
Spatiotemporal Correlation Spectroscopy Reveals a Protective Effect of Peptide-Based GLP-1 Receptor Agonism against Lipotoxicity on Insulin Granule Dynamics in Primary Human β-Cells

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Pancreatic Islets
2.2. Insulin Secretion Assays
2.3. Immunostaining
2.4. Plasmid Transfection
2.5. Fluorescence Microscopy and iMSD Analysis
2.6. Cluster Similarity Analysis
2.7. ISGs Tracking Analysis
3. Results and Discussion
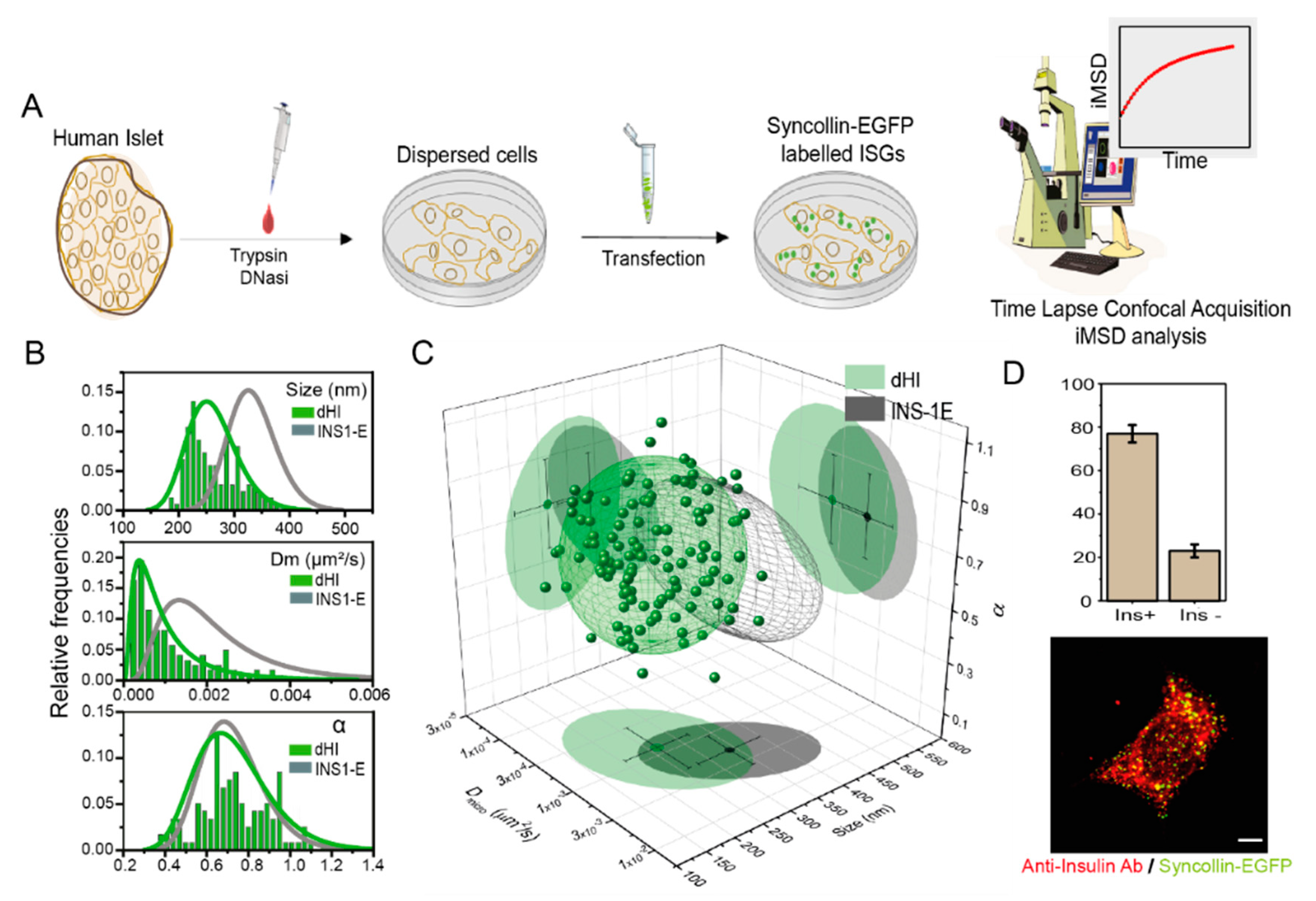
3.1. iMSD-Based Structural/Dynamic Fingerprint of ISGs from Human β-Cells
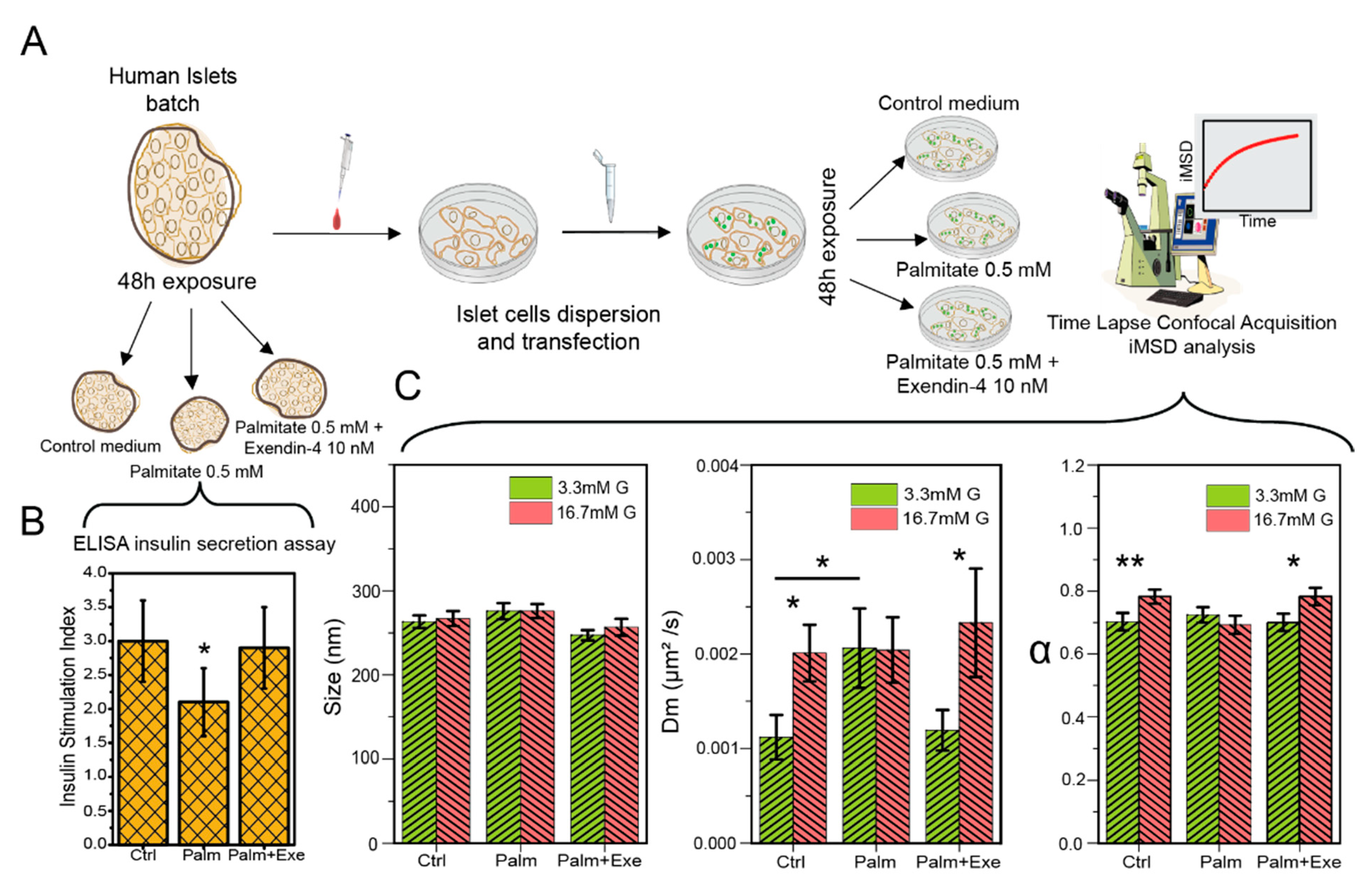
3.2. GLP-1 Agonism Effect on Human β-Cell ISGs under Lipotoxic Stress
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Kahn, S.E. The Relative Contributions of Insulin Resistance and Beta-Cell Dysfunction to the Pathophysiology of Type 2 Diabetes. Diabetologia 2003, 46, 3–19. [Google Scholar] [CrossRef] [Green Version]
- Kahn, S.E. The Importance of β-Cell Failure in the Development and Progression of Type 2 Diabetes. J. Clin. Endocrinol. Metab. 2001, 86, 4047–4058. [Google Scholar] [CrossRef] [Green Version]
- Oh, Y.S.; Bae, G.D.; Baek, D.J.; Park, E.-Y.; Jun, H.-S. Fatty Acid-Induced Lipotoxicity in Pancreatic Beta-Cells During Development of Type 2 Diabetes. Front. Endocrinol. 2018, 9, 384. [Google Scholar] [CrossRef]
- Sharma, R.B.; Alonso, L.C. Lipotoxicity in the Pancreatic Beta Cell: Not Just Survival and Function, but Proliferation as Well? Curr. Diabetes Rep. 2014, 14, 492. [Google Scholar] [CrossRef] [Green Version]
- Lytrivi, M.; Castell, A.L.; Poitout, V.; Cnop, M. Recent Insights Into Mechanisms of β-Cell Lipo- and Glucolipotoxicity in Type 2 Diabetes. J. Mol. Biol. 2020, 432, 1514–1534. [Google Scholar] [CrossRef]
- Baggio, L.L.; Drucker, D.J. Glucagon-like Peptide-1 Receptor Co-Agonists for Treating Metabolic Disease. Mol. Metab. 2021, 46, 101090. [Google Scholar] [CrossRef]
- Hinnen, D. Glucagon-Like Peptide 1 Receptor Agonists for Type 2 Diabetes. Diabetes Spectr. 2017, 30, 202–210. [Google Scholar] [CrossRef] [Green Version]
