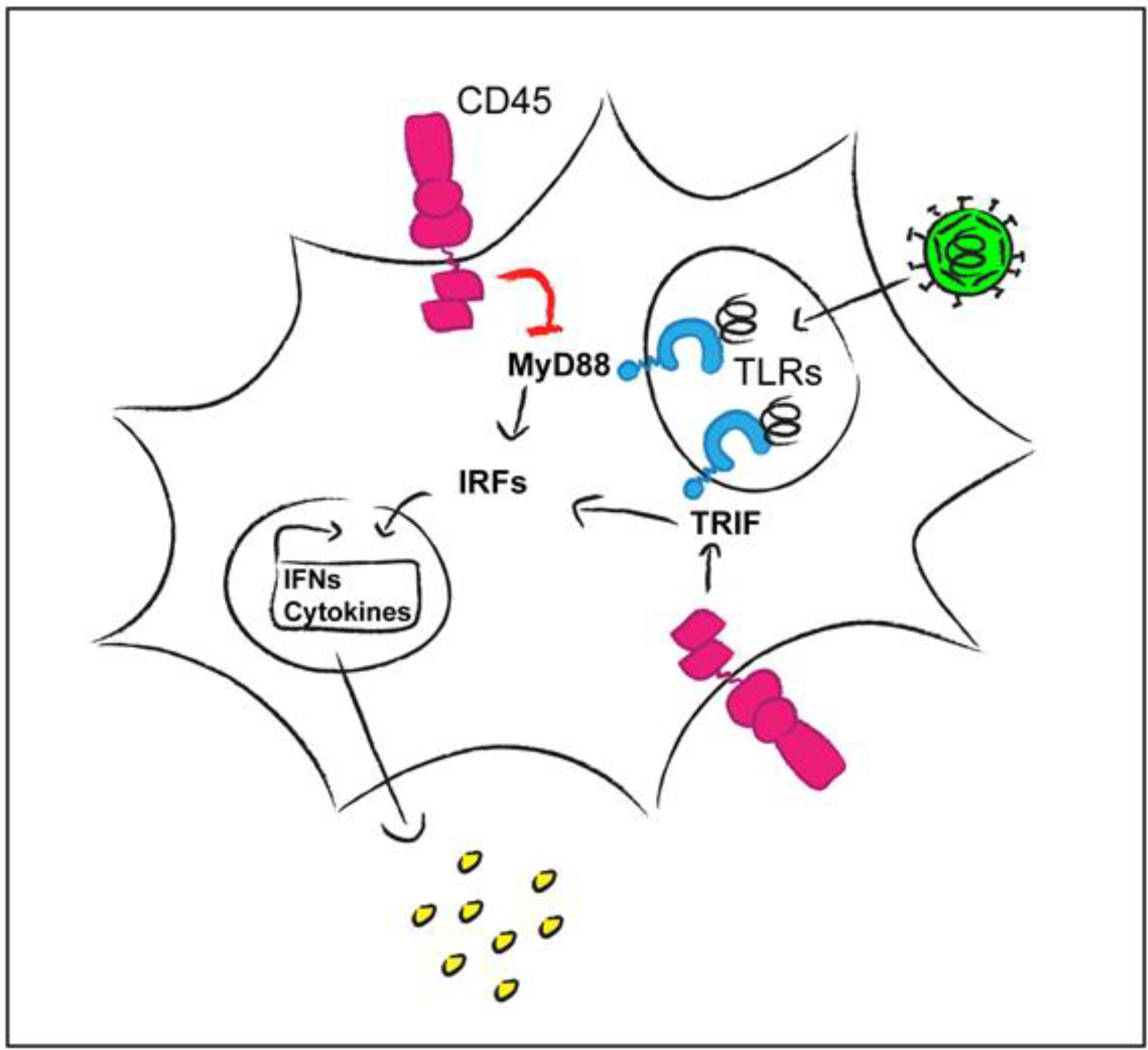
- Montoya, M.; Dawes, R.; Reid, D.; Lee, L.N.; Piercy, J.; Borrow, P.; Tchilian, E.Z.; Beverley, P.C. CD45 is required for type I IFN production by dendritic cells. Eur. J. Immunol. 2006, 36, 2150–2158. [Google Scholar] [CrossRef] [PubMed]
- Cross, J.L.; Kott, K.; Miletic, T.; Johnson, P. CD45 regulates TLR-induced proinflammatory cytokine and IFN-beta secretion in dendritic cells. J. Immunol. 2008, 180, 8020–8029. [Google Scholar] [CrossRef] [PubMed]
- Piercy, J.; Petrova, S.; Tchilian, E.Z.; Beverley, P.C. CD45 negatively regulates tumour necrosis factor and interleukin-6 production in dendritic cells. Immunology 2006, 118, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Page, T.H.; Smolinska, M.; Gillespie, J.; Urbaniak, A.M.; Foxwell, B.M. Tyrosine kinases and inflammatory signalling. Curr. Mol. Med. 2009, 9, 69–85. [Google Scholar] [CrossRef] [PubMed]
- Irie-Sasaki, J.; Sasaki, T.; Matsumoto, W.; Opavsky, A.; Cheng, M.; Welstead, G.; Griffiths, E.; Krawczyk, C.; Richardson, C.D.; Aitken, K.; et al. CD45 is a JAK phosphatase and negatively regulates cytokine receptor signalling. Nature 2001, 409, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Bijian, K.; Shen, S.H. CD45 recruits adapter protein DOK-1 and negatively regulates JAK-STAT signaling in hematopoietic cells. Mol. Immunol. 2009, 46, 2167–2177. [Google Scholar] [CrossRef] [PubMed]
- Blay, R.; Simpson, K.; Leslie, K.; Huber, S. Coxsackievirus-induced disease. CD4+ cells initiate both myocarditis and pancreatitis in DBA/2 mice. Am. J. Pathol. 1989, 135, 899–907. [Google Scholar] [PubMed]
- Roach, T.; Slater, S.; Koval, M.; White, L.; Cahir McFarland, E.D.; Okumura, M.; Thomas, M.; Brown, E. CD45 regulates Src family member kinase activity associated with macrophage integrin-mediated adhesion. Curr. Biol. 1997, 7, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Shenoi, H.; Seavitt, J.; Zheleznyak, A.; Thomas, M.L.; Brown, E.J. Regulation of integrin-mediated T cell adhesion by the transmembrane protein tyrosine phosphatase CD45. J. Immunol. 1999, 162, 7120–7127. [Google Scholar] [PubMed]
- Li, R.; Wong, N.; Jabali, M.D.; Johnson, P. CD44-initiated cell spreading induces Pyk2 phosphorylation, is mediated by Src family kinases, and is negatively regulated by CD45. J. Biol. Chem. 2001, 276, 28767–28773. [Google Scholar] [CrossRef] [PubMed]
