
Development of an Immunochromatographic Strip Using Conjugated Gold Nanoparticles for the Rapid Detection of Klebsiella pneumoniae Causing Neonatal Sepsis

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Bioinformatic Analyses of K. pneumoniae Proteome to Identify Potential Targets
2.3. Validation of the Expression of the Selected Candidates via RT-RT-PCR
2.4. Synthesis of the Target Peptide
2.5. Immunization and Purification of Polyclonal Antibodies
2.6. Synthesis and Characterization of Gold Nanoparticles (GNPs)
2.7. Preparation of GNP-Antibody Conjugate
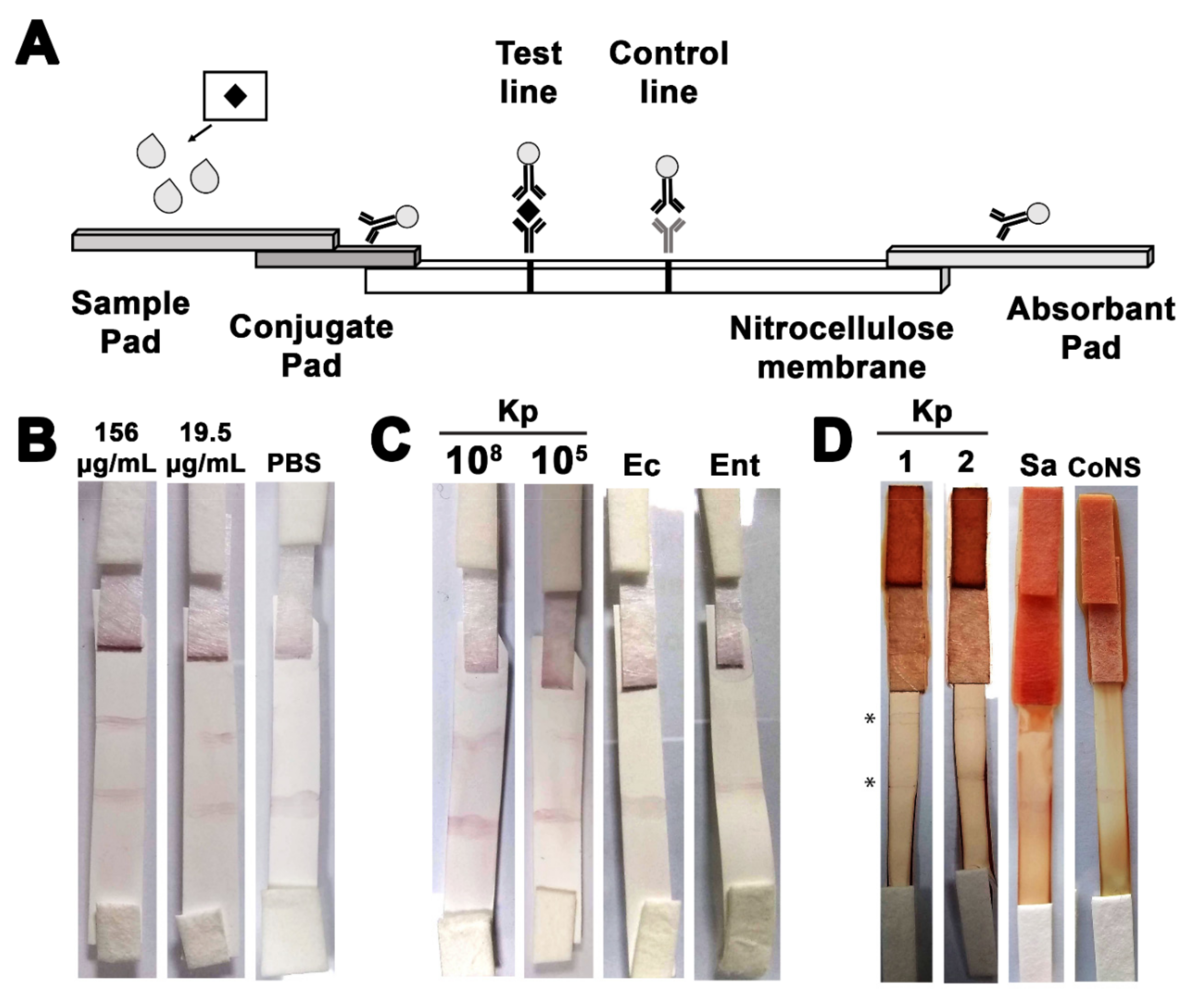
2.8. Assembly of the Immunochromatographic Strip
2.9. Determination of the Sensitivity and Specificity of the Constructed ICS
2.10. Assessment of the Ability of the Developed ICS to Detect K. Pneumoniae in Neonatal Sepsis Blood Samples
3. Results
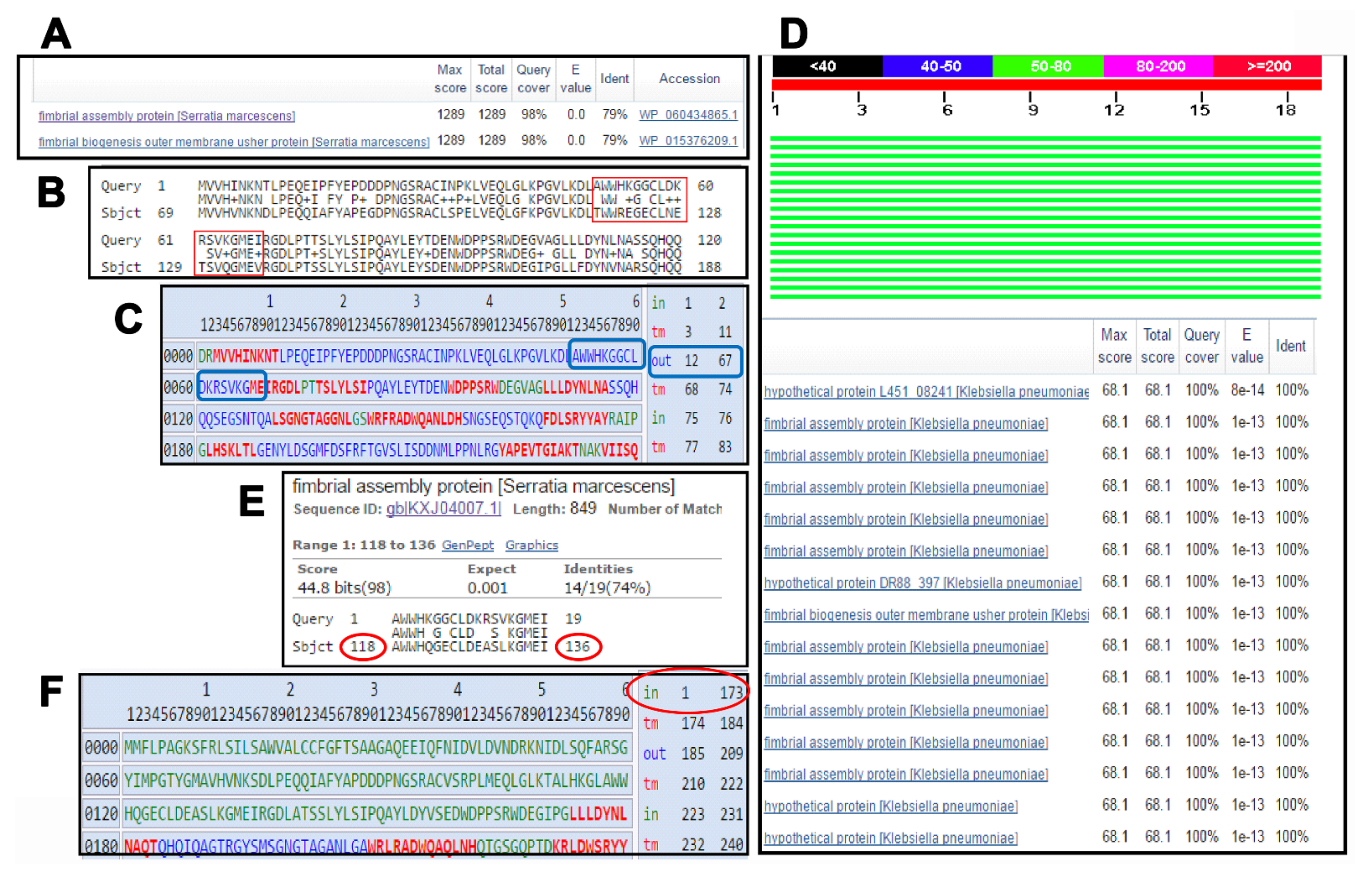
3.1. In Silico Identification of Promising Protein/Peptide Candidates
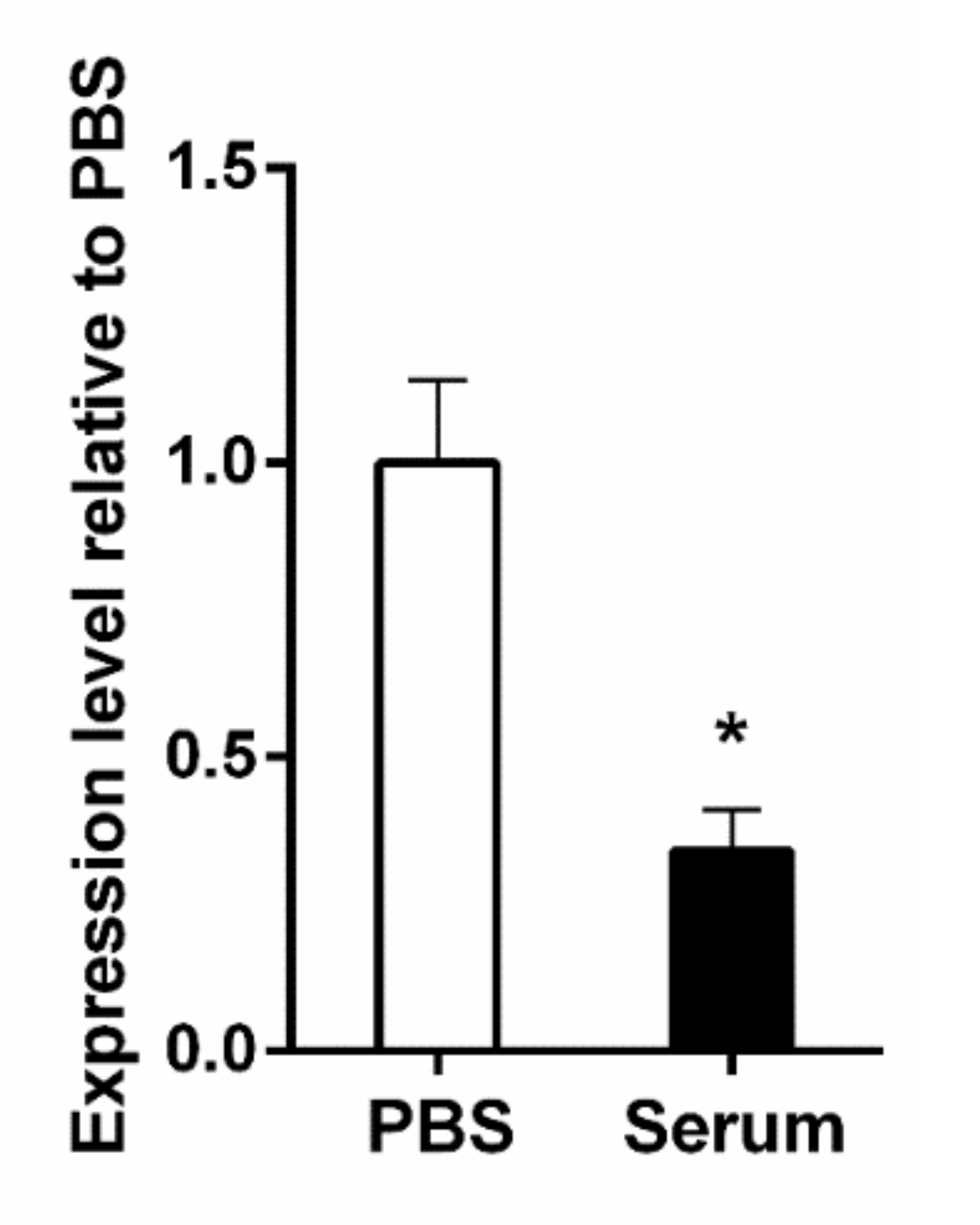
3.2. The KFJ72693 Encoding Gene (DR88_397) Is Expressed under Relevant Conditions
3.3. Evaluation of the Synthesized Peptide
3.4. The KP_397(50-68) Peptide Is Moderately Immunogenic in Mice
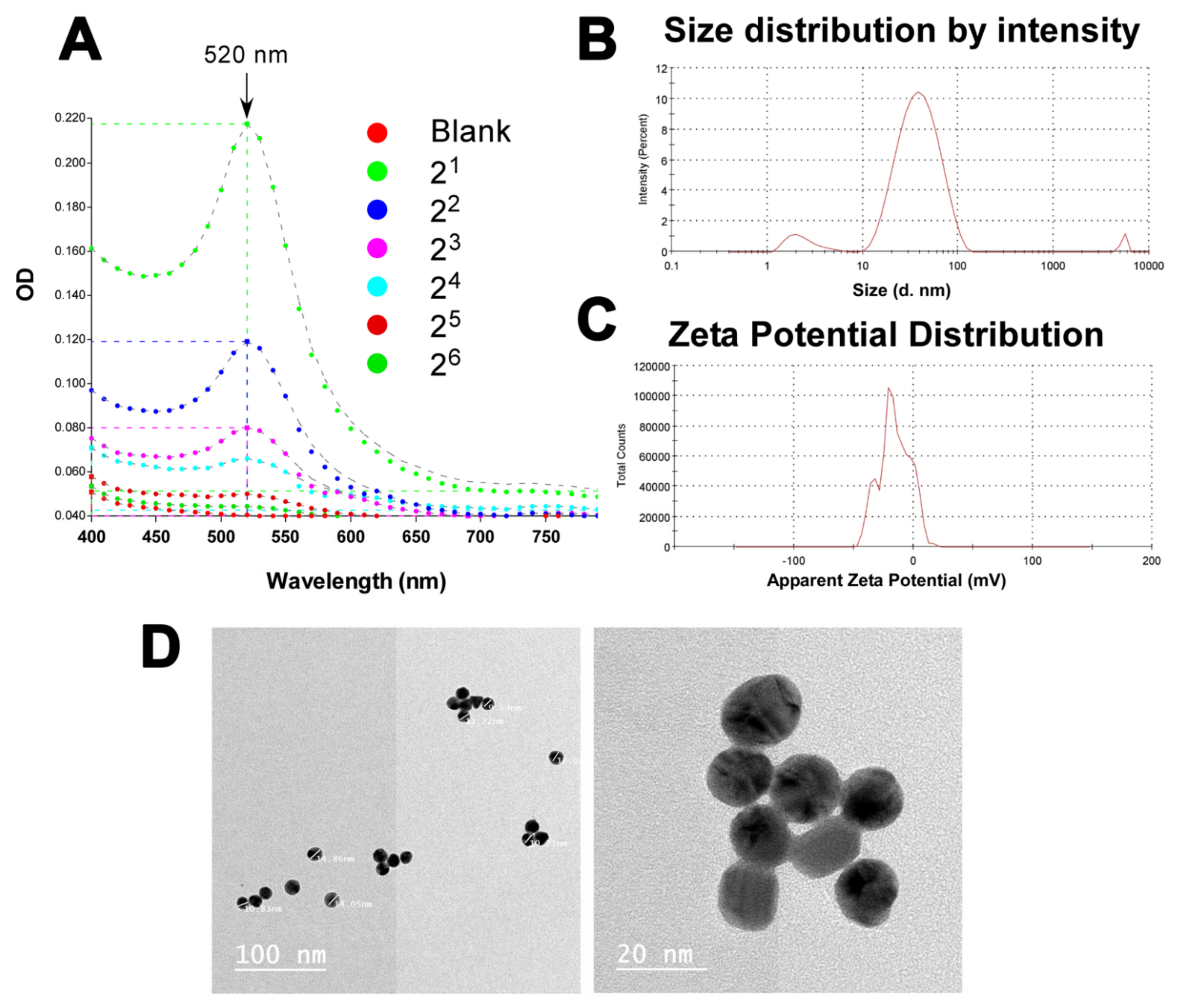
3.5. Successful Production of Gold Suspension in the Nanometer Range
3.6. The Developed ICS Specifically Detects K. pneumoniae in Neonatal Sepsis Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goldstein, B.; Giroir, B.; Randolph, A. International Consensus Conference on Pediatric, S. International pediatric sepsis consensus conference: Definitions for sepsis and organ dysfunction in pediatrics. Pediatr. Crit. Care Med. 2005, 6, 2–8. [Google Scholar] [CrossRef] [Green Version]
- Wynn, J.L. Defining neonatal sepsis. Curr. Opin. Pediatr. 2016, 28, 135–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudd, K.E.; Johnson, S.C.; Agesa, K.M.; Shackelford, K.A.; Tsoi, D.; Kievlan, D.R.; Colombara, D.V.; Ikuta, K.S.; Kissoon, N.; Finfer, S.; et al. Global, regional, and national sepsis incidence and mortality, 1990–2017: Analysis for the Global Burden of Disease Study. Lancet 2020, 395, 200–211. [Google Scholar] [CrossRef] [Green Version]
- Weiss, S.L.; Fitzgerald, J.C.; Pappachan, J.; Wheeler, D.; Jaramillo-Bustamante, J.C.; Salloo, A.; Singhi, S.C.; Erickson, S.; Roy, J.A.; Bush, J.L.; et al. Global epidemiology of pediatric severe sepsis: The sepsis prevalence, outcomes, and therapies study. Am. J. Respir. Crit. Care Med. 2015, 191, 1147–1157. [Google Scholar] [CrossRef] [PubMed]