- Oh, Y.; Jun, H.-S. Effects of Glucagon-Like Peptide-1 on Oxidative Stress and Nrf2 Signaling. IJMS 2017, 19, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zummo, F.P.; Cullen, K.S.; Honkanen-Scott, M.; Shaw, J.A.M.; Lovat, P.E.; Arden, C. Glucagon-Like Peptide 1 Protects Pancreatic β-Cells From Death by Increasing Autophagic Flux and Restoring Lysosomal Function. Diabetes 2017, 66, 1272–1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lytrivi, M.; Ghaddar, K.; Lopes, M.; Rosengren, V.; Piron, A.; Yi, X.; Johansson, H.; Lehtiö, J.; Igoillo-Esteve, M.; Cunha, D.A.; et al. Combined Transcriptome and Proteome Profiling of the Pancreatic β-Cell Response to Palmitate Unveils Key Pathways of β-Cell Lipotoxicity. BMC Genom. 2020, 21, 590. [Google Scholar] [CrossRef] [PubMed]
- Prentki, M.; Peyot, M.-L.; Masiello, P.; Madiraju, S.R.M. Nutrient-Induced Metabolic Stress, Adaptation, Detoxification, and Toxicity in the Pancreatic β-Cell. Diabetes 2020, 69, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Tomas, A.; Jones, B.; Leech, C. New Insights into Beta-Cell GLP-1 Receptor and CAMP Signaling. J. Mol. Biol. 2020, 432, 1347–1366. [Google Scholar] [CrossRef]
- Suckale, J.; Solimena, M. The Insulin Secretory Granule as a Signaling Hub. Trends Endocrinol. Metab. 2010, 21, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Bogan, J.S.; Xu, Y.; Hao, M. Cholesterol Accumulation Increases Insulin Granule Size and Impairs Membrane Trafficking. Traffic 2012, 13, 1466–1480. [Google Scholar] [CrossRef] [Green Version]
- Wan, X.; Zinselmeyer, B.H.; Zakharov, P.N.; Vomund, A.N.; Taniguchi, R.; Santambrogio, L.; Anderson, M.S.; Lichti, C.F.; Unanue, E.R. Pancreatic Islets Communicate with Lymphoid Tissues via Exocytosis of Insulin Peptides. Nature 2018, 560, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Fava, E.; Dehghany, J.; Ouwendijk, J.; Müller, A.; Niederlein, A.; Verkade, P.; Meyer-Hermann, M.; Solimena, M. Novel Standards in the Measurement of Rat Insulin Granules Combining Electron Microscopy, High-Content Image Analysis and in Silico Modelling. Diabetologia 2012, 55, 1013–1023. [Google Scholar] [CrossRef] [Green Version]
