
Impacts of NO2 Impurities on the Indigenous Microbial Community Structure and Diversity in CO2-Saline-Sandstone Interaction System

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.1.1. Water and Microbial Sample Collection
2.1.2. Microbial Scale-Up Culture
2.1.3. Sandstone Samples
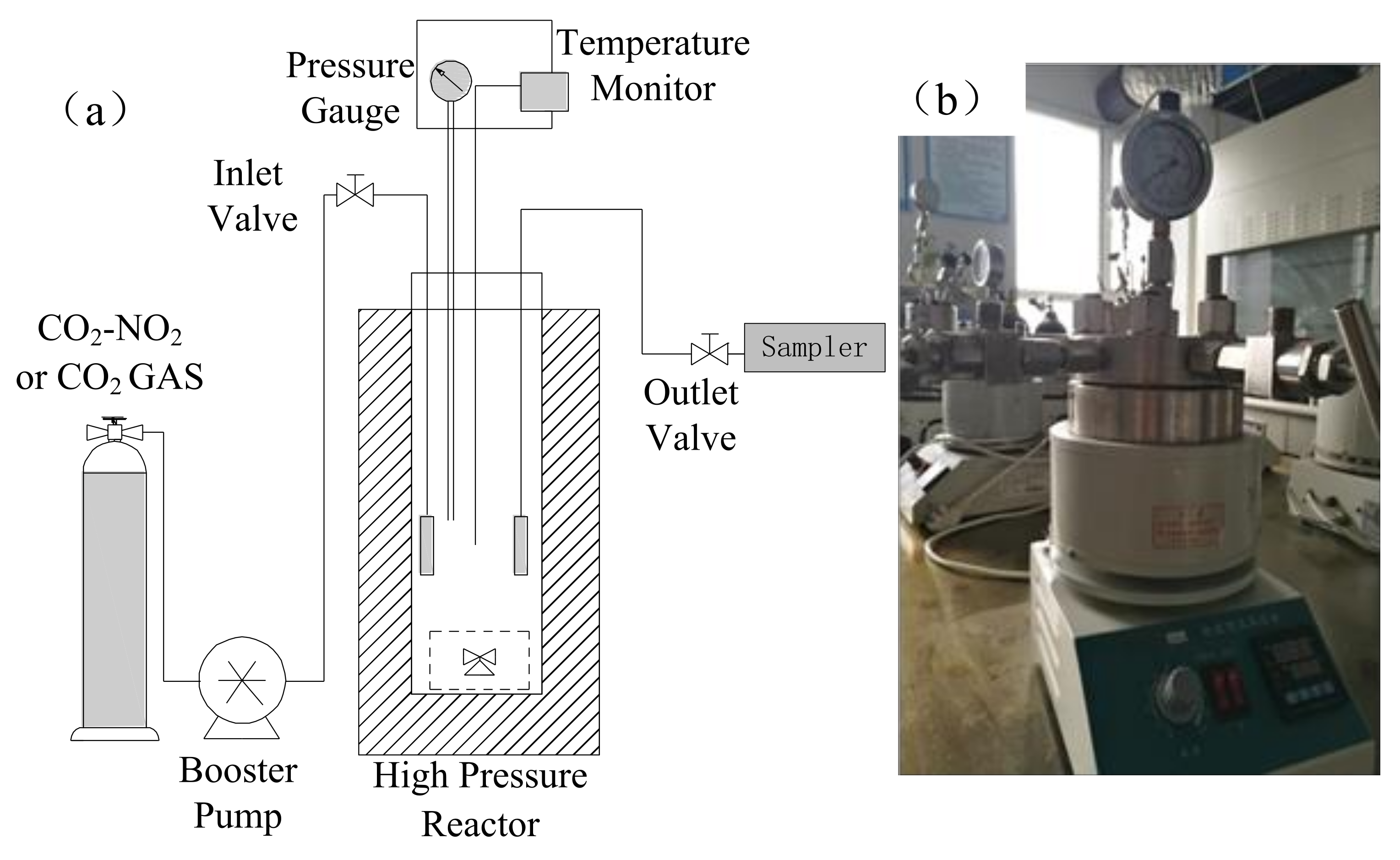
2.2. Experimental Set-Up and Approach
2.3. Microbial Analyses
2.3.1. DNA Extraction
2.3.2. 16S rRNA Gene Amplification and qPCR (Quantitative PCR)
2.3.3. 16S Gene Library Construction, Quantification and Sequence Processing
2.3.4. Statistical Analyses