- McGovern, M.; Giannoni, E.; Kuester, H.; Turner, M.A.; van den Hoogen, A.; Bliss, J.M.; Koenig, J.M.; Keij, F.M.; Mazela, J.; Finnegan, R.; et al. Challenges in developing a consensus definition of neonatal sepsis. Pediatr. Res. 2020, 88, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Fleischmann-Struzek, C.; Goldfarb, D.M.; Schlattmann, P.; Schlapbach, L.J.; Reinhart, K.; Kissoon, N. The global burden of paediatric and neonatal sepsis: A systematic review. Lancet Respir. Med. 2018, 6, 223–230. [Google Scholar] [CrossRef]
- Kilpatrick, C.; Saito, H.; Allegranzi, B.; Pittet, D. Preventing sepsis in health care–It’s in your hands: A World Health Organization call to action. J. Infect. Prev. 2018, 19, 104–106. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.; Wong, J.J.; Sultana, R.; Koh, J.; Jit, M.; Mok, Y.H.; Lee, J.H. Global Case-Fatality Rates in Pediatric Severe Sepsis and Septic Shock: A Systematic Review and Meta-analysis. JAMA Pediatr. 2019, 173, 352–362. [Google Scholar] [CrossRef]
- Group, I.C.; Brocklehurst, P.; Farrell, B.; King, A.; Juszczak, E.; Darlow, B.; Haque, K.; Salt, A.; Stenson, B.; Tarnow-Mordi, W. Treatment of neonatal sepsis with intravenous immune globulin. N. Engl. J. Med. 2011, 365, 1201–1211. [Google Scholar] [CrossRef] [Green Version]
- Barton, L.; Hodgman, J.E.; Pavlova, Z. Causes of death in the extremely low birth weight infant. Pediatrics 1999, 103, 446–451. [Google Scholar] [CrossRef]
- Shane, A.L.; Sanchez, P.J.; Stoll, B.J. Neonatal sepsis. Lancet 2017, 390, 1770–1780. [Google Scholar] [CrossRef]
- Moges, F.; Eshetie, S.; Yeshitela, B.; Abate, E. Bacterial etiologic agents causing neonatal sepsis and associated risk factors in Gondar, Northwest Ethiopia. BMC Pediatr. 2017, 17, 137. [Google Scholar] [CrossRef] [Green Version]
- Tessema, B.; Lippmann, N.; Knupfer, M.; Sack, U.; Konig, B. Antibiotic Resistance Patterns of Bacterial Isolates from Neonatal Sepsis Patients at University Hospital of Leipzig, Germany. Antibiotics 2021, 10, 323. [Google Scholar] [CrossRef] [PubMed]