- Ohara-Imaizumi, M.; Nishiwaki, C.; Kikuta, T.; Nagai, S.; Nakamichi, Y.; Nagamatsu, S. TIRF Imaging of Docking and Fusion of Single Insulin Granule Motion in Primary Rat Pancreatic β-Cells: Different Behaviour of Granule Motion between Normal and Goto–Kakizaki Diabetic Rat β-Cells. Biochem. J. 2004, 381, 13–18. [Google Scholar] [CrossRef]
- Ma, L.; Bindokas, V.P.; Kuznetsov, A.; Rhodes, C.; Hays, L.; Edwardson, J.M.; Ueda, K.; Steiner, D.F.; Philipson, L.H. Direct Imaging Shows That Insulin Granule Exocytosis Occurs by Complete Vesicle Fusion. Proc. Natl. Acad. Sci. USA 2004, 101, 9266–9271. [Google Scholar] [CrossRef] [Green Version]
- Seino, S.; Shibasaki, T.; Minami, K. Dynamics of Insulin Secretion and the Clinical Implications for Obesity and Diabetes. J. Clin. Investig. 2011, 121, 2118–2125. [Google Scholar] [CrossRef] [Green Version]
- Tabei, S.M.A.; Burov, S.; Kim, H.Y.; Kuznetsov, A.; Huynh, T.; Jureller, J.; Philipson, L.H.; Dinner, A.R.; Scherer, N.F. Intracellular Transport of Insulin Granules Is a Subordinated Random Walk. Proc. Natl. Acad. Sci. USA 2013, 110, 4911–4916. [Google Scholar] [CrossRef] [Green Version]
- Heaslip, A.T.; Nelson, S.R.; Lombardo, A.T.; Beck Previs, S.; Armstrong, J.; Warshaw, D.M. Cytoskeletal Dependence of Insulin Granule Movement Dynamics in INS-1 Beta-Cells in Response to Glucose. PLoS ONE 2014, 9, e109082. [Google Scholar] [CrossRef] [Green Version]
- Di Rienzo, C.; Gratton, E.; Beltram, F.; Cardarelli, F. Fast Spatiotemporal Correlation Spectroscopy to Determine Protein Lateral Diffusion Laws in Live Cell Membranes. Proc. Natl. Acad. Sci. USA 2013, 110, 12307–12312. [Google Scholar] [CrossRef] [Green Version]
- Digiacomo, L.; D’Autilia, F.; Durso, W.; Tentori, P.M.; Caracciolo, G.; Cardarelli, F. Dynamic Fingerprinting of Sub-Cellular Nanostructures by Image Mean Square Displacement Analysis. Sci. Rep. 2017, 7, 14836. [Google Scholar] [CrossRef] [Green Version]
