
Optimized High-Performance Liquid Chromatography Method for Determining Nine Cytokinins, Indole-3-acetic Acid and Abscisic Acid

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Instruments and Reagents
2.2. Plant Materials and Samples
2.3. Extraction and Purification
2.3.1. Sample Preparation
2.3.2. Optimization of HPLC-UV Conditions
2.3.3. Method Validation
3. Results and Discussions
3.1. Mobile Phase Elution Optimization
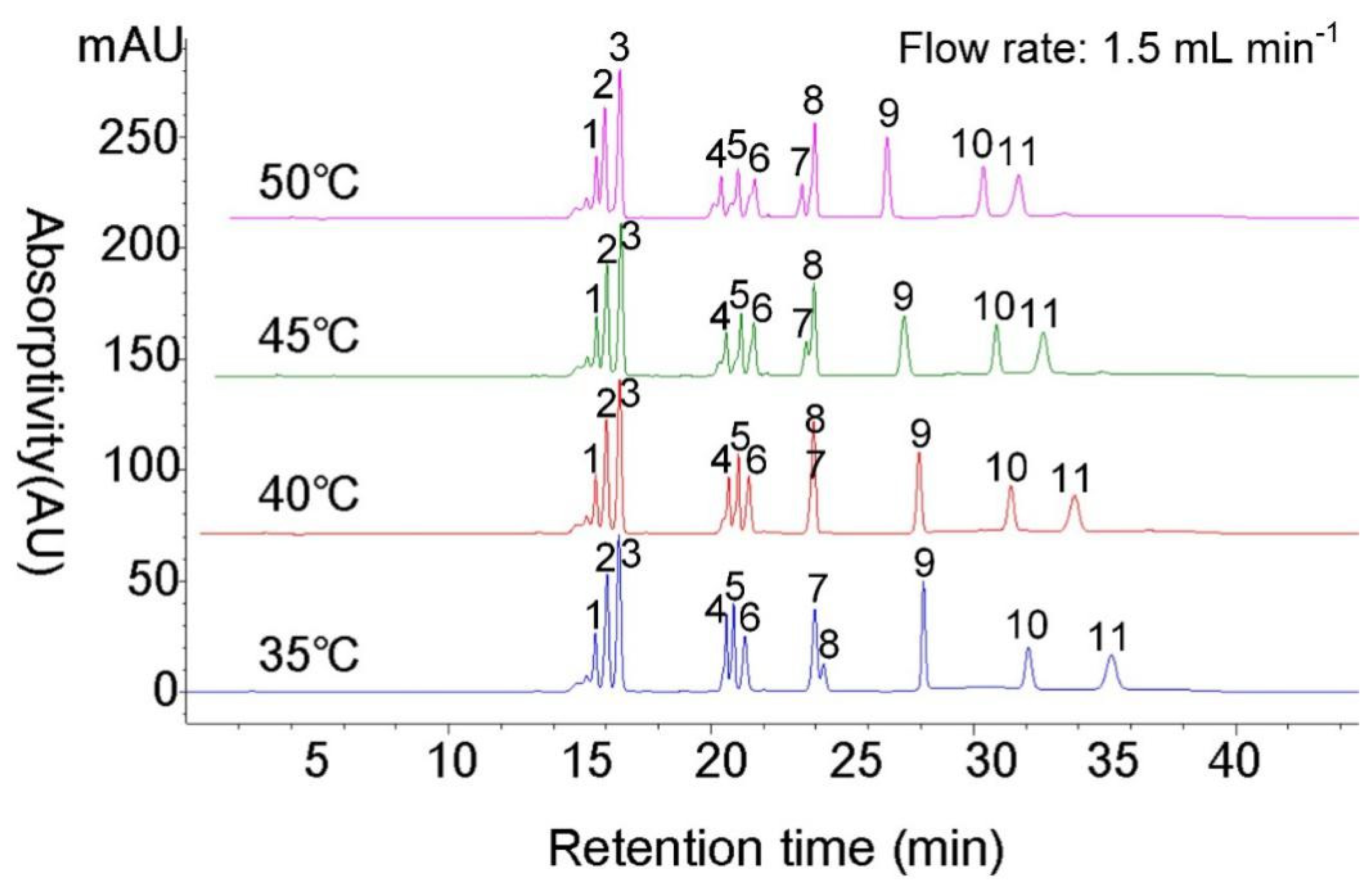
3.2. Optimizing the Column Temperature
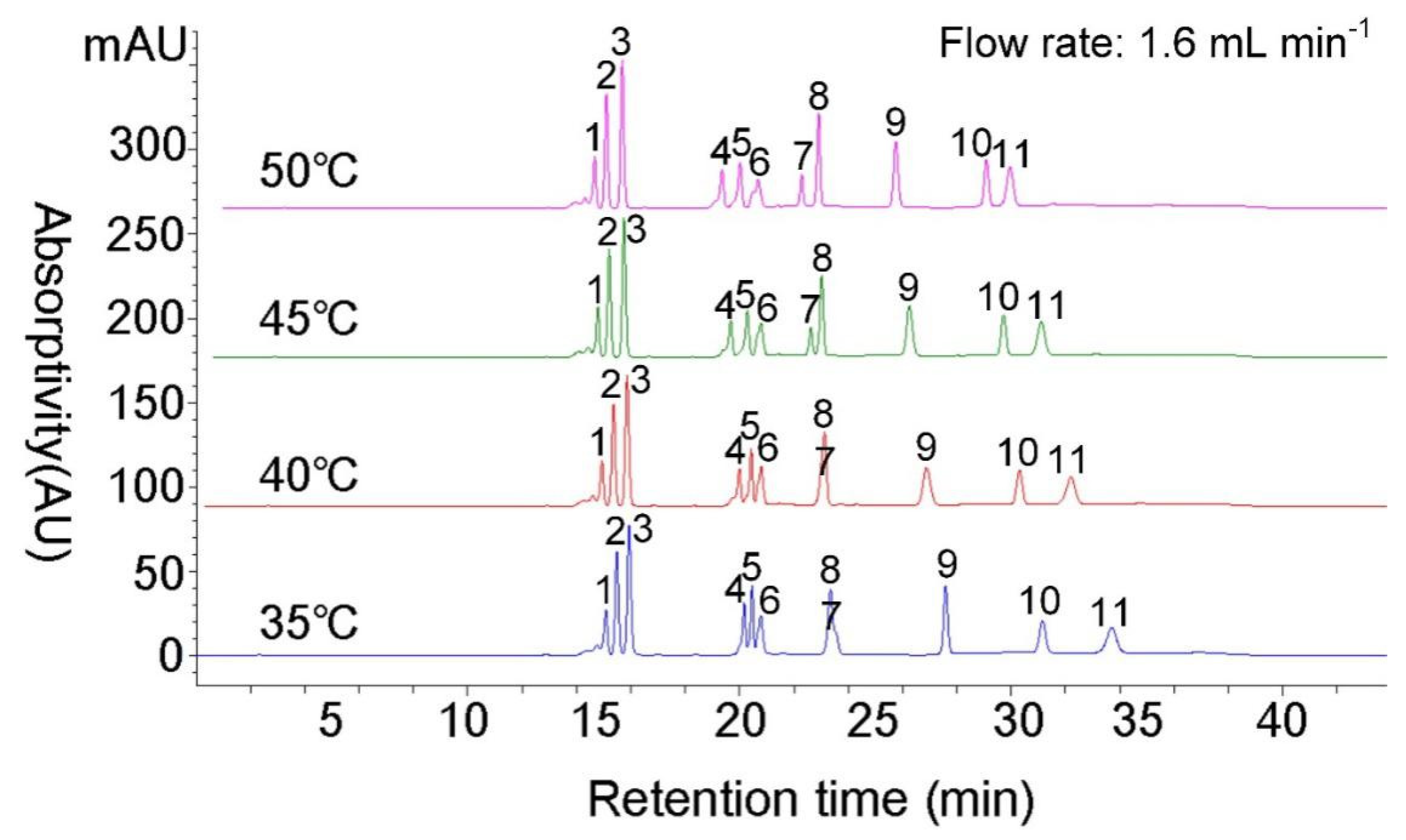
3.3. Flow Rate Optimization
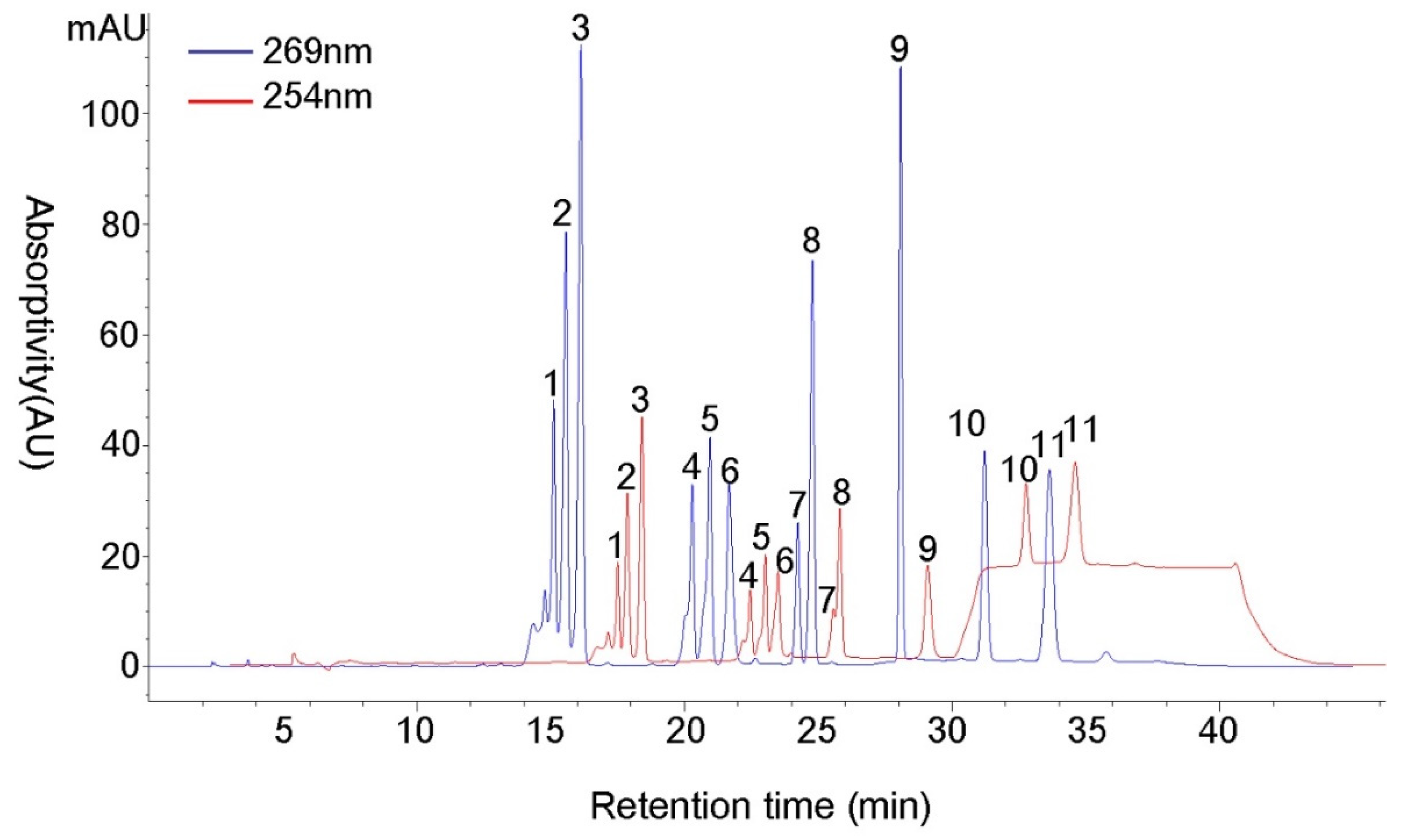
3.4. UV Conditions Optimization
3.5. Method Validation
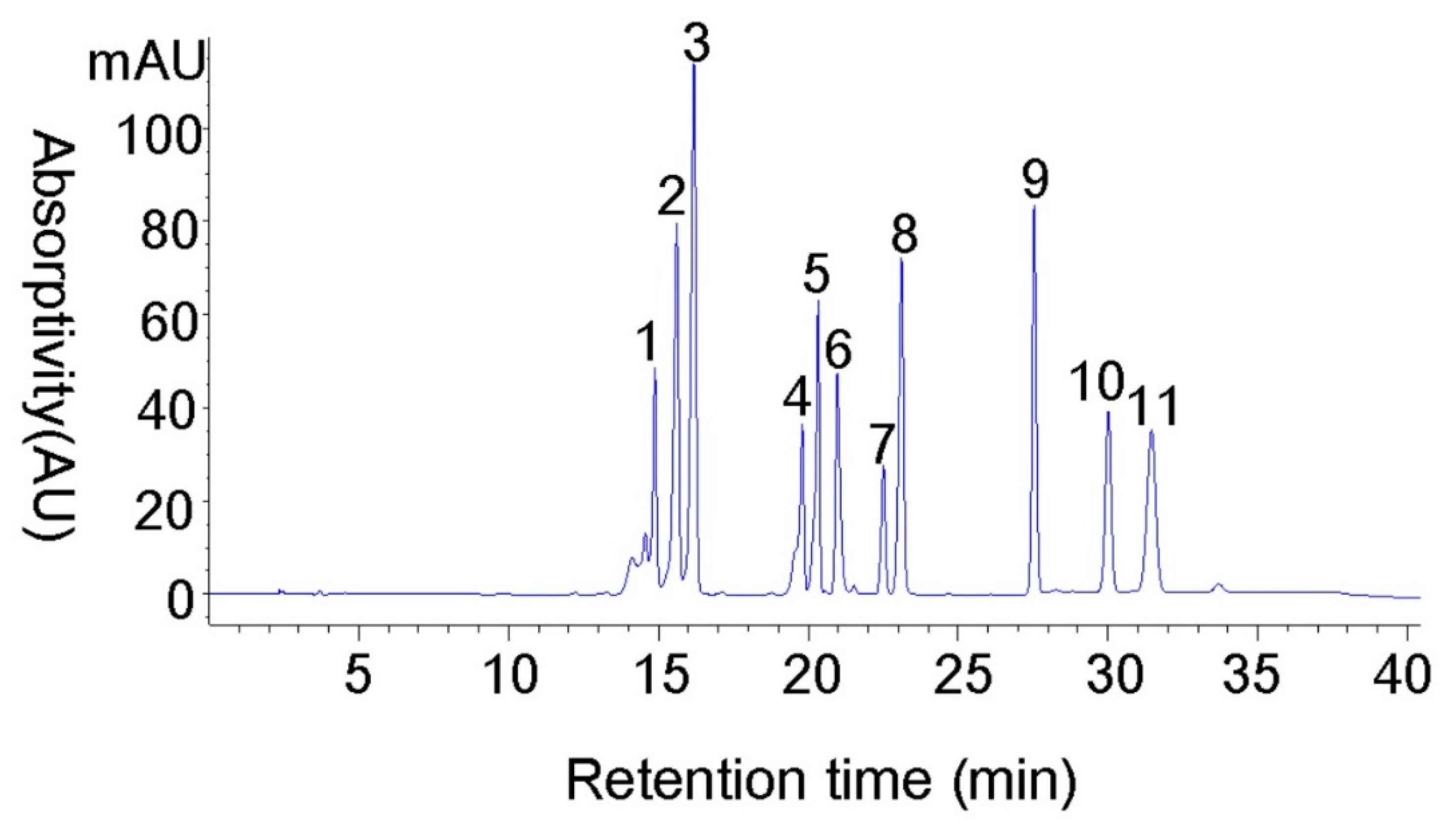
3.6. Sample Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sakakibara, H. Cytokinins: Activity, biosynthesis, and translocation. Annu. Rev. Plant Biol. 2006, 57, 431–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stirk, W.A.; Václavíková, K.; Novák, O.; Gajdošová, S.; Kotland, O.; Motyka, V.; Strnad, M.; Staden, J.V. Involvement of cis-Zeatin, dihydrozeatin, and aromatic cytokinins in germination and seedling establishment of maize, oats, and lucerne. J. Plant Growth Regul. 2012, 31, 392–405. [Google Scholar] [CrossRef]
- Wu, C.; Tang, S.; Li, G.; Wang, S.; Fahad, S.; Ding, Y. Roles of phytohormone changes in the grain yield of rice plants exposed to heat: A review. PeerJ 2019, 7, e7792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.M.; Zheng, H.X.; Zhang, X.S.; Sui, N. Cytokinins as central regulators during plant growth and stress response. Plant Cell Rep. 2021, 40, 271–282. [Google Scholar] [CrossRef]