3. Results and Discussion
3.1. Microbial Scale-Up Culture
3.2. Effect of NO2 Co—Injection on Biomass
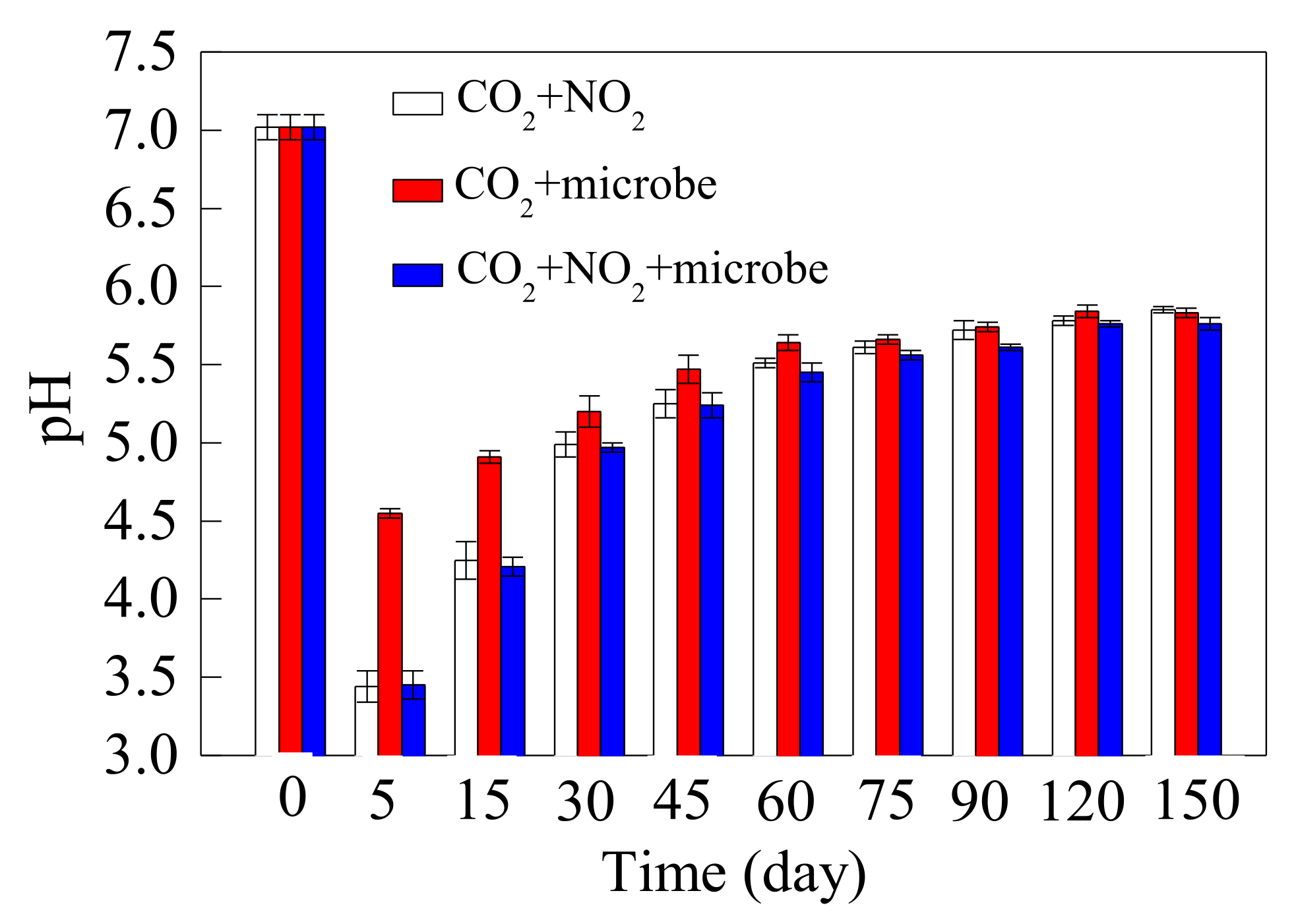
3.2.1. The Change of pH
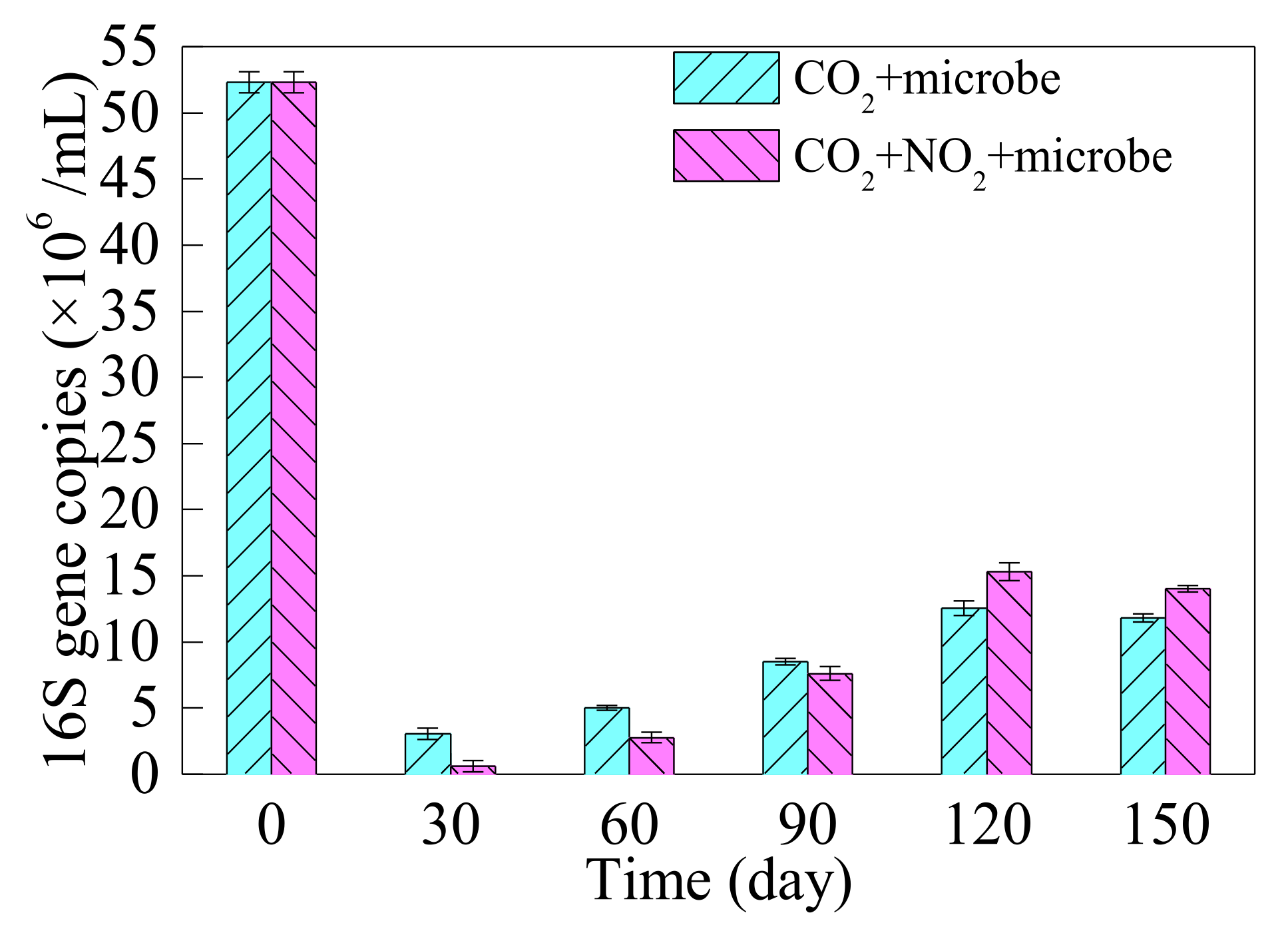
3.2.2. Biomass Change
3.3. Effects of NO2 Co—Injection on Community Structure