- Digiacomo, L.; Digman, M.A.; Gratton, E.; Caracciolo, G. Development of an Image Mean Square Displacement (iMSD)-Based Method as a Novel Approach to Study the Intracellular Trafficking of Nanoparticles. Acta Biomater. 2016, 42, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Ferri, G.; Digiacomo, L.; D’Autilia, F.; Durso, W.; Caracciolo, G.; Cardarelli, F. Time-Lapse Confocal Imaging Datasets to Assess Structural and Dynamic Properties of Subcellular Nanostructures. Sci. Data 2018, 5, 180191. [Google Scholar] [CrossRef] [Green Version]
- Ferri, G.; Digiacomo, L.; Lavagnino, Z.; Occhipinti, M.; Bugliani, M.; Cappello, V.; Caracciolo, G.; Marchetti, P.; Piston, D.W.; Cardarelli, F. Insulin Secretory Granules Labelled with Phogrin-Fluorescent Proteins Show Alterations in Size, Mobility and Responsiveness to Glucose Stimulation in Living β-Cells. Sci. Rep. 2019, 9, 2890. [Google Scholar] [CrossRef]
- Durso, W.; Martins, M.; Marchetti, L.; Cremisi, F.; Luin, S.; Cardarelli, F. Lysosome Dynamic Properties during Neuronal Stem Cell Differentiation Studied by Spatiotemporal Fluctuation Spectroscopy and Organelle Tracking. Int. J. Mol. Sci. 2020, 21, 3397. [Google Scholar] [CrossRef]
- Durso, W.; D’Autilia, F.; Amodeo, R.; Marchetti, L.; Cardarelli, F. Probing Labeling-Induced Lysosome Alterations in Living Cells by Imaging-Derived Mean Squared Displacement Analysis. Biochem. Biophys. Res. Commun. 2018, 503, 2704–2709. [Google Scholar] [CrossRef]
- Tesi, M.; Bugliani, M.; Ferri, G.; Suleiman, M.; De Luca, C.; Bosi, E.; Masini, M.; De Tata, V.; Gysemans, C.; Cardarelli, F.; et al. Pro-Inflammatory Cytokines Induce Insulin and Glucagon Double Positive Human Islet Cells That Are Resistant to Apoptosis. Biomolecules 2021, 11, 320. [Google Scholar] [CrossRef]
- Marselli, L.; Piron, A.; Suleiman, M.; Colli, M.L.; Yi, X.; Khamis, A.; Carrat, G.R.; Rutter, G.A.; Bugliani, M.; Giusti, L.; et al. Persistent or Transient Human β Cell Dysfunction Induced by Metabolic Stress: Specific Signatures and Shared Gene Expression with Type 2 Diabetes. Cell Rep. 2020, 33, 108466. [Google Scholar] [CrossRef]