- Harburger, D.S.; Calderwood, D.A. Integrin signalling at a glance. J. Cell Sci. 2009, 122, 159–163. [Google Scholar] [CrossRef] [PubMed]
- St Pierre, J.; Ostergaard, H.L. A role for the protein tyrosine phosphatase CD45 in macrophage adhesion through the regulation of paxillin degradation. PLoS ONE 2013, 8, e71531. [Google Scholar] [CrossRef] [PubMed]
- Ballingall, K.T.; Waibochi, L.; Holmes, E.C.; Woelk, C.H.; MacHugh, N.D.; Lutje, V.; McKeever, D.J. The CD45 locus in cattle: Allelic polymorphism and evidence for exceptional positive natural selection. Immunogenetics 2001, 52, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Filip, L.C.; Mundy, N.I. Rapid evolution by positive Darwinian selection in the extracellular domain of the abundant lymphocyte protein CD45 in primates. Mol. Biol. Evol. 2004, 21, 1504–1511. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, M.; Guex, N.; Patin, E.; Martin, O.; Xenarios, I.; Ciuffi, A.; Quintana-Murci, L.; Telenti, A. Evolutionary trajectories of primate genes involved in HIV pathogenesis. Mol. Biol. Evol. 2009, 26, 2865–2875. [Google Scholar] [CrossRef] [PubMed]
- Davison, A.J.; Dolan, A.; Akter, P.; Addison, C.; Dargan, D.J.; Alcendor, D.J.; McGeoch, D.J.; Hayward, G.S. The human cytomegalovirus genome revisited: Comparison with the chimpanzee cytomegalovirus genome. J. Gen. Virol. 2003, 84, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Pal, C.; Papp, B.; Lercher, M.J. An integrated view of protein evolution. Nat. Rev. Genet. 2006, 7, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Källberg, M.; Wang, H.; Wang, S.; Peng, J.; Wang, Z.; Lu, H.; Xu, J. Template-based protein structure modeling using the RaptorX web server. Nat. Protoc. 2012, 7, 1511–1522. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Wang, S.; Zhao, F.; Xu, J. Protein threading using context-specific alignment potential. Bioinformatics. 2013, 29, i257–i265. [Google Scholar] [CrossRef] [PubMed]
- Meng, E.C.; Pettersen, E.F.; Couch, G.S.; Huang, C.C.; Ferrin, T.E. Tools for integrated sequence-structure analysis with UCSF Chimera. BMC Bioinformatics. 2006, 7, 339. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.; Clamp, M.; Barton, G.J. Jalview Version 2-a multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed]
