
Interaction of Supramolecular Congo Red and Congo Red-Doxorubicin Complexes with Proteins for Drug Carrier Design

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
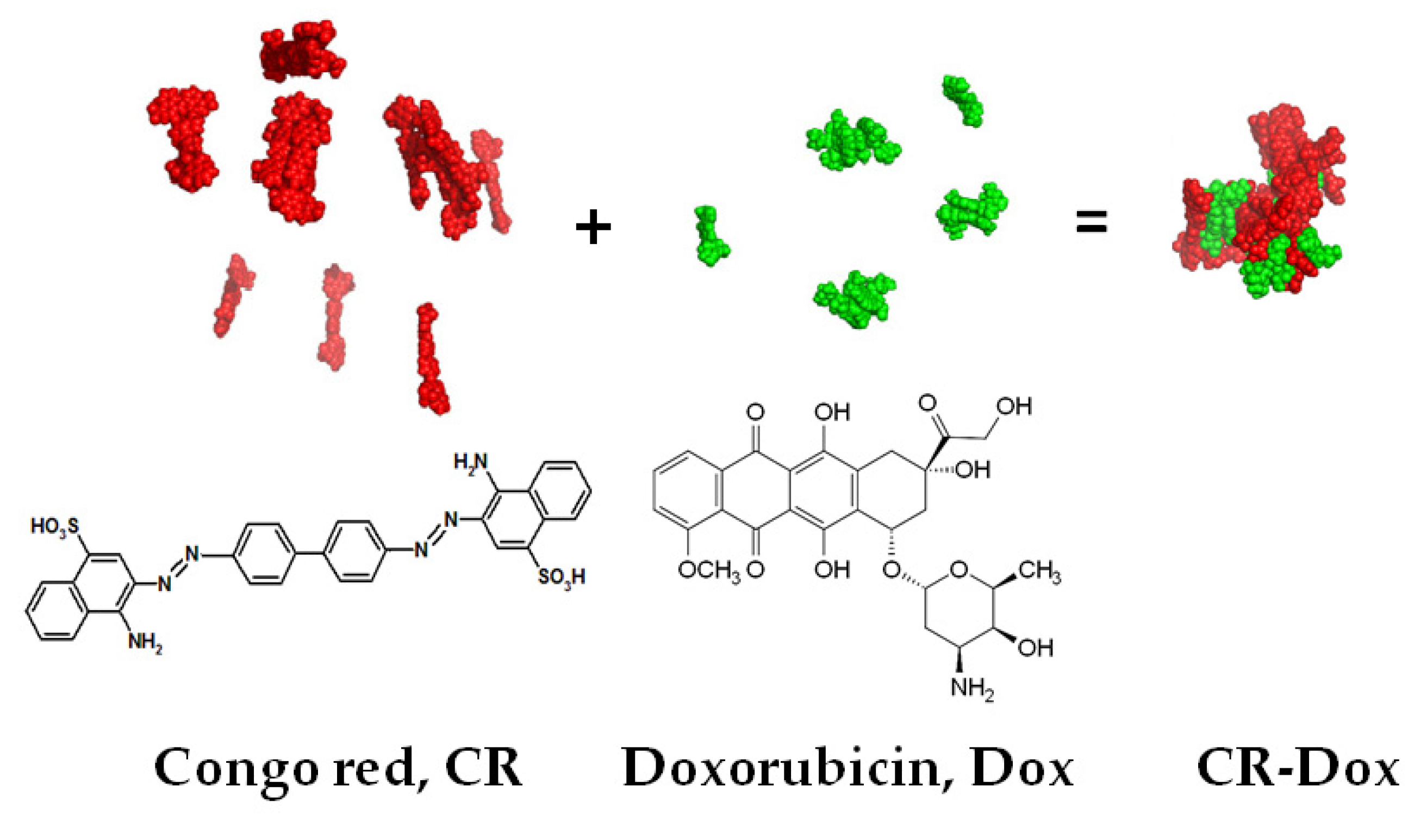
2.2. Methods of CR-Dox Preparation
2.3. Methods of CR and CR-Dox Binding with Protein
Complexes of CR and CR-Dox with Proteins
- The immunoglobulin light chain λ (Lλ)
- 2.
- Heat aggregated immunoglobulin G (HAI)
- 3.