- Wang, Y.; Li, B.; Du, M.W.; Eneji, A.E.; Wang, B.M.; Duan, L.S.; Li, Z.H.; Tian, X.L. Mechanism of phytohormone involvement in feedback regulation of cotton leaf senescence induced by potassium deficiency. J. Exp. Bot. 2012, 63, 5887–5901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannarelli, S.; Muscatello, B.; Bogani, P.; Spiriti, M.M.; Buiatti, M.; Fuoco, R. Comparative determination of some phytohormones in wild-type and genetically modified plants by gas chromatography-mass spectrometry and high-performance liquid chromatography-tandem mass spectrometry. Anal. Biochem. 2010, 398, 60–68. [Google Scholar] [CrossRef]
- Song, W.; Li, J.; Sun, H.; Huang, S.; Gong, X.; Ma, Q.; Zhang, Y.; Xu, G. Increased photosynthetic capacity in response to nitrate is correlated with enhanced cytokinin levels in rice cultivar with high responsiveness to nitrogen nutrients. Plant Soil 2013, 373, 981–993. [Google Scholar] [CrossRef]
- Cai, W.J.; Ye, T.T.; Wang, Q.; Cai, B.D.; Feng, Y.Q. A rapid approach to investigate spatiotemporal distribution of phytohormones in rice. Plant Methods 2016, 12, 47. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.L.; Dong, C.X.; Dong, Y.Y.; Xiong, C.L.; Shen, Q.R. Simultaneous determination of six endogenous cytokinins components and auxin in plant tissue by high performance liquid chromatography. Plant Nutr. Fertil. Sci. 2007, 13, 129–135. [Google Scholar]