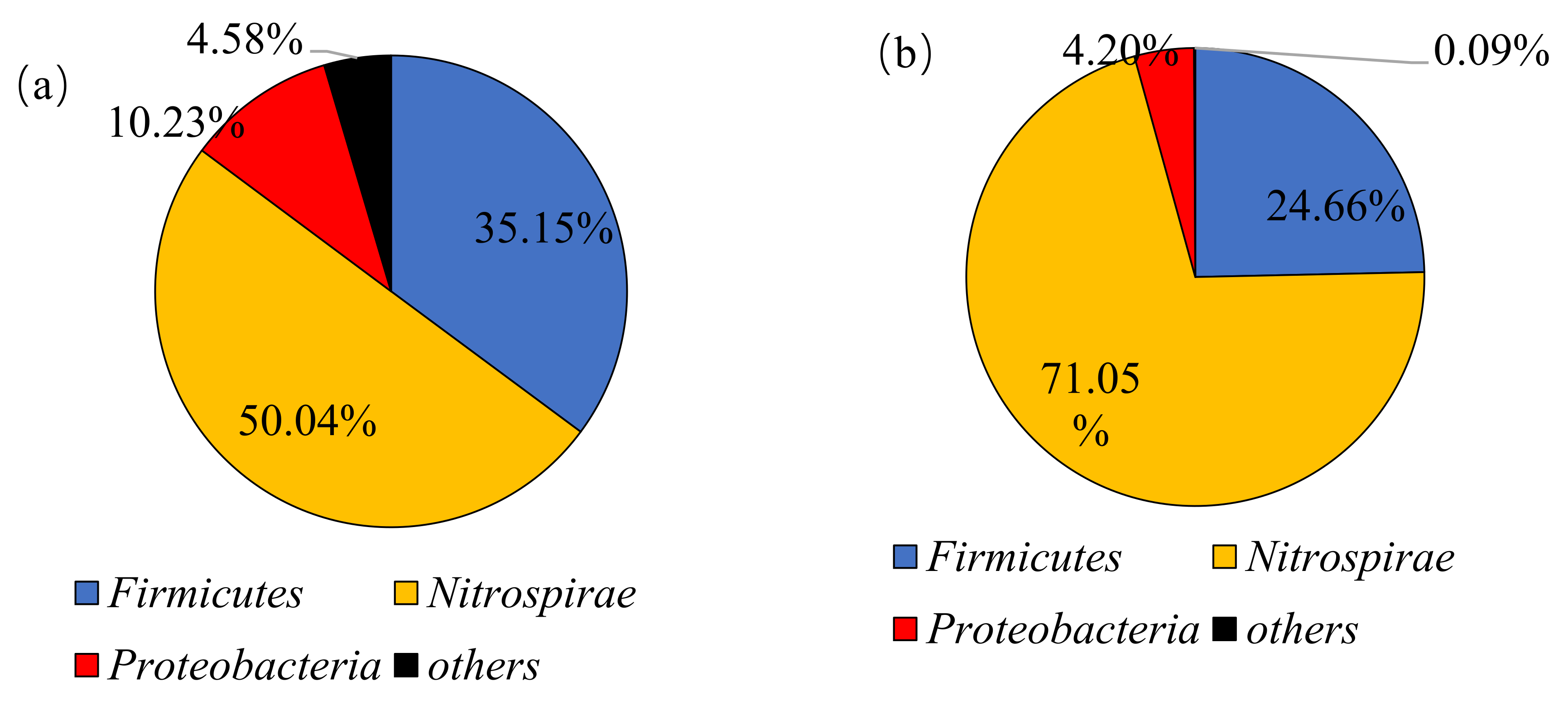
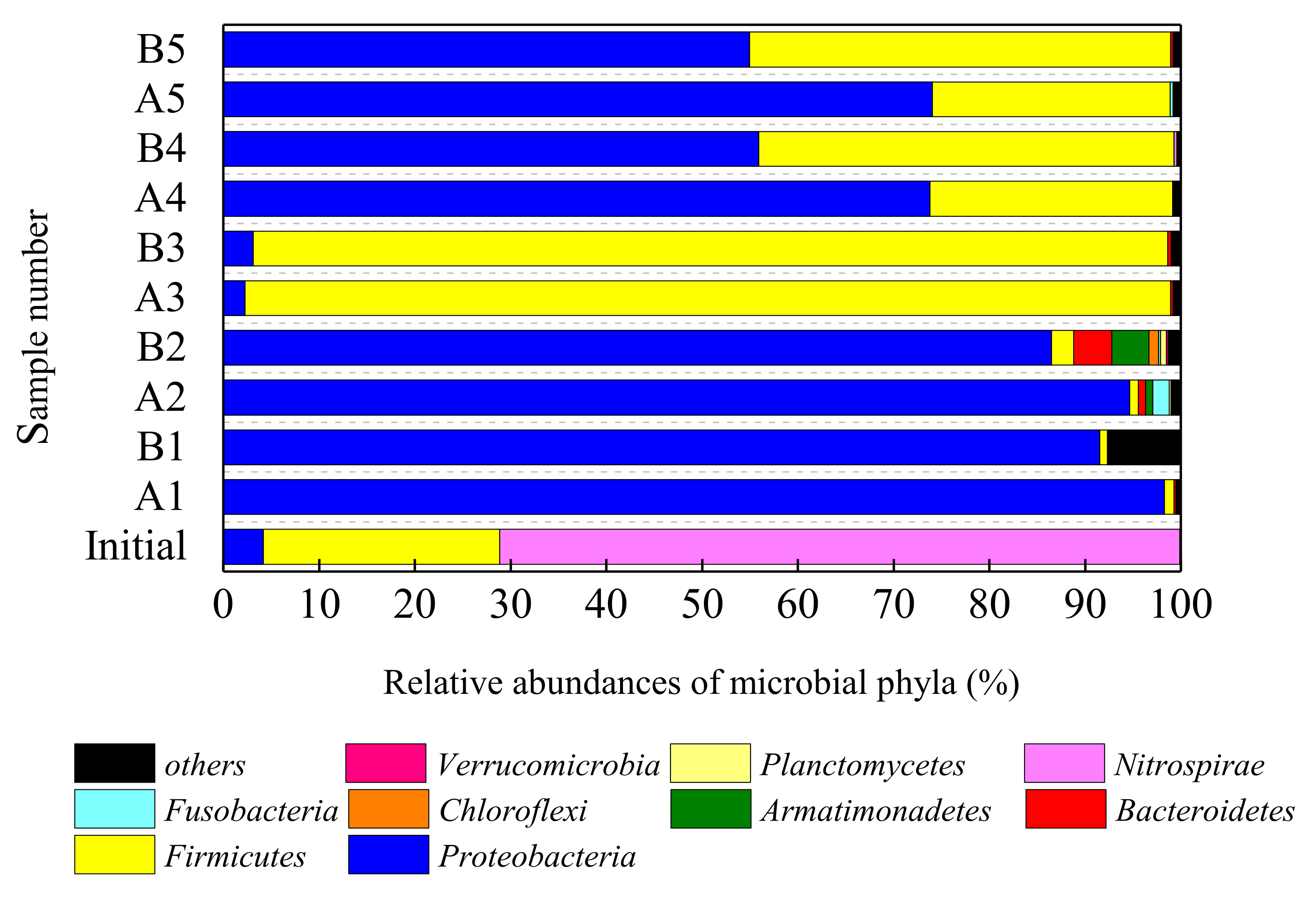
3.3.1. Changes of Phylum Classification Level
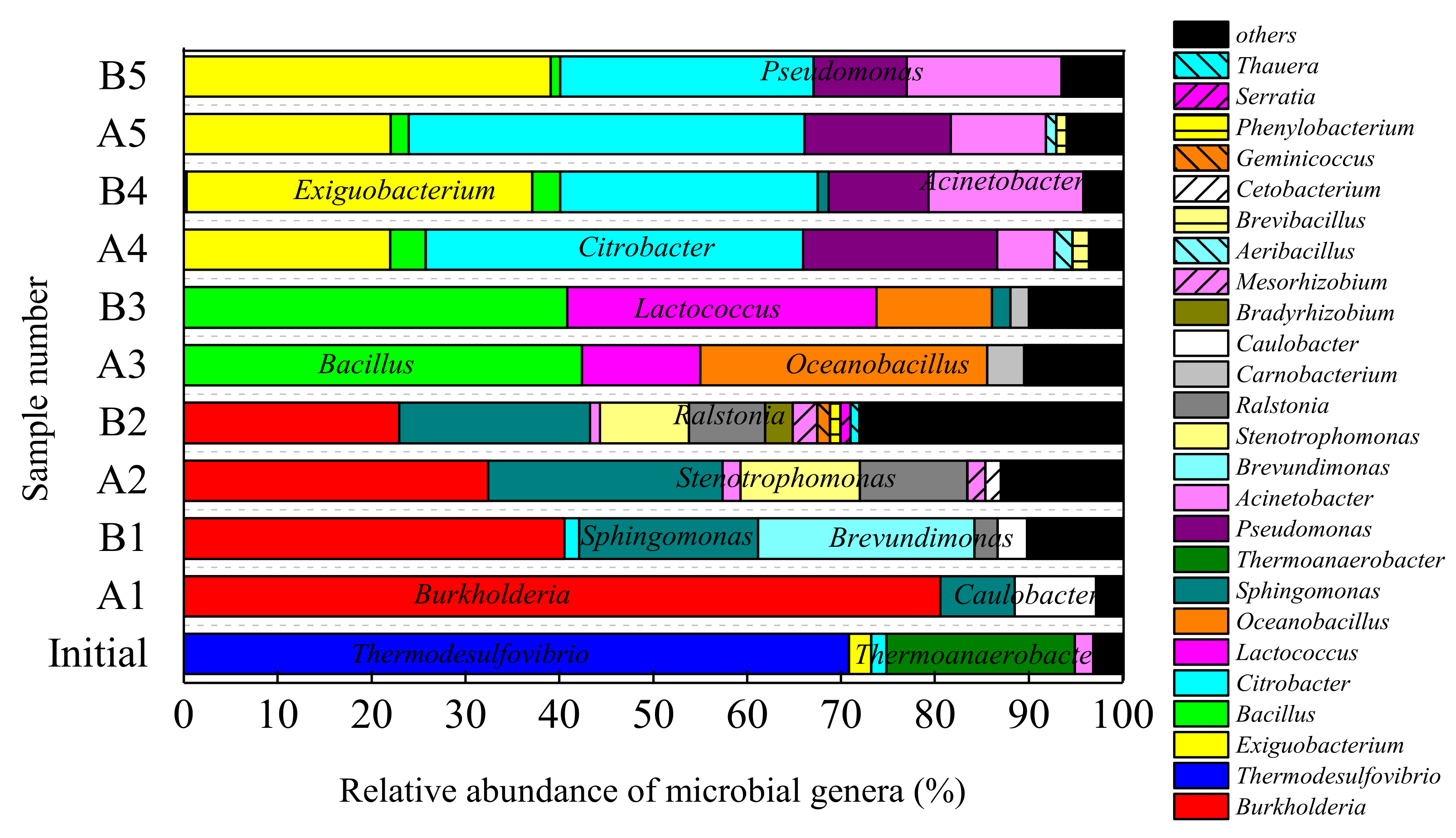
3.3.2. Microbial Community Changes at the Genus Level
3.4. Effects of NO2 Co—Injection on Biodiversity
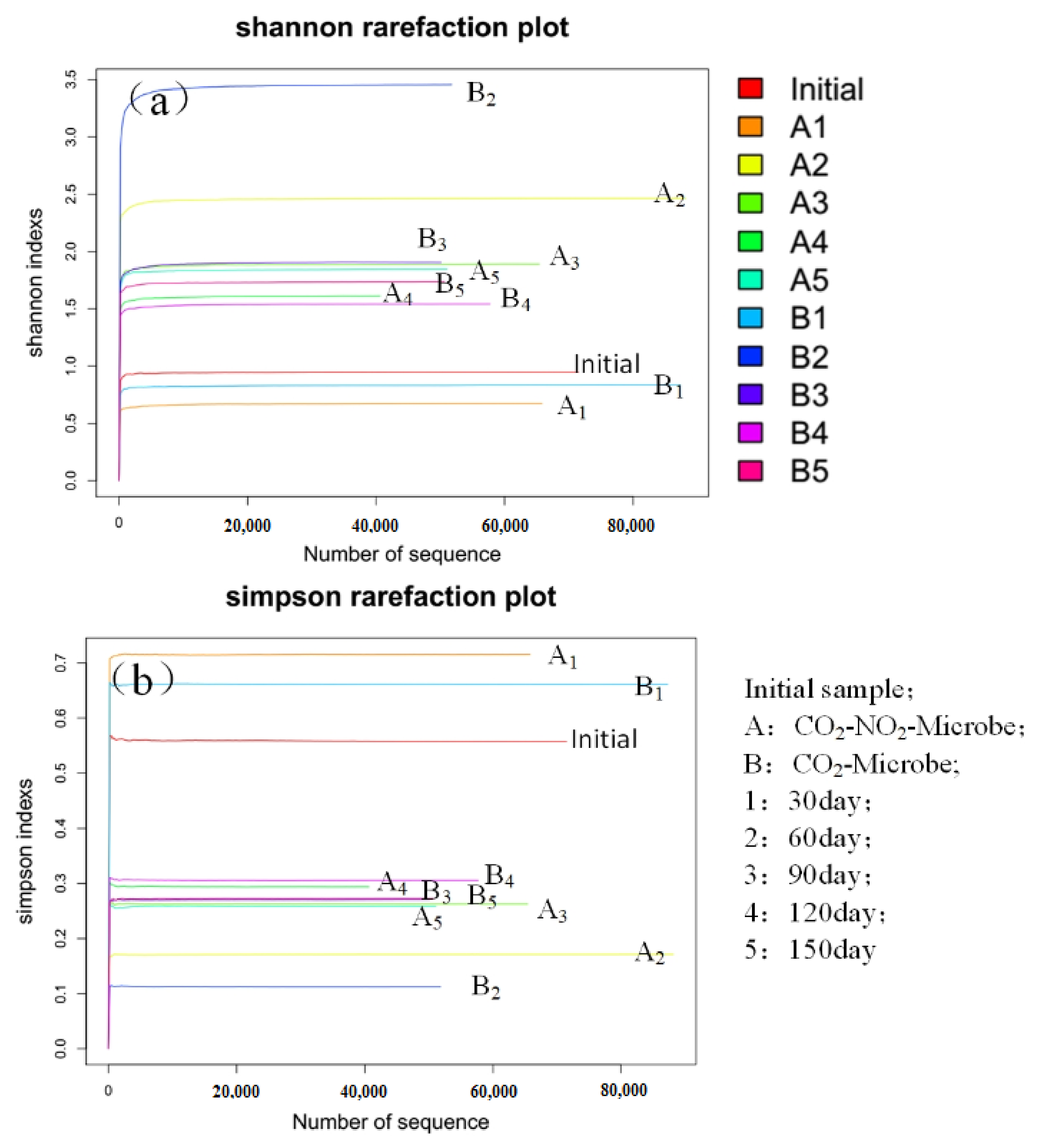