- Khalil, N.; Blunt, H.B.; Li, Z.; Hartman, T. Neonatal early onset sepsis in Middle Eastern countries: A systematic review. Arch. Dis. Child. 2020, 105, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Rafi, M.A.; Miah, M.M.Z.; Wadood, M.A.; Hossain, M.G. Risk factors and etiology of neonatal sepsis after hospital delivery: A case-control study in a tertiary care hospital of Rajshahi, Bangladesh. PLoS ONE 2020, 15, e0242275. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.W.; Blau, D.M.; Bassat, Q.; Onyango, D.; Kotloff, K.L.; Arifeen, S.E.; Mandomando, I.; Chawana, R.; Baillie, V.L.; Akelo, V.; et al. Initial findings from a novel population-based child mortality surveillance approach: A descriptive study. Lancet Glob. Health 2020, 8, e909–e919. [Google Scholar] [CrossRef]
- Hassuna, N.A.; AbdelAziz, R.A.; Zakaria, A.; Abdelhakeem, M. Extensively-Drug Resistant Klebsiella pneumoniae Recovered From Neonatal Sepsis Cases From a Major NICU in Egypt. Front. Microbiol. 2020, 11, 1375. [Google Scholar] [CrossRef]
- Ghaith, D.M.; Zafer, M.M.; Said, H.M.; Elanwary, S.; Elsaban, S.; Al-Agamy, M.H.; Bohol, M.F.F.; Bendary, M.M.; Al-Qahtani, A.; Al-Ahdal, M.N. Genetic diversity of carbapenem-resistant Klebsiella Pneumoniae causing neonatal sepsis in intensive care unit, Cairo, Egypt. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 583–591. [Google Scholar] [CrossRef]
- Morad, E.A.; Rabie, R.A.; Almalky, M.A.; Gebriel, M.G. Evaluation of Procalcitonin, C-Reactive Protein, and Interleukin-6 as Early Markers for Diagnosis of Neonatal Sepsis. Int. J. Microbiol. 2020, 2020, 8889086. [Google Scholar] [CrossRef] [PubMed]
- Almohammady, M.N.; Eltahlawy, E.M.; Reda, N.M. Pattern of bacterial profile and antibiotic susceptibility among neonatal sepsis cases at Cairo University Children Hospital. J. Taibah Univ. Med. Sci. 2020, 15, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Kellogg, J.A.; Ferrentino, F.L.; Goodstein, M.H.; Liss, J.; Shapiro, S.L.; Bankert, D.A. Frequency of low level bacteremia in infants from birth to two months of age. Pediatr. Infect. Dis. J. 1997, 16, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Farahbakhsh, N.; Shastri, S.; Sharma, P. Biomarkers for diagnosis of neonatal sepsis: A literature review. J. Matern.-Fetal Neonatal Med. 2018, 31, 1646–1659. [Google Scholar] [CrossRef]