- Blusch, J.H.; Deryckere, F.; Windheim, M.; Ruzsics, Z.; Arnberg, N.; Adrian, T.; Burgert, H.G. The novel early region 3 protein E3/49K is specifically expressed by adenoviruses of subgenus D: Implications for epidemic keratoconjunctivitis and adenovirus evolution. Virology 2002, 296, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Robinson, C.M.; Singh, G.; Lee, J.Y.; Dehghan, S.; Rajaiya, J.; Liu, E.B.; Yousuf, M.A.; Betensky, R.A.; Jones, M.S.; Dyer, D.W.; et al. Molecular evolution of human adenoviruses. Sci. Rep. 2013, 3, 1812. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Robinson, C.M.; Dehghan, S.; Jones, M.S.; Dyer, D.W.; Seto, D.; Chodosh, J. Homologous recombination in E3 genes of human adenovirus species D. J. Virol. 2013, 87, 12481–12488. [Google Scholar] [CrossRef] [PubMed]
- Arnberg, N. Adenovirus E3 protein modulates leukocyte functions. Proc. Natl. Acad. Sci. USA 2013, 110, 19976–19977. [Google Scholar] [CrossRef] [PubMed]
- Symons, A.; Cooper, D.N.; Barclay, A.N. Characterization of the interaction between galectin-1 and lymphocyte glycoproteins CD45 and Thy-1. Glycobiology 2000, 10, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Luke, P.P.; Yang, H.; Visser, L.; Sun, H.; Garcia, B.; Qian, H.; Xiang, Y.; Huang, X.; Liu, W.; et al. Anti-CD45RB monoclonal antibody prolongs renal allograft survival in cynomolgus monkeys. Am. J. Transplant. 2007, 7, 27–37. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thiel, N.; Zischke, J.; Elbasani, E.; Kay-Fedorov, P.; Messerle, M. Viral Interference with Functions of the Cellular Receptor Tyrosine Phosphatase CD45. Viruses 2015, 7, 1540-1557. https://0-doi-org.brum.beds.ac.uk/10.3390/v7031540
Thiel N, Zischke J, Elbasani E, Kay-Fedorov P, Messerle M. Viral Interference with Functions of the Cellular Receptor Tyrosine Phosphatase CD45. Viruses. 2015; 7(3):1540-1557. https://0-doi-org.brum.beds.ac.uk/10.3390/v7031540
Chicago/Turabian StyleThiel, Nadine, Jasmin Zischke, Endrit Elbasani, Penelope Kay-Fedorov, and Martin Messerle. 2015. "Viral Interference with Functions of the Cellular Receptor Tyrosine Phosphatase CD45" Viruses 7, no. 3: 1540-1557. https://0-doi-org.brum.beds.ac.uk/10.3390/v7031540