- Albumin
2.4. Characterization of Free CR-Dox Complexes or CR-Dox Bound to Albumin, Lλ Light Chain, or HAI
2.4.1. Dynamic Light Scattering (DLS)
2.4.2. Gel-Filtration Chromatography (BioGel P-10 and BioGel P-300)
2.4.3. Gel Electrophoresis
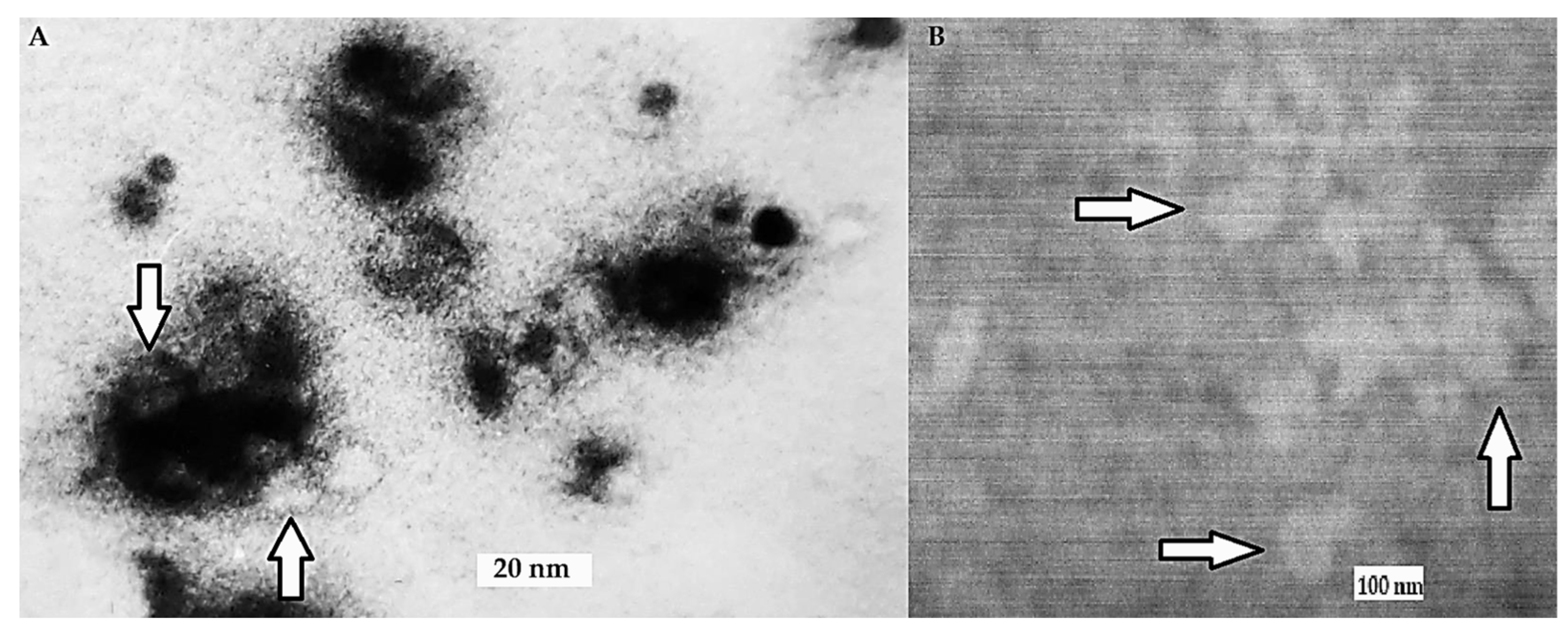
2.4.4. TEM