- Oeser, C.; Pond, M.; Butcher, P.; Bedford Russell, A.; Henneke, P.; Laing, K.; Planche, T.; Heath, P.T.; Harris, K. PCR for the detection of pathogens in neonatal early onset sepsis. PLoS ONE 2020, 15, e0226817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, X.; Pu, X.; Jun, P.; Zhu, X.; Zhu, D.; Chen, M. Rapid and quantitative detection of C-reactive protein using quantum dots and immunochromatographic test strips. Int. J. Nanomed. 2014, 9, 5619–5626. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Li, X.; Yang, G.; Fan, J.; Tang, Y.; Zhao, J.; Long, X.; Guo, S.; Zhao, Z.; Liu, Y.; et al. Serological immunochromatographic approach in diagnosis with SARS-CoV-2 infected COVID-19 patients. J. Infect. 2020, 81, e28–e32. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, S.A.; Samir, T.M.; Helmy, O.M.; Elhosseiny, N.M.; Ali, A.A.; El-Kholy, A.A.; Attia, A.S. A Novel Surface-Exposed Polypeptide Is Successfully Employed as a Target for Developing a Prototype One-Step Immunochromatographic Strip for Specific and Sensitive Direct Detection of Staphylococcus aureus Causing Neonatal Sepsis. Biomolecules 2020, 10, 1580. [Google Scholar] [CrossRef] [PubMed]
- Yu, N.Y.; Wagner, J.R.; Laird, M.R.; Melli, G.; Rey, S.; Lo, R.; Dao, P.; Sahinalp, S.C.; Ester, M.; Foster, L.J.; et al. PSORTb 3.0: Improved protein subcellular localization prediction with refined localization subcategories and predictive capabilities for all prokaryotes. Bioinformatics 2010, 26, 1608–1615. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.; Clamp, M.; Barton, G.J. Jalview Version 2--a multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [Green Version]
- Bagos, P.G.; Liakopoulos, T.D.; Spyropoulos, I.C.; Hamodrakas, S.J. PRED-TMBB: A web server for predicting the topology of beta-barrel outer membrane proteins. Nucleic Acids Res. 2004, 32, W400–404. [Google Scholar] [CrossRef] [PubMed]
- Fleri, W.; Paul, S.; Dhanda, S.K.; Mahajan, S.; Xu, X.; Peters, B.; Sette, A. The Immune Epitope Database and Analysis Resource in Epitope Discovery and Synthetic Vaccine Design. Front. Immunol. 2017, 8, 278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elder, B.L.; Boraker, D.K.; Fives-Taylor, P.M. Whole-bacterial cell enzyme-linked immunosorbent assay for Streptococcus sanguis fimbrial antigens. J. Clin. Microbiol. 