- Cnop, M.; Hannaert, J.C.; Hoorens, A.; Eizirik, D.L.; Pipeleers, D.G. Inverse Relationship Between Cytotoxicity of Free Fatty Acids in Pancreatic Islet Cells and Cellular Triglyceride Accumulation. Diabetes 2001, 50, 1771–1777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marselli, L.; Suleiman, M.; Masini, M.; Campani, D.; Bugliani, M.; Syed, F.; Martino, L.; Focosi, D.; Scatena, F.; Olimpico, F.; et al. Are we overestimating the loss of beta cells in type 2 diabetes? Diabetologia 2014, 57, 362–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosengren, A.H.; Braun, M.; Mahdi, T.; Andersson, S.A.; Travers, M.E.; Shigeto, M.; Zhang, E.; Almgren, P.; Ladenvall, C.; Axelsson, A.S.; et al. Reduced Insulin Exocytosis in Human Pancreatic β-Cells with Gene Variants Linked to Type 2 Diabetes. Diabetes 2012, 61, 1726–1733. [Google Scholar] [CrossRef] [Green Version]
- Hoboth, P.; Müller, A.; Ivanova, A.; Mziaut, H.; Dehghany, J.; Sönmez, A.; Lachnit, M.; Meyer-Hermann, M.; Kalaidzidis, Y.; Solimena, M. Aged Insulin Granules Display Reduced Microtubule-Dependent Mobility and Are Disposed within Actin-Positive Multigranular Bodies. Proc. Natl. Acad. Sci. USA 2015, 112, E667–E676. [Google Scholar] [CrossRef] [Green Version]
- Mosesson, Y.; Mills, G.B.; Yarden, Y. Derailed Endocytosis: An Emerging Feature of Cancer. Nat. Rev. Cancer 2008, 8, 835–850. [Google Scholar] [CrossRef] [PubMed]
- Cunha, D.A.; Ladrière, L.; Ortis, F.; Igoillo-Esteve, M.; Gurzov, E.N.; Lupi, R.; Marchetti, P.; Eizirik, D.L.; Cnop, M. Glucagon-Like Peptide-1 Agonists Protect Pancreatic β-Cells From Lipotoxic Endoplasmic Reticulum Stress Through Upregulation of BiP and JunB. Diabetes 2009, 58, 2851–2862. [Google Scholar] [CrossRef] [Green Version]
- Wajchenberg, B.L. β-Cell Failure in Diabetes and Preservation by Clinical Treatment. Endocr. Rev. 2007, 28, 187–218. [Google Scholar] [CrossRef] [Green Version]
- Marrano, N.; Biondi, G.; Cignarelli, A.; Perrini, S.; Laviola, L.; Giorgino, F.; Natalicchio, A. Functional Loss of Pancreatic Islets in Type 2 Diabetes: How Can We Halt It? Metabolism 2020, 110, 154304. [Google Scholar] [CrossRef] [PubMed]
