
Alleviation of Cadmium Phytotoxicity Using Silicon Fertilization in Wheat by Altering Antioxidant Metabolism and Osmotic Adjustment

 , , ,
, , ,  , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Site, Experimental Design and Treatments
2.2. Crop Management
2.3. Growth Attributes
2.4. Non-Enzymatic Antioxidant and Physiobiochemical Attributes
2.5. Antioxidant Enzymes Activities
2.6. Cadmium and Silicon Contents and Statistics Analysis
3. Results
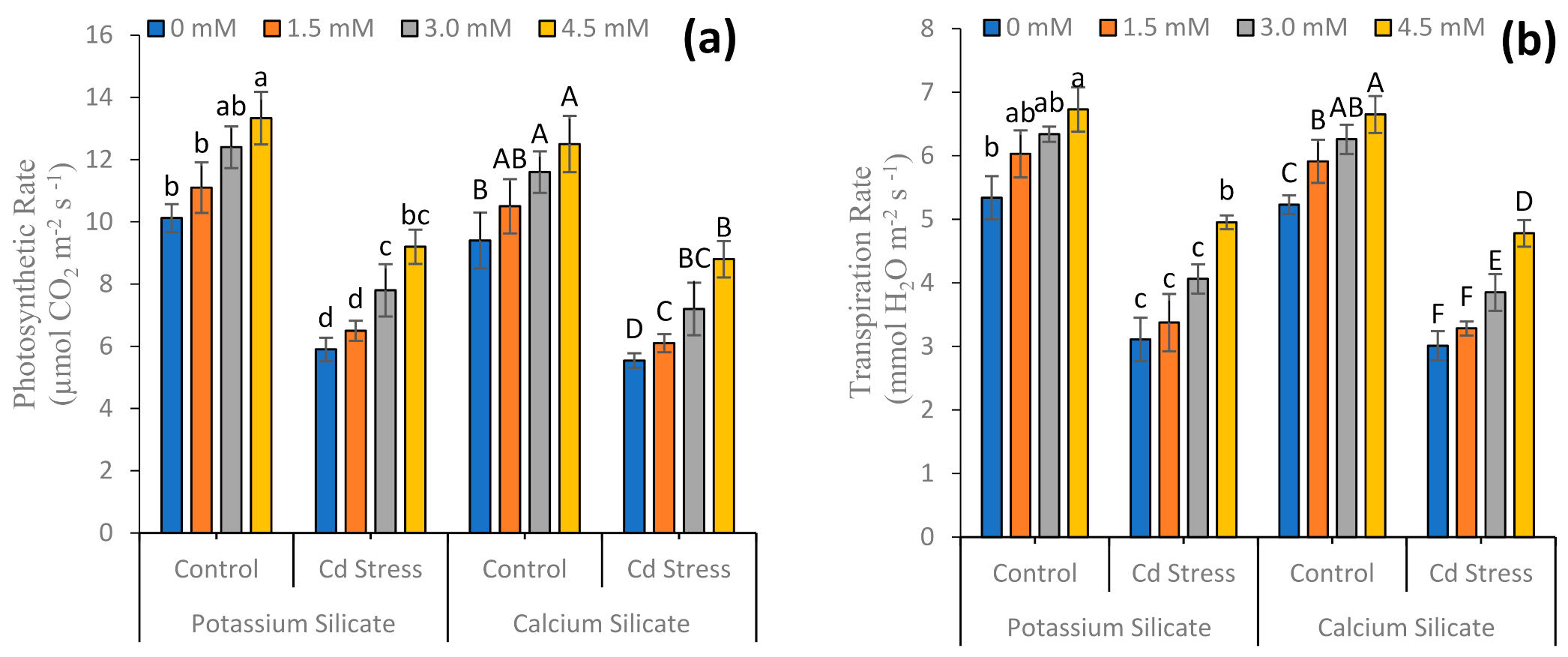
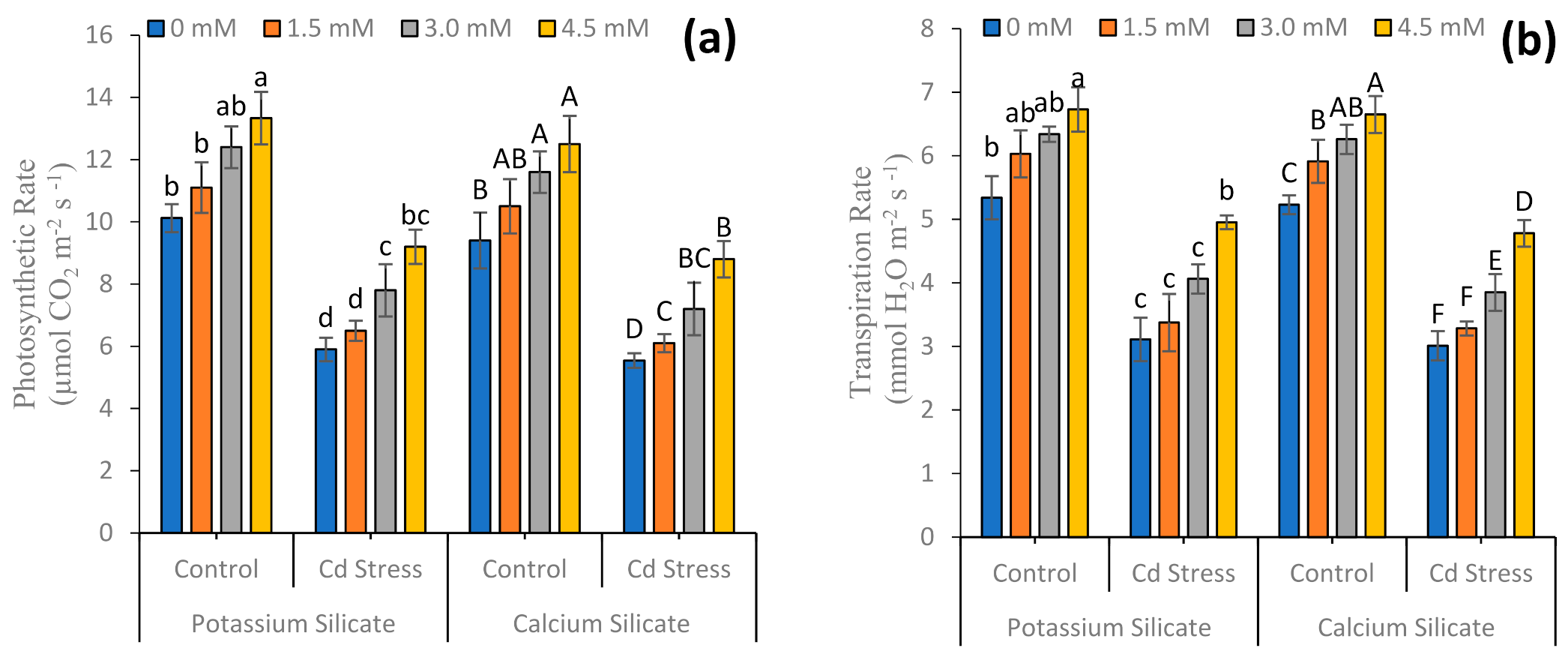
3.1. Growth Attributes
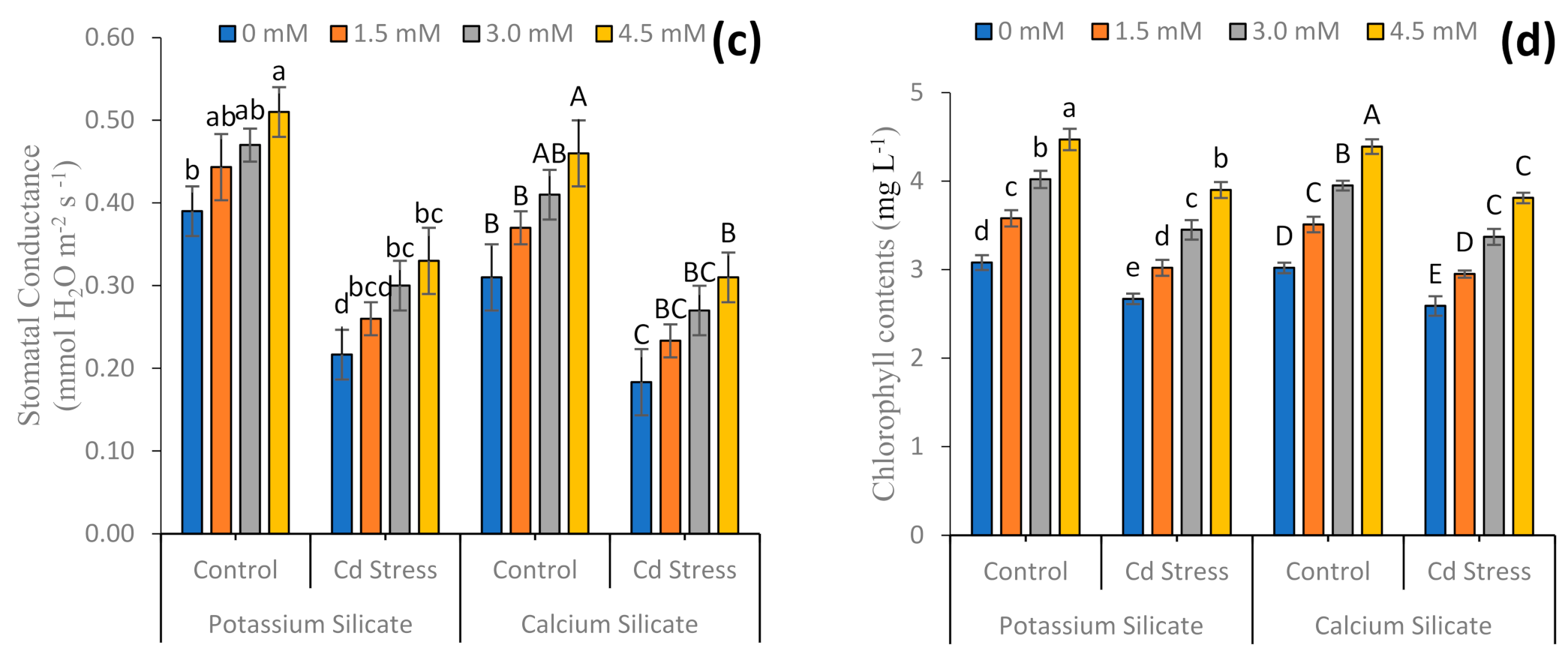
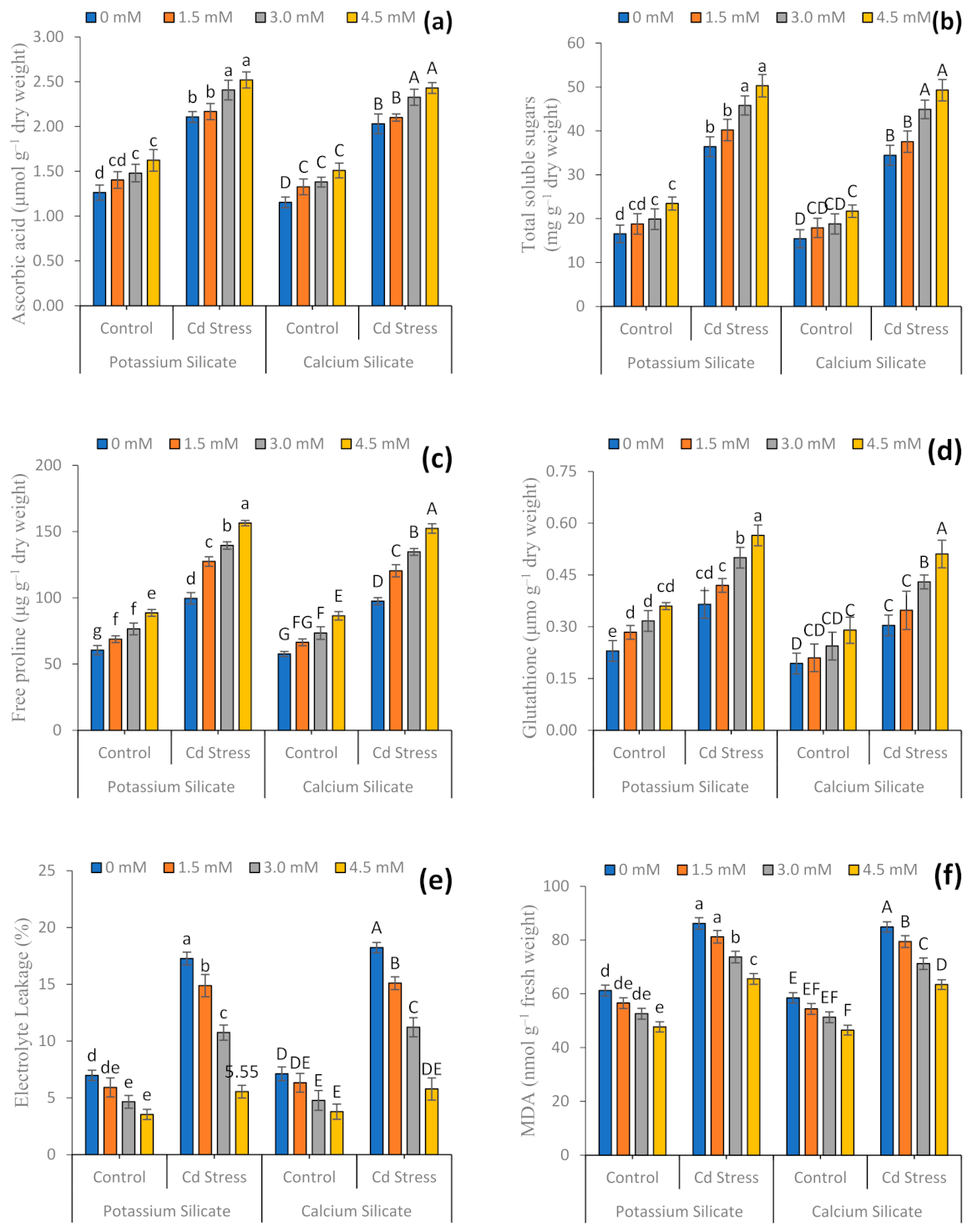
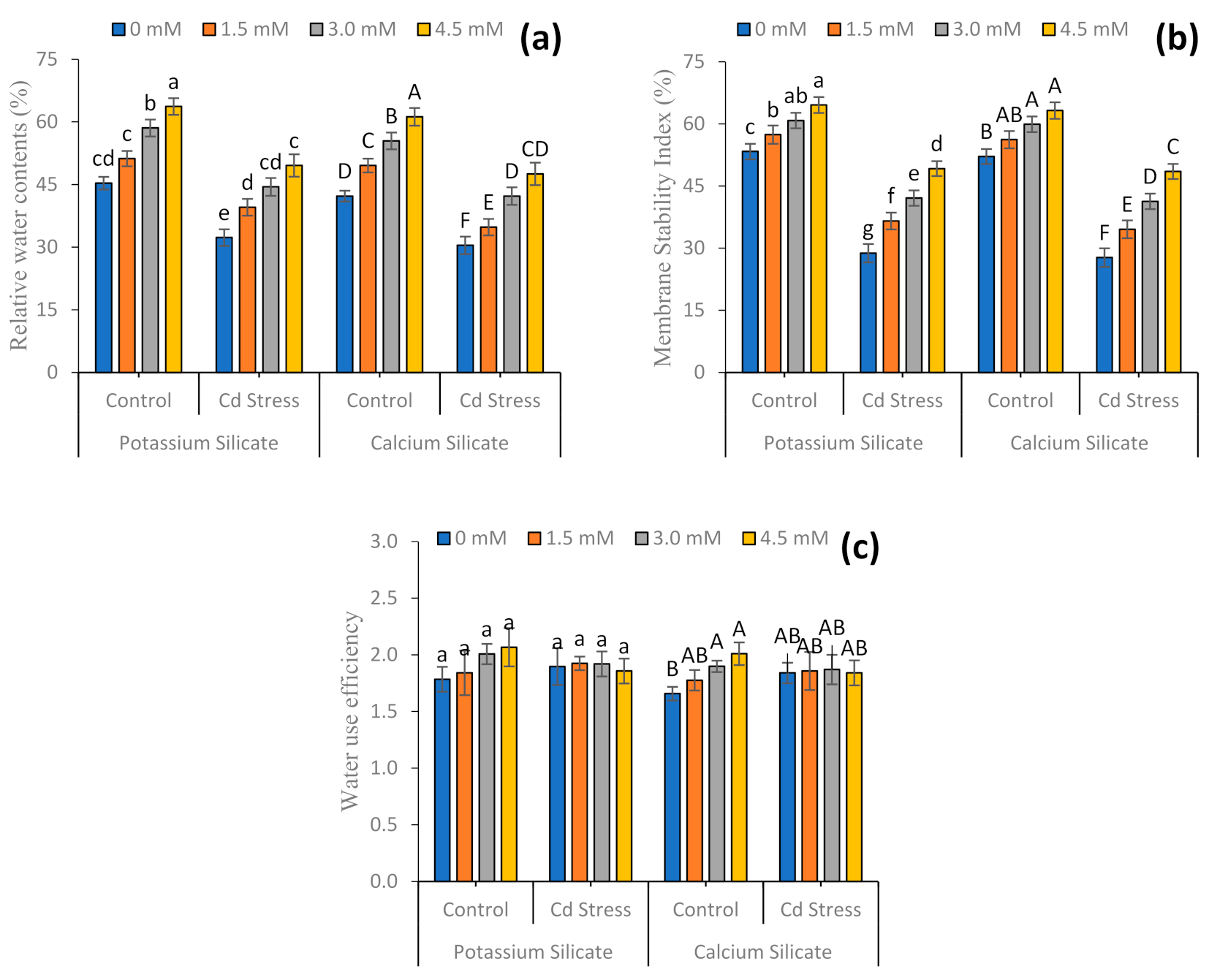
3.2. Non-Enzymatic Antioxidant and Physio-Biochemical Attributes
3.3. Enzymatic Antioxidant Attributes
3.4. Cadmium and Silicon Contents in Plant Parts
4. Discussion
4.1. Plants Growth and Biomass