1982, 16, 141–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailes, J.; Mayoss, S.; Teale, P.; Soloviev, M. Gold nanoparticle antibody conjugates for use in competitive lateral flow assays. Methods Mol. Biol. 2012, 906, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Khalil, M.A.; Azzazy, H.M.; Attia, A.S.; Hashem, A.G. A sensitive colorimetric assay for identification of Acinetobacter baumannii using unmodified gold nanoparticles. J. Appl. Microbiol. 2014, 117, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Shnoudeh, A.J.; Hamad, I.; Abdo, R.W.; Qadumii, L.; Jaber, A.Y.; Surchi, H.S.; Alkelany, S.Z. Synthesis, characterization, and applications of metal nanoparticles. In Biomaterials and Bionanotechnology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 527–612. [Google Scholar]
- El Assasy, A.E.I.; Younes, N.F.; Makhlouf, A.I.A. Enhanced Oral Absorption of Amisulpride Via a Nanostructured Lipid Carrier-Based Capsules: Development, Optimization Applying the Desirability Function Approach and In Vivo Pharmacokinetic Study. AAPS PharmSciTech 2019, 20, 82. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Roberts, D.; Wood, K.E.; Light, B.; Parrillo, J.E.; Sharma, S.; Suppes, R.; Feinstein, D.; Zanotti, S.; Taiberg, L.; et al. Duration of hypotension before initiation of effective antimicrobial therapy is the critical determinant of survival in human septic shock. Crit. Care Med. 2006, 34, 1589–1596. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Ji, R.; Chai, R.; Yuan, N.; Zhang, J.; Jing, Y.; Yang, M.; Zhang, L.; Hong, Y.; Lin, J.; et al. A novel fluorescence immunochromatographic assay strip for the diagnosis of schistosomiasis japonica. Parasit. Vectors 2021, 14, 8. [Google Scholar] [CrossRef] [PubMed]
- Sadaow, L.; Sanpool, O.; Rodpai, R.; Yamasaki, H.; Ittiprasert, W.; Mann, V.H.; Brindley, P.J.; Maleewong, W.; Intapan, P.M. Development of an Immunochromatographic Point-of-Care Test for Serodiagnosis of Opisthorchiasis and Clonorchiasis. Am. J. Trop. Med. Hyg. 2019, 101, 1156–1160. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, D.; Amgalanbaatar, T.; Davaasuren, B.; Molefe, N.I.; Battur, B.; Battsetseg, B.; Inoue, N.; Yokoyama, N.; Suganuma, K. The utility of an rTeGM6-4r-based immunochromatographic test for the serological diagnosis of non-tsetse-transmitted equine trypanosomosis in rural areas of Mongolia. Parasit. Res. 2018, 117, 2913–2919. [Google Scholar] [CrossRef] [PubMed]