2.5. Methods of Analysis of Competition between BSA and HAI for Binding of the CR-RhoB Complexes (Congo Red-Rhodamine B) or CR-Dox (Congo Red-Doxorubicin) Complexes
3. Results
3.1. Characterization of CR-Dox Co-Micelles
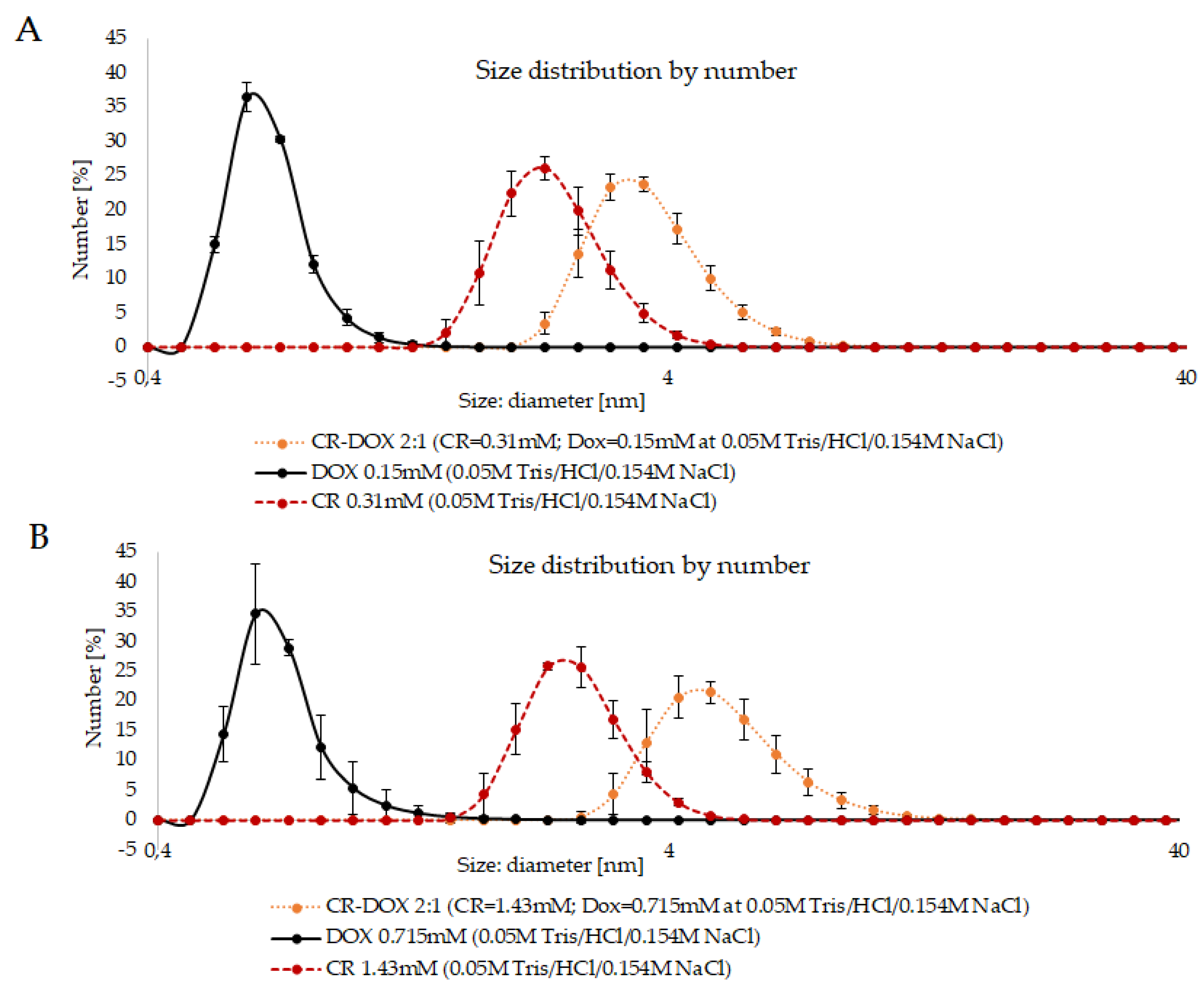