4.2. Physiological and Biochemical Parameters
4.3. Relative Water Contents and Electrolyte Leakage
4.4. Enzymatic and Non-Enzymatic Antioxidants
4.5. Cadmium and Si Accumulation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgment
Conflicts of Interest
References
- Tariq, F.; Wang, X.; Saleem, M.H.; Khan, Z.I.; Ahmad, K.; Saleem Malik, I.; Munir, M.; Mahpara, S.; Mehmood, N.; Ahmad, T.; et al. Risk Assessment of Heavy Metals in Basmati Rice: Implications for Public Health. Sustainability 2021, 13, 8513. [Google Scholar] [CrossRef]
- Mao, C.; Song, Y.; Chen, L.; Ji, J.; Li, J.; Yuan, X.; Yang, Z.; Ayoko, G.A.; Frost, R.L.; Theiss, F. Human health risks of heavy metals in paddy rice based on transfer characteristics of heavy metals from soil to rice. Catena 2019, 175, 339–348. [Google Scholar] [CrossRef]
- Zaheer, I.E.; Ali, S.; Saleem, M.H.; Noor, I.; El-Esawi, M.A.; Hayat, K.; Rizwan, M.; Abbas, Z.; El-Sheikh, M.A.; Alyemeni, M.N. Iron–Lysine Mediated Alleviation of Chromium Toxicity in Spinach (Spinacia oleracea L.) Plants in Relation to Morpho-Physiological Traits and Iron Uptake When Irrigated with Tannery Wastewater. Sustainability 2020, 12, 6690. [Google Scholar] [CrossRef]
- Aziz, H.; Wang, X.; Murtaza, G.; Ashar, A.; Hussain, S.; Abid, M.; Murtaza, B.; Saleem, M.H.; Fiaz, S.; Ali, S. Evaluation of Compost and Biochar to Mitigate Chlorpyrifos Pollution in Soil and Their Effect on Soil Enzyme Dynamics. Sustainability 2021, 13, 9695. [Google Scholar] [CrossRef]
- Kusvuran, S.; Kiran, S.; Ellialtioglu, S.S. Antioxidant Enzyme Activities and Abiotic Stress Tolerance Relationship in Vegetable Crops. In Abiotic and Biotic Stress in Plants—Recent Advances and Future Perspectives, 1st ed.; InTech: Rijeka, Croatia, 2016; pp. 481–503. [Google Scholar]