- Guedez-Lopez, G.V.; Alguacil-Guillen, M.; Gonzalez-Donapetry, P.; Bloise, I.; Tornero-Marin, C.; Gonzalez-Garcia, J.; Mingorance, J.; Garcia-Rodriguez, J.; Group, S.A.-C.-W. Evaluation of three immunochromatographic tests for rapid detection of antibodies against SARS-CoV-2. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 2289–2297. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Duan, Y.; Zhang, D.; Zhang, S.; Li, Z.; Wang, X.; Yang, P. Development of an immunochromatographic kit for rapid detection of human influenza B virus infection. Clin. Lab. 2014, 60, 815–819. [Google Scholar] [CrossRef]
- Wong, C.L.; Yong, C.Y.; Ong, H.K.; Ho, K.L.; Tan, W.S. Advances in the Diagnosis of Foot-and-Mouth Disease. Front. Vet. Sci. 2020, 7, 477. [Google Scholar] [CrossRef] [PubMed]
- Guswanto, A.; Allamanda, P.; Mariamah, E.S.; Munkjargal, T.; Tuvshintulga, B.; Takemae, H.; Sivakumar, T.; AbouLaila, M.; Terkawi, M.A.; Ichikawa-Seki, M.; et al. Evaluation of immunochromatographic test (ICT) strips for the serological detection of Babesia bovis and Babesia bigemina infection in cattle from Western Java, Indonesia. Vet. Parasit. 2017, 239, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hou, L.; Ye, J.; Liu, X.; Dan, H.; Jin, M.; Chen, H.; Cao, S. Development of a convenient immunochromatographic strip for the diagnosis of infection with Japanese encephalitis virus in swine. J. Virol. Methods 2010, 168, 51–56. [Google Scholar] [CrossRef]
- Ngom, B.; Guo, Y.; Wang, X.; Bi, D. Development and application of lateral flow test strip technology for detection of infectious agents and chemical contaminants: A review. Anal. Bioanal. Chem. 2010, 397, 1113–1135. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wang, Y.; Weng, T.; Hu, C.; Wang, F.X.C.; Wu, Z.; Yu, D.; Lu, H.; Yao, H. Preparation of immunochromatographic strips for rapid detection of early secreted protein ESAT-6 and culture filtrate protein CFP-10 from Mycobacterium tuberculosis. Medicine 2017, 96, e9350. [Google Scholar] [CrossRef]
- Nagasawa, Y.; Kiku, Y.; Sugawara, K.; Yabusaki, N.; Oono, K.; Fujii, K.; Suzuki, T.; Maehana, K.; Hayashi, T. Rapid Staphylococcus aureus Detection From Clinical Mastitis Milk by Colloidal Gold Nanoparticle-Based Immunochromatographic Strips. Front. Vet. Sci. 2019, 6, 504. [Google Scholar] [CrossRef] [PubMed]