3.4.1. Alpha Diversity Analysis
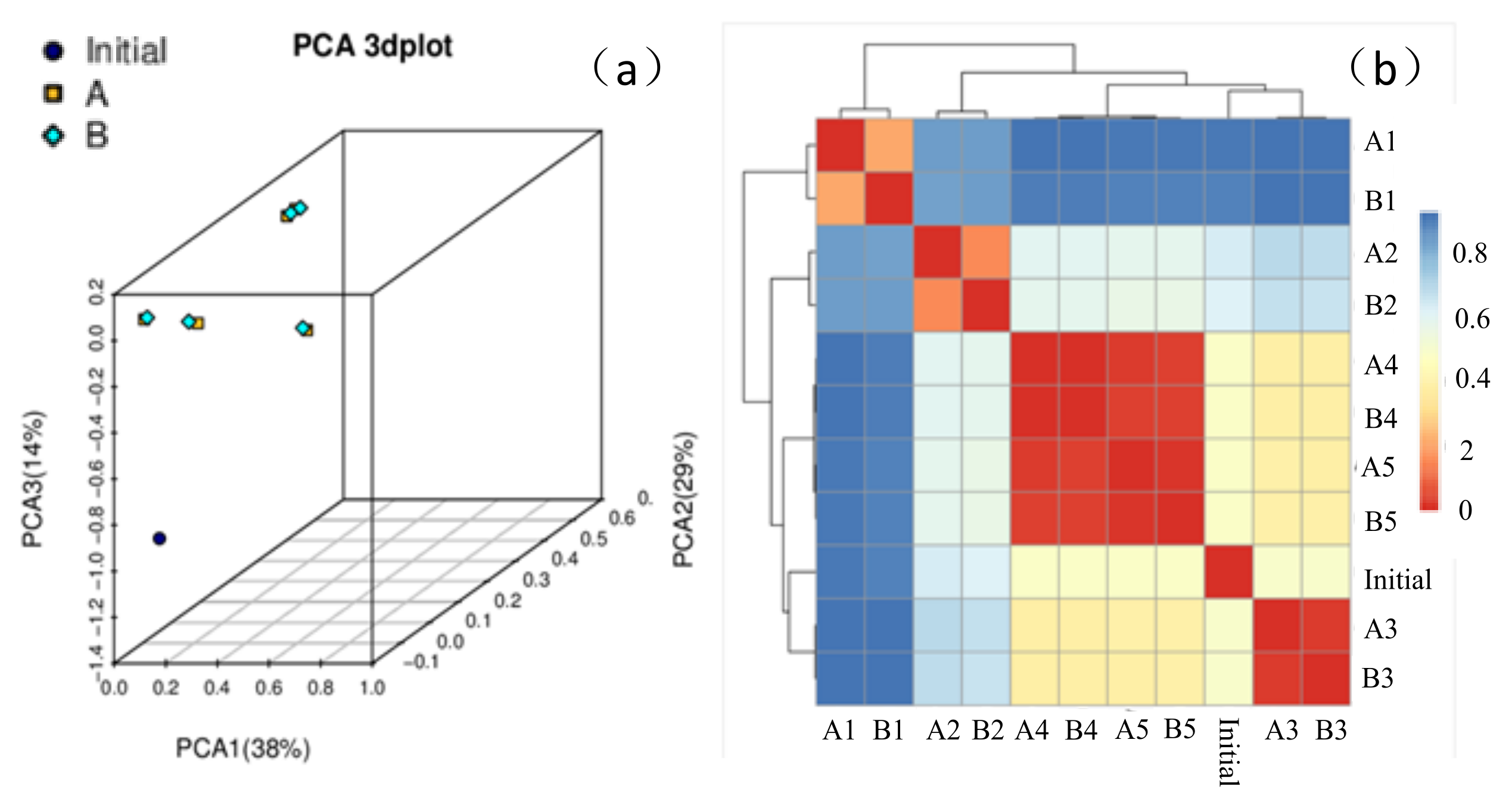
3.4.2. Multidimensional Analysis and Beta Diversity Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- NOAA. 2021. Available online: https://www.esrl.noaa.gov/gmd/ccgg/trends/global.html (accessed on 11 May 2021).