3.1.1. DLS Analysis of CR-Dox Complexes—The Effect of Concentration on the Complex Size
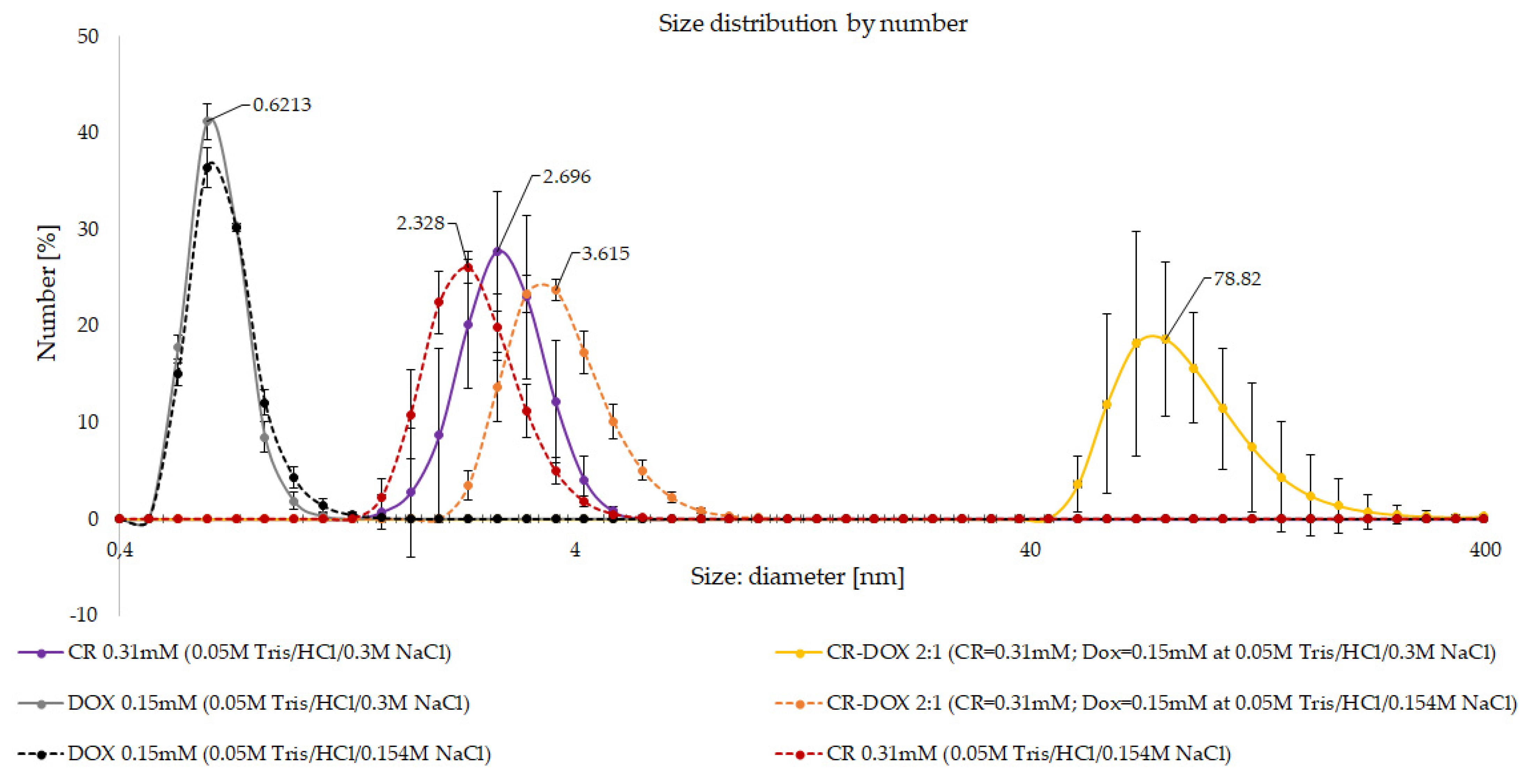
3.1.2. DLS Analysis of CR-Dox Complexes—The Effect of Ionic Strength on Complex Size
3.1.3. Gel-Filtration Chromatography (BioGel P-10): Elution Volume (Ve) of CR, Dox, and CR-Dox