- Karakus, C.; Salih, B.A. Comparison of the lateral flow immunoassays (LFIA) for the diagnosis of Helicobacter pylori infection. J. Immunol. Methods 2013, 396, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Hao, H.; Li, J.; Wang, M.; Wang, Y.; Zhang, G.; Hu, Z. Colloidal gold immunochromatographic strip for rapid detection of Haemophilus influenzae. Sheng Wu Gong Cheng Xue Bao 2019, 35, 901–909. [Google Scholar] [CrossRef]
- Watchirs Smith, L.A.; Hillman, R.; Ward, J.; Whiley, D.M.; Causer, L.; Skov, S.; Donovan, B.; Kaldor, J.; Guy, R. Point-of-care tests for the diagnosis of Neisseria gonorrhoeae infection: A systematic review of operational and performance characteristics. Sex. Transm. Infect. 2013, 89, 320–326. [Google Scholar] [CrossRef]
- Purcell, A.W.; McCluskey, J.; Rossjohn, J. More than one reason to rethink the use of peptides in vaccine design. Nat. Rev. Drug Discov. 2007, 6, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Struve, C.; Bojer, M.; Krogfelt, K.A. Characterization of Klebsiella pneumoniae type 1 fimbriae by detection of phase variation during colonization and infection and impact on virulence. Infect. Immun. 2008, 76, 4055–4065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grodzki, A.C.; Berenstein, E. Introduction to the purification of antibodies. Methods Mol. Biol. 2010, 588, 11–13. [Google Scholar] [CrossRef] [PubMed]
- Serebrennikova, K.; Samsonova, J.; Osipov, A. Hierarchical Nanogold Labels to Improve the Sensitivity of Lateral Flow Immunoassay. Nano-Micro Lett. 2018, 10, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khlebtsov, B.; Khlebtsov, N. Enhanced solid-phase immunoassay using gold nanoshells: Effect of nanoparticle optical properties. Nanotechnology 2008, 19, 435703. [Google Scholar] [CrossRef] [PubMed]
- Dykman, L.A.; Khlebtsov, N.G. Gold nanoparticles in biology and medicine: Recent advances and prospects. Acta. Nat. 2011, 3, 34–55. [Google Scholar] [CrossRef] [Green Version]
- Iconaru, S.L.; Prodan, A.M.; Buton, N.; Predoi, D. Structural Characterization and Antifungal Studies of Zinc-Doped Hydroxyapatite Coatings. Molecules 2017, 22, 604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Predoi, D.; Iconaru, S.L.; Deniaud, A.; Chevallet, M.; Michaud-Soret, I.; Buton, N.; Prodan, A.M. Textural, Structural and Biological Evaluation of Hydroxyapatite Doped with Zinc at Low Concentrations. Materials 2017, 10, 229. [Google Scholar] [CrossRef] [Green Version]