- Chou, C.C.; Chen, W.S.; Huang, K.L.; Yu, H.C.; Liao, L.J. Changes in cytokinin levels of Phalaenopsis leaves at high temperature. Plant Physiol. Biochem. 2000, 38, 309–314. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, L.; Zhang, W.; Deng, Z.; Wang, F.; Li, Y.; Meng, L.; Wu, N.; Ban, F.; Zhang, S. Simultaneous analysis of two phytohormones in chili and wheat using HPLC using novel calixarene as SPE sorbent. J. Chromatogr. Sci. 2017, 55, 358–365. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; He, M.; Chen, B.; Hu, B. Simultaneous determination of acidic phytohormones in cucumbers and green bean sprouts by ion-pair stir bar sorptive extraction-high performance liquid chromatography. Talanta 2017, 170, 128–136. [Google Scholar] [CrossRef]
- Wu, C.; Cui, K.; Wang, W.; Li, Q.; Fahad, S.; Hu, Q.; Huang, J.; Nie, L.; Peng, S. Heat-induced phytohormone changes are associated with disrupted early reproductive development and reduced yield in rice. Sci. Rep. 2016, 6, 34978. [Google Scholar] [CrossRef]
- Zhang, Z.; Peng, J.; Li, S.; Peng, H.; Wang, X.; Long, D.; Chen, J.; Xian, H.; Ni, R. Simultaneous determination of cytokinins by high performance liquid chromatography with resonance Rayleigh scattering and mechanism discussion. Analyst 2019, 144, 5186–5192. [Google Scholar] [CrossRef]