3.2. Interaction of Free CR and CR-Dox Co-Micelles with Proteins
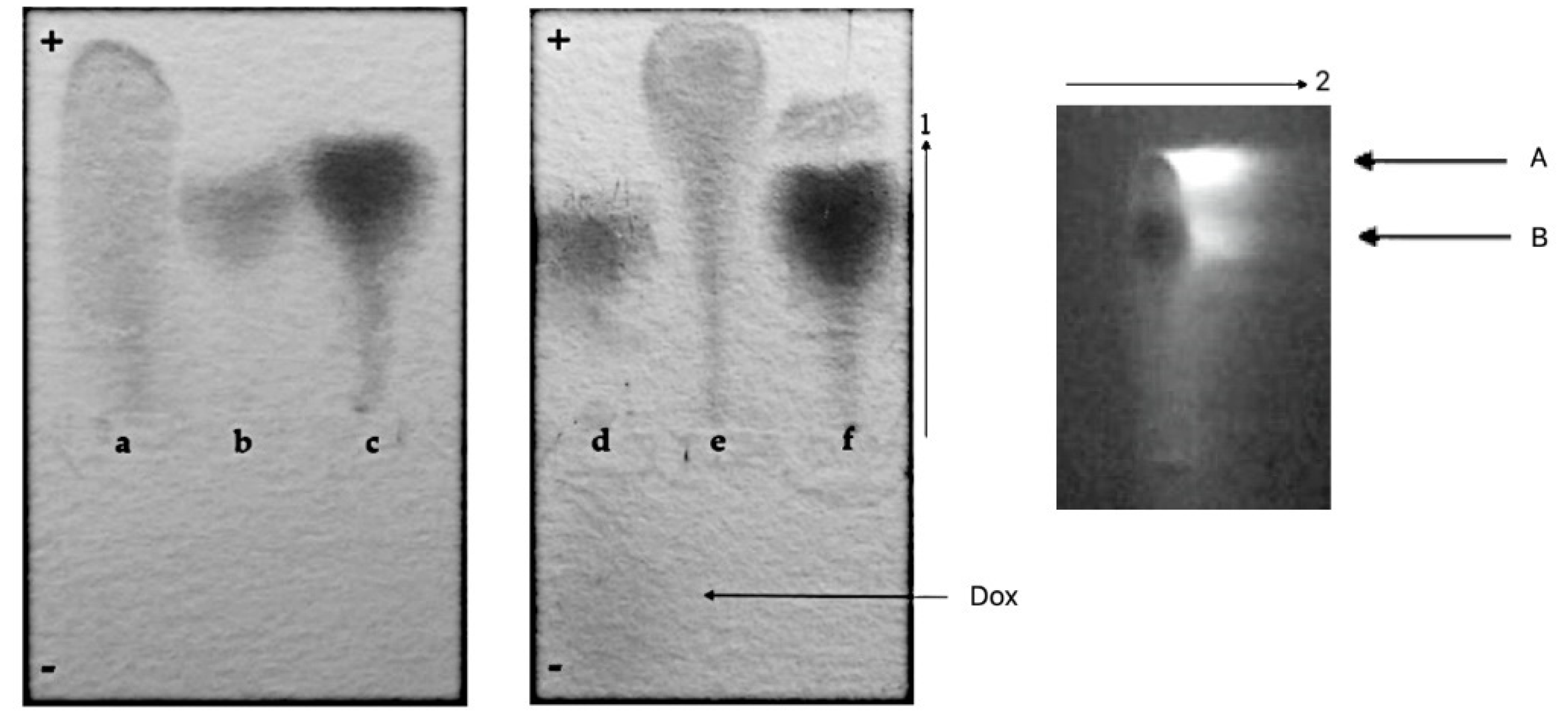
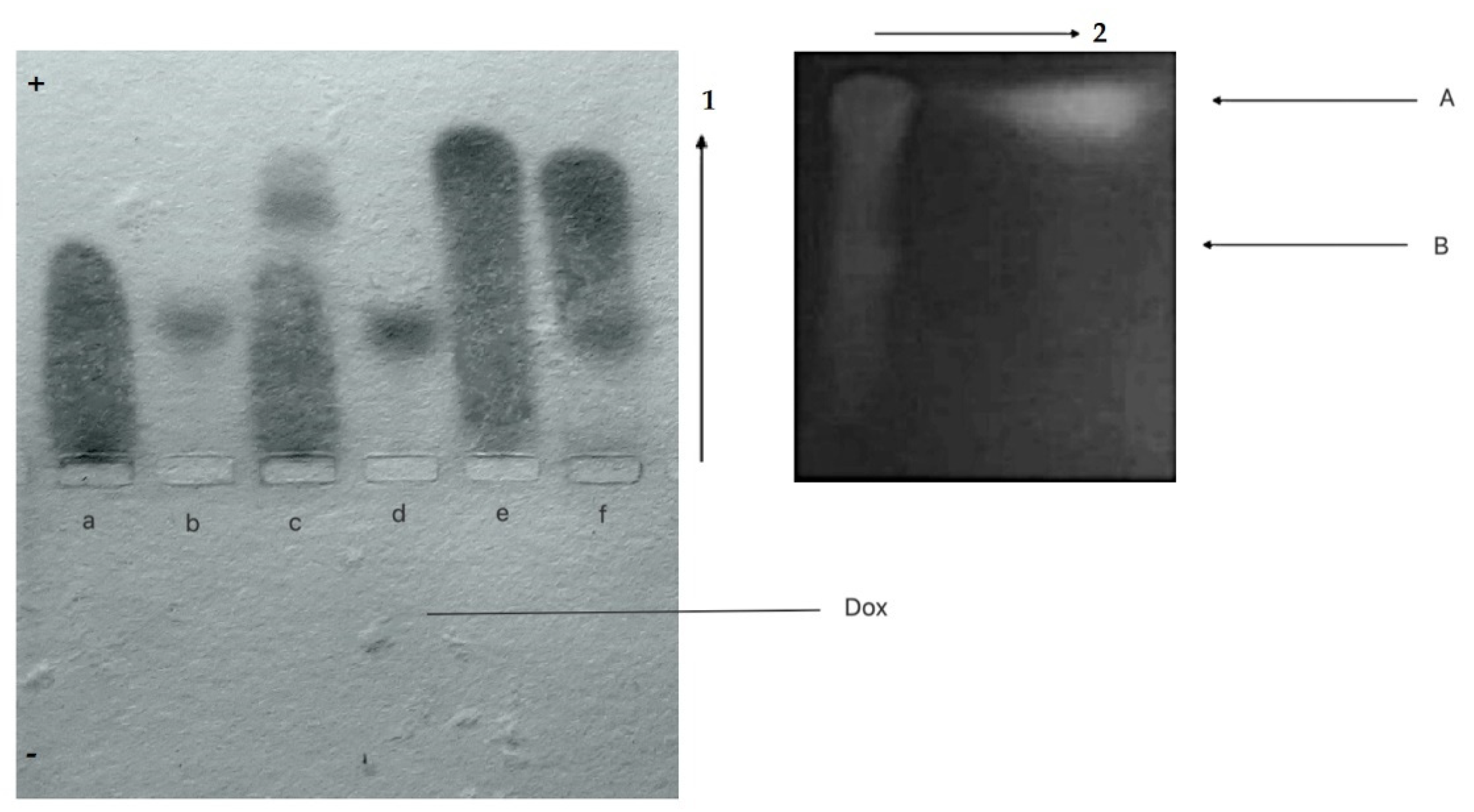
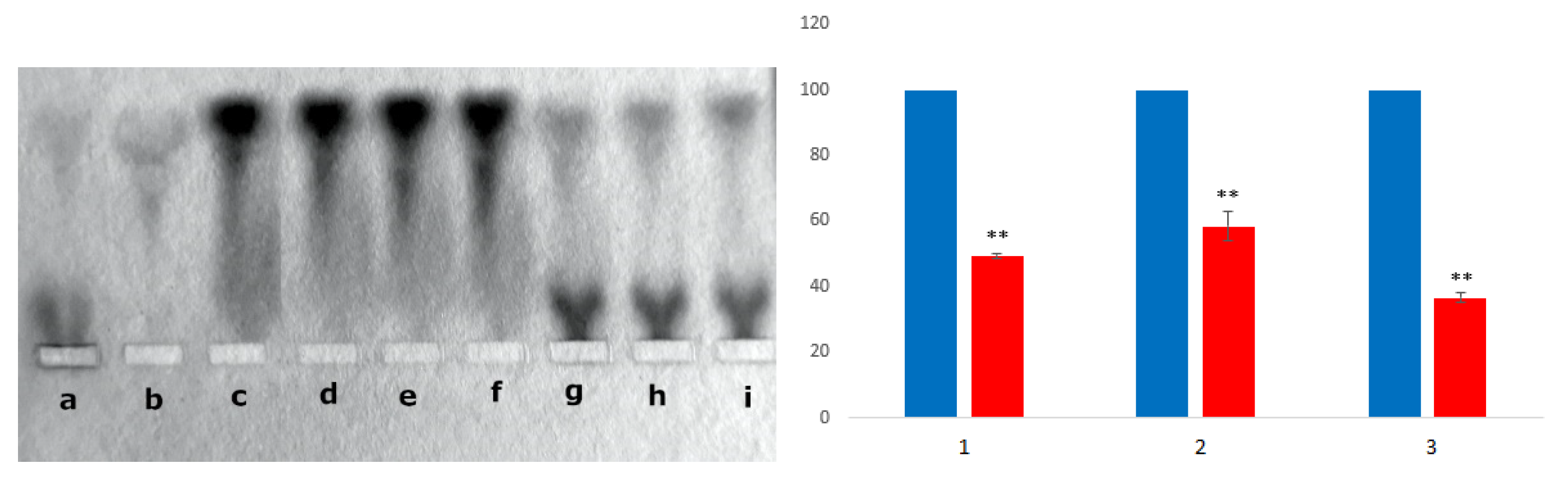
3.2.1. Agarose Gel Electrophoresis and Chromatographic Analysis
- CR-Dox co-micelles form complexes with BSA (BSA-CR-Dox)
- 2.