- Haszeldine, R.S. Carbon Capture and Storage: How Green Can Black Be? Science 2009, 325, 1647–1652. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.; Chung, Y.; Lee, J.; Seo, H. Knowledge spillover efficiency of carbon capture, utilization, and storage technology: A comparison among countries. J. Clean. Prod. 2020, 246, 119003. [Google Scholar] [CrossRef]
- Sokama-Neuyam, Y.A.; Adu-Boahene, F.; Boakye, P.; Aggrey, W.N.; Ursin, J.R. Theoretical modeling of the effect of temperature on CO2 injectivity in deep saline formations. Greenh. Gases Sci. Technol. 2020, 10, 4–14. [Google Scholar] [CrossRef]
- Trémosa, J.; Castillo, C.; Vong, C.Q.; Kervévan, C.; Lassin, A.; Audigane, P. Long-term assessment of geochemical reactivity of CO2 storage in highly saline aquifers: Application to Ketzin, In Salah and Snøhvit storage sites. Int. J. Greenh. Gas Control 2014, 20, 2–26. [Google Scholar] [CrossRef] [Green Version]
- Rendel, P.M.; Wolff-Boenish, D.; Gavrielli, I.; Ganor, J. Planned Experimental Studies into the Nucleation and Precipitation Kinetics of Sulfate-bearing Minerals under Conditions Relevant to CO2 Geological Storage. Energy Procedia 2013, 37, 5494–5500. [Google Scholar] [CrossRef] [Green Version]
- Fischer, S.; Liebscher, A.; De Lucia, M.; Hecht, L.; Ketzin Team. Reactivity of sandstone and siltstone samples from the Ketzin pilot CO2 storage site-Laboratory experiments and reactive geochemical modeling. Env. Earth Sci. 2013, 70, 3687–3708. [Google Scholar] [CrossRef]
- De Silva, G.; Ranjith, P.; Perera, M. Geochemical aspects of CO2 sequestration in deep saline aquifers: A review. Fuel 2015, 155, 128–143. [Google Scholar] [CrossRef]
- Li, D.; Jiang, X. Numerical investigation of convective mixing in impure CO2 geological storage into deep saline aquifers. Int. J. Greenh. Gas Control 2020, 96, 103015. [Google Scholar] [CrossRef]
- Talman, S. Subsurface geochemical fate and effects of impurities contained in a CO2 stream injected into a deep saline aquifer: What is known. Int. J. Greenh. Gas Control 2015, 40, 267–291. [Google Scholar] [CrossRef]
- Aminu, M.; Ali, N.S.; Vasilije, M. CO2-brine-rock interactions: The effect of impurities on grain size distribution and reservoir permeability. Int. J. Greenh. Gas Control 2018, 78, 168–176. [Google Scholar] [CrossRef]
- Hedayati, M.; Wigston, A.; Wolf, J.L.; Rebscher, D.; Niemi, A. Impacts of SO2 gas impurity within a CO2 stream on reservoir rock of a CCS pilot site: Experimental and modelling approach. Int. J. Greenh. Gas Control 2018, 70, 32–44. [Google Scholar] [CrossRef]
- Nicot, J.-P.; Solano, S.; Lu, J.; Mickler, P.; Romanak, K.; Yang, C.; Zhang, X. Potential Subsurface Impacts of CO2 Stream Impurities on Geologic Carbon Storage. Energy Procedia 2013, 37, 4552–4559. [Google Scholar] [CrossRef] [Green Version]
- Uroz, S.; Calvaruso, C.; Turpault, M.-P.; Frey-Klett, P. Mineral weathering by bacteria: Ecology, actors and mechanisms. Trends Microbiol. 2009, 17, 378–387. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Bachani, P.; Jain, D.; Patidar, S.K.; Mishra, S. Extraction of potassium from K-feldspar through potassium solubilization in the halophilic Acinetobacter soli (MTCC 5918) isolated from the experimental salt farm. Int. J. Miner. Process. 2016, 152, 53–57. [Google Scholar] [CrossRef]
- Mitchell, A.C.; Dideriksen, K.; Spangler, L.H.; Cunningham, A.B.; Gerlach, R. Microbially Enhanced Carbon Capture and Storage by Mineral-Trapping and Solubility-Trapping. Env. Sci. Technol. 2010, 44, 5270–5276. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Lee, D.-J.; Wang, A.; Ren, N.; Su, A.; Lai, J.-Y. Isolation of Fe(III)-reducing bacterium, Citrobacter sp. LAR-1, for startup of microbial fuel cell. Int. J. Hydrog. Energy 2016, 41, 4498–4503. [Google Scholar] [CrossRef]
- Li, C.; Zhong, S.; Zhang, F.; Wang, Z.; Jiang, F.; Wan, Y. Response of microbial communities to supercritical CO2 and biogeochemical influences on microbially mediated CO2-saline-sandstone interactions. Chem. Geol. 2017, 473, 1–9. [Google Scholar] [CrossRef]
- Zhang, F.; Song, Y.; Li, C.; Zhang, S.; Lyu, C.; Fan, K. The Impact of Indigenous Microorganisms on the Mineral Corrosion and Mineral Trapping in the SO2 Co-injected CO2-Saline-Sandstone Interaction. Geomicrobiol. J. 2019, 36, 110–122. [Google Scholar] [CrossRef]
- Li, W.; Wang, L.-Y.; Duan, R.-Y.; Liu, J.-F.; Gu, J.-D.; Mu, B.-Z. Microbial community characteristics of petroleum reservoir production water amended with n-alkanes and incubated under nitrate-, sulfate-reducing and methanogenic conditions. Int. Biodeterior. Biodegrad. 2012, 69, 87–96. [Google Scholar] [CrossRef]