- Du, F.; Sun, L.; Zhen, X.; Nie, H.; Zheng, Y.; Ruan, G.; Li, J. High-internal-phase-emulsion polymeric monolith coupled with liquid chromatography–electrospray tandem mass spectrometry for enrichment and sensitive detection of trace cytokinins in plant samples. Anal. Bioanal. Chem. 2015, 407, 6071–6079. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Cui, K.; Wang, W.; Li, Q.; Fahad, S.; Hu, Q.; Huang, J.; Nie, L.; Mohapatra, P.K.; Peng, S. Heat-induced cytokinin transportation and degradation are associated with reduced panicle cytokinin expression and fewer spikelets per panicle in rice. Front. Plant Sci. 2017, 8, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoyerová, K.; Gaudinová, A.; Malbeck, J.; Dobrev, P.I.; Kocábek, T.; Šolcová, B.; Trávníčková, A.; Kamínek, M. Efficiency of different methods of extraction and purification of cytokinins. Phytochemistry 2006, 67, 1151–1159. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.; Chattha, M.; Khan, I.; Chattha, B.; Barbanti, L.; Aamer, M.; Iqbal, M.; Nawaz, M.; Mahmood, A.; Ali, A.; et al. Heat stress in cultivated plants: Nature, impact, mechanisms, and mitigation strategies—A review. Plant Biosyst. 2020, 1–56. [Google Scholar] [CrossRef]
- Bose, A. HPLC calibration process parameters in terms of system suitability test. Austin Chromatogr. 2014, 1, 1–4. [Google Scholar]
- Brenner, M.L. Modern methods for plant growth substance analysis. Annu. Rev. Plant Phys. 2003, 32, 511–538. [Google Scholar] [CrossRef]