- CR-Dox co-micelles do not form complexes with Lλ
3.2.2. Gel-Filtration Chromatography (BioGel P-300): Elution Volumes of CR and CR-Dox Complexes with Proteins
3.2.3. CR Binds to Heat Aggregated Immunoglobulins G
3.2.4. Competition of BSA and HAI for Binding of the CR-RhoB Complexes (Congo Red-Rhodamine B) or CR-Dox (Congo Red-Doxorubicin) Complexes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Riley, R.S.; June, C.H.; Langer, R.; Mitchell, M.J. Delivery Technologies for Cancer Immunotherapy. Nat. Rev. Drug Discov. 2019, 18, 175–196. [Google Scholar] [CrossRef]
- Milling, L.; Zhang, Y.; Irvine, D.J. Delivering safer immunotherapies for cancer. Adv. Drug Deliv. Rev. 2017, 15, 79–101. [Google Scholar] [CrossRef] [PubMed]
- Vanneman, M.; Dranoff, G. Combining immunotherapy and targeted therapies in cancer treatment. Nat. Rev. Cancer. 2012, 12, 237–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conniot, J.; Silva, J.M.; Fernandes, J.G.; Silva, L.; Gaspar, R.; Ebrocchini, S.; Florindo, H.F.; Barata, T.S. Cancer immunotherapy: Nanodelivery approaches for immune cell targeting and tracking. Front. Chem. 2014, 2, 105. [Google Scholar] [CrossRef] [Green Version]
- Cai, S.S.; Li, T.; Akinade, T.; Zhu, Y.; Leong, K.W. Drug delivery carriers with therapeutic functions. Adv. Drug Deliv. Rev. 2021, 176, 113884. [Google Scholar] [CrossRef] [PubMed]
- Roterman, I.; Konieczny, L. Self-assembled molecules–New kind of protein ligands. In Supramolecular Ligands, 1st ed.; Springer: Cham, Switzerland, 2017. [Google Scholar] [CrossRef]
- Piekarska, B.; Skowronek, M.; Rybarska, J.; Stopa, B.; Roterman, I.; Konieczny, L. Congo red-stabilized intermediates in the lambda light chain transition from native to molten state. Biochimie 1996, 78, 183–189. [Google Scholar] [CrossRef]
- Jagusiak, A.; Panczyk, T. Interaction of Congo Red, Evans Blue and Titan Yellow with doxorubicin in aqueous solutions. A molecular dynamics study. J. Mol. Liquids 2019, 279, 640–648. [Google Scholar] [CrossRef]
- Piekarska, B.; Konieczny, L.; Rybarska, J.; Stopa, B.; Zemanek, G.; Szneler, E.; Król, M.; Nowak, M.; Roterman, I. Heat-induced formation of a specific binding site for self-assembled Congo red in the V domain of immunoglobulin L chain lambda. Biopolymers 2001, 59, 446–456. [Google Scholar] [CrossRef]
- Stopa, B.; Rybarska, J.; Drozd, A.; Konieczny, L.; Król, M.; Lisowski, M.; Piekarska, B.; Roterman, I.; Spólnik, P.; Zemanek, G. Albumin binds self-assembling dyes as specific polymolecular ligands. Int. J. Biol. Macromol. 2006, 40, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Peters, T., Jr. All about Albumin: Biochemistry, Genetics, and Medical Applications; Academic Press: San Diego, CA, USA, 1996. [Google Scholar] [CrossRef]
- Bertucci, C.; Domenici, E. Reversible and covalent binding of drugs to human serum albumin: Methodological approaches and physiological relevance. Curr. Med. Chem. 2002, 9, 1463–1481. [Google Scholar] [CrossRef]
- Yamasaki, K.; Chuang, V.T.; Maruyama, T.; Otagiri, M. Albumin-drug interaction and its clinical implication. Biochim. Biophys. Acta. 2013, 1830, 5435–5443. [Google Scholar] [CrossRef]
- Otagiri, M. A molecular functional study on the interactions of drugs with plasma proteins. Drug Metab. Pharmacokinet. 2005, 20, 309–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ascenzi, P.; Bocedi, A.; Notari, S.; Fanali, G.; Fesce, R.; Fasano, M. Allosteric modulation of drug binding to human serum albumin. Mini Rev. Med. Chem. 2006, 6, 483–489. [Google Scholar] [CrossRef]
- Yang, F.; Zhang, Y.; Liang, H. Interactive Association of Drugs Binding to Human Serum Albumin. Int. J. Mol. Sci. 2014, 15, 3580–3595. [Google Scholar] [CrossRef]
- Atwood, J.L.; Steed, J.W. Encyclopedia of Supramolecular Chemistry; CRC Press Tylor&Francis Group: Boca Raton, FL, USA, 2004. [Google Scholar] [CrossRef]
- Chattopadhyay, D.P. Chapter-Chemistry of dyeing. In Handbook of Textile and Industrial Dyeing; Woodhead Publishing Series in Textiles; Woodhead Publishing: Sawston, Cambridge, UK, 2011; Volume 1, pp. 150–183. [Google Scholar] [CrossRef]
- Brown, M.A.; Mitar, D.A.; Whitworth, J.A. Measurement of Plasma Volume in Pregnancy. Clin. Sci. 1992, 83, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Natascha, G.A.; Zeinab, Y.M.; Sarah, Y.Y.; Jerome, W.B. Chapter Four-Endothelial Protrusions in Junctional Integrity and Barrier Function; Belvitch, P., Dudek, S., Eds.; Current Topics in Membranes; Academic Press: Cambridge, MA, USA, 2018; Volume 82, pp. 93–140. [Google Scholar] [CrossRef]