- Predoi, D.; Iconaru, S.L.; Predoi, M.V.; Motelica-Heino, M.; Buton, N.; Megier, C. Obtaining and characterizing thin layers of magnesium doped hydroxyapatite by dip coating procedure. Coatings 2020, 10, 510. [Google Scholar] [CrossRef]
- Tantra, R.; Schulze, P.; Quincey, P. Effect of nanoparticle concentration on zeta-potential measurement results and reproducibility. Particuology 2010, 8, 279–285. [Google Scholar] [CrossRef]
- Shin, H.S.; Kim, C.K.; Shin, K.S.; Chung, H.K.; Heo, T.R. Pretreatment of whole blood for use in immunochromatographic assays for hepatitis B virus surface antigen. Clin. Diagn. Lab. Immunol. 2001, 8, 9–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tominaga, T. Rapid detection of Klebsiella pneumoniae, Klebsiella oxytoca, Raoultella ornithinolytica and other related bacteria in food by lateral-flow test strip immunoassays. J. Microbiol. Methods 2018, 147, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, T. Rapid detection of coliform bacteria using a lateral flow test strip assay. J. Microbiol. Methods 2019, 160, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.H.; Lu, P.L.; Liu, E.Y.; Chen, Y.Y.; Lin, F.M.; Lin, Y.T.; Chang, F.Y.; Lin, J.C. Rapid identification of capsular serotype K1/K2 Klebsiella pneumoniae in pus samples from liver abscess patients and positive blood culture samples from bacteremia cases via an immunochromatographic strip assay. Gut Pathog. 2019, 11, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elhosseiny, N.M.; Samir, T.M.; Ali, A.A.; El-Kholy, A.A.; Attia, A.S. Development of an Immunochromatographic Strip Using Conjugated Gold Nanoparticles for the Rapid Detection of Klebsiella pneumoniae Causing Neonatal Sepsis. Pharmaceutics 2021, 13, 1141. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13081141
Elhosseiny NM, Samir TM, Ali AA, El-Kholy AA, Attia AS. Development of an Immunochromatographic Strip Using Conjugated Gold Nanoparticles for the Rapid Detection of Klebsiella pneumoniae Causing Neonatal Sepsis. Pharmaceutics. 2021; 13(8):1141. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13081141
Chicago/Turabian StyleElhosseiny, Noha M., Tamer M. Samir, Aliaa A. Ali, Amani A. El-Kholy, and Ahmed S. Attia. 2021. "Development of an Immunochromatographic Strip Using Conjugated Gold Nanoparticles for the Rapid Detection of Klebsiella pneumoniae Causing Neonatal Sepsis" Pharmaceutics 13, no. 8: 1141. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13081141