- Wu, Y.; Hu, B. Simultaneous determination of several phytohormones in natural coconut juice by hollow fiber-based liquid-liquid-liquid microextraction-high performance liquid chromatography. J. Chromatogr. A 2009, 1216, 7657–7663. [Google Scholar] [CrossRef] [PubMed]
- Chiang, P.C.; Hu, Y. Simultaneous determination of LogD, LogP, and pKa of drugs by using a reverse phase HPLC coupled with a 96-Well plate auto injector. Comb. Chem. High Throughput Screen. 2009, 12, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Zhang, Y.Z. Determination of plant intrinsic hormones by reversed phase high performance liquid chromatography. J. Instrum. Anal. 2001, 20, 61–62. [Google Scholar]
- Naqqash, T.; Hameed, S.; Imran, A.; Hanif, M.K.; Majeed, A.; Dirk, J.; Elsas, V. Differential growth stimulation response of potato towards inoculation with taxonomically diverse plant growth promoting rhizobacteria. Front. Plant Sci. 2016, 7, 144. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.Y.; Chen, Y.H.; Chen, M.X.; Wang, Z.Q.; Wu, C.F.; Bian, X.C.; Yang, J.C.; Zhang, J.H. Growth characteristics and endosperm structure of superior and inferior spikelets of indica rice under high-temperature stress. Biol. Plant. 2016, 60, 532–542. [Google Scholar] [CrossRef]
- Liu, H.; Li, X.; Xiao, J.; Wang, S. A convenient method for simultaneous quantification of multiple phytohormones and metabolites: Application in study of rice-bacterium interaction. Plant Methods 2012, 8, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida-Trapp, M.; Mithöfer, A. Quantification of phytohormones by HPLC-MS/MS including phytoplasma-infected plants. Methods Mol. Biol. 2019, 1875, 345–358. [Google Scholar] [PubMed]