- Grohe, M.E. Chapter 9—Clinical Evaluation of Adults. Dysphagia Clinical management in Adults and Children, 3rd ed.; Grohe, M.G., Crary, M.A., Eds.; Mosby; Elsevier: St. Louis, MI, USA, 2021; pp. 149–178. [Google Scholar] [CrossRef]
- Spolnik, P.; Stopa, B.; Piekarska, B.; Jagusiak, A.; Konieczny, L.; Rybarska, J.; Król, M.; Roterman, I.; Urbanowicz, B.; Zieba-Palus, J. The use of rigid, fibrillar Congo red nanostructures for scaffolding protein assemblies and inducing the formation of amyloid-like arrangement of molecules. Chem. Biol. Drug Des. 2007, 70, 491–501. [Google Scholar] [CrossRef]
- Ji, W.; Yuan, C.; Chakraborty, P.; Gilead, S.; Yan, X.; Gazit, E. Stoichiometry-controlled secondary structure transition of amyloid-derived supramolecular dipeptide co-assemblies. Commun. Chem. 2019, 2, 65. [Google Scholar] [CrossRef] [Green Version]
- Krol, M.; Roterman, I.; Piekarska, B.; Konieczny, L.; Rybarska, J.; Stopa, B.; Spolnik, P. Analysis of Correlated Domain motions in IgG light chain reveals possible mechanisms of immunological signal transduction. Proteins Struct. Funct. Bioinform. 2005, 59, 545–554. [Google Scholar] [CrossRef]
- Piekarska, B.; Drozd, A.; Konieczny, L.; Krol, M.; Jurkowski, W.; Roterman, I.; Spolnik, P.; Stopa, B.; Rybarska, J. The indirect generation of long-distance structural changes in antibodies upon their binding to antigen. Chem. Biol. Drug Des. 2006, 68, 276–283. [Google Scholar] [CrossRef]
- Krol, M.; Roterman, I.; Drozd, A.; Konieczny, L.; Piekarska, B.; Rybarska, J.; Spólnik, P.; Stopa, B. The increased flexibility of CDR loops generated in antibodies by Congo red complexation favors antigen binding. J. Biomol. Struct. Dyn. 2006, 23, 407–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, R.B. The anthracyclines: Will we ever find a better doxorubicin? Semin. Oncol. 1992, 19, e670–e686. [Google Scholar]
- Zhu, J.; Liao, L.; Bian, X.; Kong, J.; Yang, P.; Liu, B. pH-controlled delivery of doxorubicin to cancer cells, based on small mesoporous carbon nanospheres. Small 2012, 8, 2715–2720. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.Y.; Teng, M.C.; Lu, M.; Liang, H.F.; Lee, Y.R.; Yen, C.C.; Liang, M.L.; Wong, T.T. Treating glioblastoma multiforme with selective high-dose liposomal doxorubicin chemotherapy, induced by repeated focused ultrasound. Int. J. Nanomed. 2012, 7, 965–974. [Google Scholar] [CrossRef] [Green Version]
- Konieczny, L.; Piekarska, B.; Rybarska, J.; Skowronek, M.; Stopa, B.; Tabor, B.; Dabros, W.; Pawlicki, R.; Roterman, I. The use of Congo Red as a lyotropic liquid crystal to carry stain in a model immunotargeting system–microscopic studies. Folia Histochem. Cytobiol. 1997, 35, 203–210. [Google Scholar]
- Schellman, J.A.; Reese, H.R. Extensions to the theory of intercalation. Biopolymers 1995, 39, 161–171. [Google Scholar] [CrossRef]
- Tuite, E.; Kelly, J.M. The interaction of methylene blue, azure B, and thionine with DNA: Formation of complexes with polynucleotides as model systems. Biopolymers 1994, 35, 419–433. [Google Scholar] [CrossRef]
- Konieczny, L.; Piekarska, B.; Rybarska, J.; Stopa, B.; Krzykwa, B.; Noworolski, J.; Pawlicki, R.; Roterman, I. Bis azo dye liquid crystalline micelles as possible drug carriers in immunotergeting technique. J. Physiol. Pharmacol. 1994, 45, 441–454. [Google Scholar]
- Jagusiak, A.; Chlopas, K.; Zemanek, G.; Jemiola-Rzeminska, M.; Piekarska, B.; Stopa, B.; Panczyk, T. Self-assembled supramolecular ribbon-like structures complexed to single-walled carbon nanotubes as possible anticancer drug delivery systems. Int. J. Mol. Sci. 2019, 20, 2064. [Google Scholar] [CrossRef] [Green Version]
- Ptak-Kaczor, M.; Kwiecinska, K.; Korchowiec, J.; Chlopas, K.; Banas, M.; Roterman, I.; Jagusiak, A. Structure and Location of Protein Sites Binding Self-Associated Congo Red Molecules with Intercalated Drugs as Compact Ligands-Theoretical Studies. Biomolecules 2021, 11, 501. [Google Scholar] [CrossRef]
- Laginha, K.M.; Verwoert, S.; Charrois, G.J.R.; Allen, T.M. Determination of doxorubicin levels in whole tumor and tumor nuclei in murine breast cancer tumors. Clin. Cancer Res. 2005, 11, 6944–6949. [Google Scholar] [CrossRef] [Green Version]