- Sanz, J.L.; Köchling, T. Molecular biology techniques used in wastewater treatment: An overview. Process. Biochem. 2007, 42, 119–133. [Google Scholar] [CrossRef]
- Tantikachornkiat, M.; Sakakibara, S.; Neuner, M.; Durall, D.M. The use of propidium monoazide in conjunction with qPCR and Illumina sequencing to identify and quantify live yeasts and bacteria. Int. J. Food Microbiol. 2016, 234, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, C.; Xu, J.; Wu, X.; Dong, F.; Liu, X.; Tian, C.; Zheng, Y. Effects of hexaconazole application on soil microbes community and nitrogen transformations in paddy soils. Sci. Total Environ. 2017, 609, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Cheng, C.; He, L.; Huang, Z.; Sheng, X. Characterization of Depth-Related Changes in Bacterial Communities Involved in Mineral Weathering Along a Mineral-Rich Soil Profile. Geomicrobiol. J. 2014, 31, 431–444. [Google Scholar] [CrossRef]
- Li, X.; Zhao, L.; Adam, M. Biodegradation of marine crude oil pollution using a salt-tolerant bacterial consortium isolated from Bohai Bay, China. Mar. Pollut. Bull. 2016, 105, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Wang, S.; Xue, L.; Liu, D.; Zhu, X.; Ma, J.; Huang, C.; Yu, H.; Jiang, S. Research of the Impact of Elevated CO2 on Soil Microbial Diversity. Energy Procedia 2017, 114, 3070–3076. [Google Scholar] [CrossRef]
- Lepinay, C.; Mihajlovski, A.; Seyer, D.; Touron, S.; Bousta, F.; Di Martino, P. Biofilm communities survey at the areas of salt crystallization on the walls of a decorated shelter listed at UNESCO World cultural Heritage. Int. Biodeterior. Biodegrad. 2017, 122, 116–127. [Google Scholar] [CrossRef]
- Hirsch, P. Microbial life at extremely low nutrient levels. Adv. Space Res. 1986, 6, 287–298. [Google Scholar] [CrossRef]
- Rusznyák, A.; Akob, D.; Nietzsche, S.; Eusterhues, K.; Totsche, K.U.; Neu, T.R.; Frosch, T.; Popp, J.; Keiner, R.; Geletneky, J.; et al. Calcite Biomineralization by Bacterial Isolates from the Recently Discovered Pristine Karstic Herrenberg Cave. Appl. Environ. Microbiol. 2011, 78, 1157–1167. [Google Scholar] [CrossRef] [Green Version]
- Gao, P.; Sun, X.; Xiao, E.; Xu, Z.; Li, B.; Sun, W. Characterization of iron-metabolizing communities in soils contaminated by acid mine drainage from an abandoned coal mine in Southwest China. Environ. Sci. Pollut. Res. 2019, 26, 9585–9598. [Google Scholar] [CrossRef]
- Wang, Q.; Xie, Q.; He, L.; Sheng, X. The abundance and mineral-weathering effectiveness of Bacillus strains in the altered rocks and the soil. J. Basic Microbiol. 2018, 58, 770–781. [Google Scholar] [CrossRef]
- Jadhav, V.V.; Yadav, A.; Shouche, Y.S.; Aphale, S.; Moghe, A.; Pillai, S.; Arora, A.; Bhadekar, R.K. Studies on biosurfactant from Oceanobacillus sp. BRI 10 isolated from Antarctic sea water. Desalination 2013, 318, 64–71. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Baek, K.H. Physiological and biochemical perspectives of non-salt tolerant plants during bacterial interaction against soil salinity. Plant Physiol. Biochem. 2017, 116, 116–126. [Google Scholar] [CrossRef]
- Yanmis, D.; Orhan, F.; Gulluce, M.; Sahin, F. Biotechnological magnesite enrichment using a carbonate dissolving microorganism, Lactococcus sp. Int. J. Miner. Process. 2015, 144, 21–25. [Google Scholar] [CrossRef]
- Sofos, J.; Pierson, M.; Blocher, J.; Busta, F. Mode of action of sorbic acid on bacterial cells and spores. Int. J. Food Microbiol. 1986, 3, 1–17. [Google Scholar] [CrossRef]
- Ko, M.-S.; Cho, K.; Jeong, D.; Lee, S. Identification of the microbes mediating Fe reduction in a deep saline aquifer and their influence during managed aquifer recharge. Sci. Total Environ. 2016, 545–546, 486–492. [Google Scholar] [CrossRef]
- Govorukha, V.M.; Tashyrev, O.B. The regularities of iron compounds transformation by Citrobacter freundii MI-31.1/1. Mikrobiolohichnyi Zhurnal 2016, 78, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Sen, S.K.; Jana, A.; Bandyopadhyay, P.; Das Mohapatra, P.K.; Raut, S. Thermostable amylase production from hot spring isolate Exiguobacterium sp: A promising agent for natural detergents. Sustain. Chem. Pharm. 2016, 3, 59–68. [Google Scholar] [CrossRef]
- Zhuzheng, C. Isolation and Identification of a Strain of Acid Tolerant Denitrifying Bacteria and Its Denitrification Characteristics; East China University of Technology: Nanchang, China, 2017. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | CO2 | N2 | O2 | H2 | CH4 | |