- Chen, J.; Tang, H.H.; Li, L.; Qin, S.J.; Wang, G.P.; Hong, N. Effects of virus infection on plant growth, root development and phytohormone levels in in vitro-cultured pear plants. Plant Cell Tiss. Org. 2017, 131, 359–368. [Google Scholar] [CrossRef]
- Kudo, T.; Kiba, T.; Sakakibara, H. Metabolism and long-distance translocation of cytokinins. J. Integr. Plant Biol. 2010, 52, 53–60. [Google Scholar] [CrossRef]
- Takei, K.; Sakakibara, H.; Taniguchi, M.; Sugiyama, T. Nitrogen-dependent accumulation of cytokinins in root and the translocation to leaf: Implication of cytokinin species that induces gene expression of maize response regulator. Plant Cell Physiol. 2001, 42, 85–93. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (min) | Methanol (%) | ddH2O (%) | 4.5% Acetic Acid (%) |
|---|---|---|---|
| 0 | 0 | 100 | 0 |
| 17 | 30 | 70 | 0 |
| 18 | 40 | 60 | 0 |
| 22 | 40 | 60 | 0 |
| 24 | 35 | 65 | 0 |
| 25 | 35 | 0 | 65 |
| 34 | 35 | 0 | 65 |
| 35 | 0 | 100 | 0 |
| 45 | 0 | 100 | 0 |
| Compound | Calibration Equation | r | Limit of Detection (ug/L) (S/N = 3) | Recovery (%) | RSD (%) |
|---|---|---|---|---|---|
| tZ9G | y = 47.397x − 0.4608 | 0.998 | 0.774 | 90.9 | 3.4 |
| tZ | y = 87.393x + 0.4238 | 0.999 | 0.475 | 109.4 | 8.6 |
| diZ | y = 156.59x + 0.0892 | 1.000 | 0.421 | 102.2 | 8.7 |
| tZR | y = 45.161x + 0.0508 | 0.999 | 0.959 | 97.8 | 7.8 |