- Sparreboom, A.; Scripture, C.D.; Trieu, V.; Williams, P.J.; De, T.Y.A.; Beals, B.; Figg, W.D.; Hawkins, M.; Desai, N. Comparative preclinical and clinical pharmacokinetics of a cremophor-free, nanoparticle albumin-bound paclitaxel (abi-007) and paclitaxel formulated in cremophor (taxol). Clin. Cancer Res. 2005, 11, 4136–4143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skowronek, M.; Stopa, B.; Konieczny, L.; Rybarska, J.; Piekarska, B.; Szneler, E.; Bakalarski, G.; Roterman, I. Self-assembly of Congo Red—A theoretical and experimental approach to identify its supramolecular organization in water and salt solutions. Biopolymers 1998, 46, 267–281. [Google Scholar] [CrossRef]
- Kwiecińska, K.; Stachowicz-Kuśnierz, A.; Jagusiak, A.; Roterman, I.; Korchowiec, J. Impact of Doxorubicin on Self-Organization of Congo Red: Quantum Chemical Calculations and Molecular Dynamics Simulations. ACS Omega 2020, 5, 19377–19384. [Google Scholar] [CrossRef]
- Jagusiak, A.; Chlopas, K.; Zemanek, G.; Wolski, P.; Panczyk, T. Controlled Release of Doxorubicin from the Drug Delivery Formulation Composed of Single-Walled Carbon Nanotubes and Congo Red: A Molecular Dynamics Study and Dynamic Light Scattering Analysis. Pharmaceutics 2020, 12, 622. [Google Scholar] [CrossRef] [PubMed]
- Spólnik, P.; Król, M.; Stopa, B.; Konieczny, L.; Piekarska, B.; Rybarska, J.; Zemanek, G.; Jagusiak, A.; Piwowar, P.; Szoniec, G. Influence of the electric field on supramolecular structure and properties of amyloid-specific reagent Congo Red. Eur. Biophys. J. 2011, 40, 1187–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elzoghby, A.O.; Samy, W.M.; Elgindy, N.A. Albumin-based nanoparticles as potential controlled release drug delivery systems. J. Control. Release 2012, 157, 168–182. [Google Scholar] [CrossRef]
- Karimi, M.; Bahrami, S.; Ravari, S.B.; Zangabad, P.S.; Mirshekari, H.; Bozorgomid, M.; Shahreza, S.; Sori, M.; Hamblin, M.R. Albumin nano-structures as advanced drug delivery systems. Expert Opin. Drug Deliv. 2016, 13, 1609–1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsadek, B.; Kratz, F. Impact of albumin on drug delivery–new applications on the horizon. J. Control. Release 2012, 157, 4–28. [Google Scholar] [CrossRef] [PubMed]
- Kouchakzadeh, H.; Safavi, M.S.; Shojaosadati, S.A. Efficient delivery of therapeutic agents by using targeted albumin nanoparticles. Adv. Protein Chem. Struct. Biol. 2015, 98, 121–143. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Li, Y. Physicochemical characteristics of nanoparticles affect circulation, biodistribution, cellular internalization, and trafficking. Small 2013, 9, 1521–1532. [Google Scholar] [CrossRef]
- Greish, K. Enhanced permeability and retention (EPR) effect for anticancer nanomedicine drug targeting. In Cancer Nanotechnology; Grobmyer, S., Moudgil, B., Eds.; Humana Press: New York, NY, USA, 2010; pp. 624–625. [Google Scholar] [CrossRef]
- Agudelo, D.; Bourassa, P.; Bruneau, J.; Bérubé, G.; Asselin, E.; Tajmir-Riahi, H.A. Probing the binding sites of antibiotic drugs doxorubicin and N-(trifluoroacetyl) doxorubicin with human and bovine serum albumins. PLoS ONE 2012, 7, e43814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Park, S.; Kim, J.B.; Kim, J.; Kim, H. Entrapped doxorubicin nanoparticles for the treatment of metastatic anoikis-resistant cancer cells. Cancer Lett. 2013, 332, 110–119. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CR | Dox | CR-Dox (2:1 Molar Ratio) | |
|---|---|---|---|
| Elution volume (Ve) (mL) | 2.3 | 0.9 | 0.3 |
| Mixtures | |||||||
|---|---|---|---|---|---|---|---|
| Elution Volume (Ve) | BSA-CR-Dox | Lλ-CR-Dox | CR-Dox | BSA-CR | Lλ-CR | BSA | Lλ |
| BSA (mL) | 0.3 | - | - | 0.6 | - | 0.7 | - |
| Lλ (mL) | - | 0.8 | - | - | 0.9 | - | 1 |
| CR (mL) | 0.3 | 0.4 | 0.3 | 0.6 | 0.8 | - | - |
| Dox (mL) | 0.3 | 0.4 | 0.3 | - | - | - | - |
| Complex formation: | YES | NO | YES | YES | ± | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jagusiak, A.; Chłopaś, K.; Zemanek, G.; Kościk, I.; Roterman, I. Interaction of Supramolecular Congo Red and Congo Red-Doxorubicin Complexes with Proteins for Drug Carrier Design. Pharmaceutics 2021, 13, 2027. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13122027
Jagusiak A, Chłopaś K, Zemanek G, Kościk I, Roterman I. Interaction of Supramolecular Congo Red and Congo Red-Doxorubicin Complexes with Proteins for Drug Carrier Design. Pharmaceutics. 2021; 13(12):2027. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13122027
Chicago/Turabian StyleJagusiak, Anna, Katarzyna Chłopaś, Grzegorz Zemanek, Izabela Kościk, and Irena Roterman. 2021. "Interaction of Supramolecular Congo Red and Congo Red-Doxorubicin Complexes with Proteins for Drug Carrier Design" Pharmaceutics 13, no. 12: 2027. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13122027