- Ly, L.D.; Xu, S.; Choi, S.-K.; Ha, C.-M.; Thoudam, T.; Cha, S.-K.; Wiederkehr, A.; Wollheim, C.B.; Lee, I.-K.; Park, K.-S. Oxidative Stress and Calcium Dysregulation by Palmitate in Type 2 Diabetes. Exp. Mol. Med. 2017, 49, e291. [Google Scholar] [CrossRef] [PubMed]
- Sargsyan, E.; Artemenko, K.; Manukyan, L.; Bergquist, J.; Bergsten, P. Oleate Protects Beta-Cells from the Toxic Effect of Palmitate by Activating pro-Survival Pathways of the ER Stress Response. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2016, 1861, 1151–1160. [Google Scholar] [CrossRef]
- Lee, Y.-S.; Jun, H.-S. Anti-Inflammatory Effects of GLP-1-Based Therapies beyond Glucose Control. Mediat. Inflamm. 2016, 2016, 3094642. [Google Scholar] [CrossRef] [Green Version]
- Di Rienzo, C.; Gratton, E.; Beltram, F.; Cardarelli, F. Spatiotemporal Fluctuation Analysis: A Powerful Tool for the Future Nanoscopy of Molecular Processes. Biophys. J. 2016, 111, 679–685. [Google Scholar] [CrossRef] [Green Version]
- Petrenko, V.; Gandasi, N.R.; Sage, D.; Tengholm, A.; Barg, S.; Dibner, C. In Pancreatic Islets from Type 2 Diabetes Patients, the Dampened Circadian Oscillators Lead to Reduced Insulin and Glucagon Exocytosis. Proc. Natl. Acad. Sci. USA 2020, 117, 2484–2495. [Google Scholar] [CrossRef] [Green Version]
- Pulimeno, P.; Mannic, T.; Sage, D.; Giovannoni, L.; Salmon, P.; Lemeille, S.; Giry-Laterriere, M.; Unser, M.; Bosco, D.; Bauer, C.; et al. Autonomous and Self-Sustained Circadian Oscillators Displayed in Human Islet Cells. Diabetologia 2013, 56, 497–507. [Google Scholar] [CrossRef] [Green Version]
- Ghazvini Zadeh, E.H.; Huang, Z.; Xia, J.; Li, D.; Davidson, H.W.; Li, W. ZIGIR, a Granule-Specific Zn2+ Indicator, Reveals Human Islet α Cell Heterogeneity. Cell Rep. 2020, 32, 107904. [Google Scholar] [CrossRef]
- Chen, S.; Huang, Z.; Kidd, H.; Kim, M.; Suh, E.H.; Xie, S.; Ghazvini Zadeh, E.H.; Xu, Y.; Sherry, A.D.; Scherer, P.E.; et al. In Vivo ZIMIR Imaging of Mouse Pancreatic Islet Cells Shows Oscillatory Insulin Secretion. Front. Endocrinol. 2021, 12, 613964. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Cell | Labelling | Size (nm) | Dm (μm2/s) × 10−3 | α | N | SCD * |