| diZR | y = 64.128x − 0.5331 | 0.998 | 0.845 | 99.1 | 8.1 |
| iPMP | y = 44.345x − 0.0941 | 0.999 | 1.087 | 62.1 | 7.1 |
| IAA | y = 32.701x + 0.0265 | 0.999 | 1.071 | 82.5 | 9.5 |
| iP9G | y = 73.153x − 0.0861 | 0.999 | 0.824 | 96.7 | 8.5 |
| iP | y = 91.569x − 0.6583 | 0.999 | 0.214 | 96.7 | 7.4 |
| iPA | y = 55.29x − 0.0045 | 0.999 | 0.893 | 97.2 | 8.9 |
| ABA | y = 85.338x − 0.3667 | 0.999 | 1.772 | 99.3 | 8.8 |
| Compound | Inter-Day | Intra-Day | ||
|---|---|---|---|---|
| Retention Time | Peak Area | Retention Time | Peak Area | |
| tZ9G | 0.41 | 2.65 | 1.00 | 9.05 |
| tZ | 0.87 | 1.25 | 1.71 | 3.50 |
| diZ | 0.65 | 0.70 | 1.68 | 0.98 |
| tZR | 0.75 | 2.87 | 0.97 | 2.16 |
| diZR | 0.38 | 1.65 | 0.80 | 2.86 |
| iPMP | 0.60 | 2.05 | 1.23 | 9.13 |
| IAA | 0.82 | 2.01 | 1.45 | 5.81 |
| iP9G | 0.22 | 1.14 | 0.22 | 4.26 |
| iP | 0.46 | 0.70 | 0.74 | 1.43 |
| iPA | 0.41 | 1.60 | 0.71 | 0.68 |
| ABA | 0.28 | 1.31 | 0.36 | 4.04 |
| Compound | Nagina22 | Huanghuazhan | ||||
|---|---|---|---|---|---|---|
| Young Panicle (ng/g FW) | Root (ng/g FW) | Xylem Sap (ng/L) | Young Panicle (ng/g FW) | Root (ng/g FW) | Xylem Sap (ng/L) | |
| tZ9G | 111.2 ± 8.0 | 355.0 ± 14.2 | ND | 183.7 ± 8.8 | 351.2 ± 8.2 | ND |
| tZ | 37.3 ± 2.9 | 62.6 ± 2.4 | 312.5 ± 43.2 | 57.5 ± 2.1 | 61.4 ± 2.6 | 435.6 ± 24.6 |
| diZ | ND | ND | ND | ND | ND | ND |
| tZR | 76.4 ± 1.9 | 301.0 ± 22.6 | 986.4 ± 37.2 | 111.4 ± 7.2 | 664.4 ± 41.1 | 1264.4 ± 78.6 |
| diZR | ND | ND | ND | ND | ND | ND |