|---|---|---|---|---|---|---|
| Mole fraction (%) | Minimum | 75 | 0.02 | 0.04 | 0.06 | 0.7 |
| Maximum | 99 | 10 | 5 | 4 | 4 | |
| Components | Ar | SO2 | H2S + COS | NOx | CO | |
| Mole fraction (%) | Minimum | 0.005 | <0.0001 | 0.01 | 0.0002 | 0.0001 |
| Maximum | 3.5 | 1.5 | 1.5 | 0.3 | 0.2 | |
| Properties | ρ (mg/L) | Properties | ρ (mg/L) |
|---|---|---|---|
| Ca2+ | 1067.33 | SO42− | 46.28 |
| Mg2+ | 4.47 | NO3− | 72.67 |
| K+ | 76.42 | Cl− | 5339.34 |
| Na+ | 2356 | HCO3− | 520.26 |
| Total Iron | 27.47 | TOC | 78 |
| pH | 7.02 |
| Properties | Before Cultivation | Abundance (%) | After Cultivation | Abundance (%) |
|---|---|---|---|---|
| The abundance of major genera | Thermodesulfovibrio | 49.36 | Thermodesulfovibrio | 70.84 |
| Thermoanaerobacter | 22.12 | Thermoanaerobacter | 20.04 | |
| Bacillus | 3.84 | Exiguobacterium | 2.37 | |
| Moorella | 3.55 | Acinetobacter | 1.96 | |
| Pseudomonas | 3.15 | Citrobacter | 1.64 | |
| Exiguobacterium | 2.51 | Clostridium III | 0.58 | |
| Streptococcus | 2.23 | Pseudomonas | 0.57 | |
| Citrobacter | 2.05 | Desulfitobacterium | 0.40 | |
| Brevibacillus | 2.05 | Brevibacillus | 0.33 | |
| Acinetobacter | 1.63 | Moorella | 0.26 | |
| Clostridium III | 1.56 | Bacillus | 0.04 | |
| Enterococcus | 1.02 | Enterococcus | 0.02 | |
| Others | 4.93 | Others | 0.95 | |
| 16S rRNA gene copies | 2.67 × 105 ± 0.73 × 105 copies/mL | 8.89 × 107 ± 0.80 × 107 copies/mL | ||
| Time (Days) | Phylum—Genera | Abundance % | Identity (Similarity %): Characteristics in Refs. | |
|---|---|---|---|---|
| CO2-NO2 | CO2 | |||
| 30-day | Proteobacteria-Burkholderia | 80.60 | 40.53 | B. spp. (98%): acid-tolerant, dissolution [25] |
| Proteobacteria-Brevundimonas | <1.0 | 23.01 | B sp. (97%): acid and salt-tolerant [26] | |
| Proteobacteria-sphingomonas | 7.91 | 19.06 | S.alaskensis (99%): acid and salt-tolerant [27,28] | |
| Proteobacteria-Caulobacter | 8.69 | 3.10 | C. spp. (97%): salt-tolerant [29] | |
| 60-day | Proteobacteria-Burkholderia | 32.41 | 22.97 | B. spp. (98%): acid-tolerant, dissolution [25] |
| Proteobacteria-phingomonas | 24.96 | 20.32 | S.alaskensis (99%): acid and salt-tolerant [27,28] | |
| Proteobacteria-Stenotrophomonas | 12.71 | 9.48 | S. maltophilia (98%): acid-tolerant [30,31] | |
| Time | Phylum—Genera | Abundance % | Identity (Similarity %): Characteristics in Refs. | |
|---|---|---|---|---|
| CO2-NO2 | CO2 | |||
| 90-day | Firmicutes-Bacillus | 42.40 | 40.85 | B. cereus (99%): Resistant to acid, dissolution [32] |
| Firmicutes-Oceanobacillus | 30.55 | 12.30 | O. sp. (97%): Resistant to severe stress and acid [33,34] | |
| Firmicutes-Lactococcus | 12.64 | 32.95 | L. sp. (99%): acid-producing [35] | |
| Time | Phylum—Genera | Abundances % | Identity (Similarity %): Characteristics in Refs. | |
|---|---|---|---|---|
| CO2-NO2 | CO2 | |||
| 120-day | Proteobacteria-Citrobacter | 40.17 | 27.43 | C. freundii (99%): Fe(III) reduction [37,38] |
| Firmicutes-Exiguobacterium | 21.97 | 36.78 | E. sp. (99%): Dissolution [39] | |
| Proteobacteria-Pseudomonas | 20.66 | 10.64 | P. aeruginosa (99%): Denitrification [40] | |
| Firmicutes-Acinetobacter | 6.13 | 16.49 | A. soli (97%): Dissolution [15] | |
| Firmicutes-Bacillus | 3.82 | 3.00 | B. cereus (98%): Dissolution [32] | |
| 150-day | Proteobacteria-Citrobacter | 42.16 | 26.97 | C. freundii (99%): Fe(III) reduction [37,38] |
| Firmicutes-Exiguobacterium | 22.04 | 39.08 | E. sp. (99%): Dissolution [39] | |
| Proteobacteria-Pseudomonas | 15.54 | 9.92 | P. aeruginosa (99%): Denitrification [40] | |
| Firmicutes-Acinetobacter | 10.15 | 16.50 | A.soli (97%): Dissolution [15] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Song, Y.; Zhu, Y.; Meng, Q.; Lyu, Y. Impacts of NO2 Impurities on the Indigenous Microbial Community Structure and Diversity in CO2-Saline-Sandstone Interaction System. Sustainability 2021, 13, 5916. https://0-doi-org.brum.beds.ac.uk/10.3390/su13115916
Li C, Song Y, Zhu Y, Meng Q, Lyu Y. Impacts of NO2 Impurities on the Indigenous Microbial Community Structure and Diversity in CO2-Saline-Sandstone Interaction System. Sustainability. 2021; 13(11):5916. https://0-doi-org.brum.beds.ac.uk/10.3390/su13115916
Chicago/Turabian StyleLi, Chenyang, Yunpeng Song, Yongjuan Zhu, Qingwei Meng, and Ying Lyu. 2021. "Impacts of NO2 Impurities on the Indigenous Microbial Community Structure and Diversity in CO2-Saline-Sandstone Interaction System" Sustainability 13, no. 11: 5916. https://0-doi-org.brum.beds.ac.uk/10.3390/su13115916