|---|---|---|---|---|---|---|
| INS-1 E | Syn-EGFP | 333 ± 44 | 2.1 ± 1.2 | 0.72 ± 0.14 | 48 | / |
| dHI | Syn-EGFP | 261 ±45 | 1.0 ± 0.8 | 0.72 ± 0.17 | 123 | 0.536 |
| Condition | [Glucose] | Size (nm) | Dm (μm2/s) × 10−3 | A | N |
|---|---|---|---|---|---|
| Control | Low | 263 ± 49 | 1.1 ± 0.4 | 0.70 ± 0.18 | 44 |
| High | 266 ± 52 | 2.0 ± 1.1 | 0.78 ± 0.13 | 36 | |
| Palm | Low | 275 ± 50 | 2.0 ± 1.3 | 0.72 ± 0.13 | 28 |
| High | 275 ± 49 | 2.0 ± 1.6 | 0.69 ± 0.16 | 35 | |
| Palm + Exe-4 | Low | 247 ± 32 | 1.2 ± 0.1 | 0.70 ± 0.14 | 29 |
| High | 256 ± 43 | 2.3 ± 1.0 | 0.78 ± 0.11 | 19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferri, G.; Tesi, M.; Pesce, L.; Bugliani, M.; Grano, F.; Occhipinti, M.; Suleiman, M.; De Luca, C.; Marselli, L.; Marchetti, P.; et al. Spatiotemporal Correlation Spectroscopy Reveals a Protective Effect of Peptide-Based GLP-1 Receptor Agonism against Lipotoxicity on Insulin Granule Dynamics in Primary Human β-Cells. Pharmaceutics 2021, 13, 1403. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13091403
Ferri G, Tesi M, Pesce L, Bugliani M, Grano F, Occhipinti M, Suleiman M, De Luca C, Marselli L, Marchetti P, et al. Spatiotemporal Correlation Spectroscopy Reveals a Protective Effect of Peptide-Based GLP-1 Receptor Agonism against Lipotoxicity on Insulin Granule Dynamics in Primary Human β-Cells. Pharmaceutics. 2021; 13(9):1403. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13091403
Chicago/Turabian StyleFerri, Gianmarco, Marta Tesi, Luca Pesce, Marco Bugliani, Francesca Grano, Margherita Occhipinti, Mara Suleiman, Carmela De Luca, Lorella Marselli, Piero Marchetti, and et al. 2021. "Spatiotemporal Correlation Spectroscopy Reveals a Protective Effect of Peptide-Based GLP-1 Receptor Agonism against Lipotoxicity on Insulin Granule Dynamics in Primary Human β-Cells" Pharmaceutics 13, no. 9: 1403. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13091403