| iPMP | 137.5 ± 11.3 | 178.1 ± 18.8 | 78.1 ± 9.1 | 165.3 ± 2.7 | 140.9 ± 9.1 | 97.8 ± 7.9 |
| IAA | 133.9 ± 6.5 | 99.2 ± 5.3 | 730.9 ± 91.5 | 122.8 ± 6.1 | 117.3 ± 5.0 | 1292.8 ± 42.7 |
| iP9G | 34.2 ± 2.1 | 74.0 ± 5.7 | ND | 12.7 ± 0.9 | 104.3 ± 1.7 | ND |
| iP | 16.3 ± 1.2 | 9.1 ± 0.3 | 19.4 ± 1.1 | 4.7 ± 0.3 | 8.8 ± 0.3 | 34.1 ± 3.3 |
| iPA | 68.8 ± 6.5 | 45.2 ± 2.0 | 89.2 ± 10.2 | 37.5 ± 2.8 | 50.8 ± 3.3 | 124.9 ± 3.8 |
| ABA | 86.7 ± 5.8 | 72.0 ± 3.9 | 273.6 ± 16.8 | 58.3 ± 3.9 | 82.7 ± 4.8 | 261.7 ± 23.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, B.; Wu, C.; Liang, H.; Cui, K.; Fahad, S.; Wang, M.; Liu, B.; Nie, L.; Huang, J.; Tang, H. Optimized High-Performance Liquid Chromatography Method for Determining Nine Cytokinins, Indole-3-acetic Acid and Abscisic Acid. Sustainability 2021, 13, 6998. https://0-doi-org.brum.beds.ac.uk/10.3390/su13136998
Qi B, Wu C, Liang H, Cui K, Fahad S, Wang M, Liu B, Nie L, Huang J, Tang H. Optimized High-Performance Liquid Chromatography Method for Determining Nine Cytokinins, Indole-3-acetic Acid and Abscisic Acid. Sustainability. 2021; 13(13):6998. https://0-doi-org.brum.beds.ac.uk/10.3390/su13136998
Chicago/Turabian StyleQi, Beibei, Chao Wu, Huiling Liang, Kehui Cui, Shah Fahad, Manlian Wang, Baoyu Liu, Lixiao Nie, Jianliang Huang, and Hui Tang. 2021. "Optimized High-Performance Liquid Chromatography Method for Determining Nine Cytokinins, Indole-3-acetic Acid and Abscisic Acid" Sustainability 13, no. 13: 6998. https://0-doi-org.brum.beds.ac.uk/10.3390/su13136998