- Rigby, H.; Smith, S.R. The significance of cadmium entering the human food chain via livestock ingestion from the agricultural use of biosolids, with special reference to the UK. Environ. Int. 2020, 143, 105844. [Google Scholar] [CrossRef]
- Kim, Y.-H.; Khan, A.L.; Lee, I.-J. Silicon: A duo synergy for regulating crop growth and hormonal signaling under abiotic stress conditions. Crit. Rev. Biotechnol. 2016, 36, 1099–1109. [Google Scholar] [CrossRef]
- Javed, M.T.; Saleem, M.H.; Aslam, S.; Rehman, M.; Iqbal, N.; Begum, R.; Ali, S.; Alsahli, A.A.; Alyemeni, M.N.; Wijaya, L. Elucidating silicon-mediated distinct morpho-physio-biochemical attributes and organic acid exudation patterns of cadmium stressed Ajwain (Trachyspermum ammi L.). Plant Physiol. Biochem. 2020, 157, 23–37. [Google Scholar] [CrossRef]
- Shah, V.; Daverey, A. Effects of sophorolipids augmentation on the plant growth and phytoremediation of heavy metal contaminated soil. J. Clean. Prod. 2021, 280, 124406. [Google Scholar] [CrossRef]
- Chen, L.; Hu, W.-f.; Long, C.; Wang, D. Exogenous plant growth regulator alleviate the adverse effects of U and Cd stress in sunflower (Helianthus annuus L.) and improve the efficacy of U and Cd remediation. Chemosphere 2021, 262, 127809. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Hussain, A.; Ali, Q.; Shakoor, M.B.; Zia-ur-Rehman, M.; Farid, M.; Asma, M. Effect of zinc-lysine on growth, yield and cadmium uptake in wheat (Triticum aestivum L.) and health risk assessment. Chemosphere 2017, 187, 35–42. [Google Scholar] [CrossRef]
- Hussain, A.; Rizwan, M.; Ali, Q.; Ali, S. Seed priming with silicon nanoparticles improved the biomass and yield while reduced the oxidative stress and cadmium concentration in wheat grains. Environ. Sci. Pollut. Res. 2019, 26, 7579–7588. [Google Scholar] [CrossRef]
- Hussain, A.; Rizwan, M.; Ali, S.; ur Rehman, M.Z.; Qayyum, M.F.; Nawaz, R.; Ahmad, A.; Asrar, M.; Ahmad, S.R.; Alsahli, A.A. Combined use of different nanoparticles effectively decreased cadmium (Cd) concentration in grains of wheat grown in a field contaminated with Cd. Ecotoxicol. Environ. Saf. 2021, 215, 112139. [Google Scholar] [CrossRef]
- Beres, B.L.; Hatfield, J.L.; Kirkegaard, J.A.; Eigenbrode, S.D.; Pan, W.L.; Lollato, R.P.; Hunt, J.R.; Strydhorst, S.; Porker, K.; Lyon, D. Towards a better understanding of Genotype × Environment × Management interactions—A global Wheat Initiative agronomic research strategy. Front. Plant Sci. 2020, 11, 828. [Google Scholar] [CrossRef]
- Pequeno, D.N.L.; Hernandez-Ochoa, I.M.; Reynolds, M.; Sonder, K.; Molero-Milan, A.; Robertson, R.; Lopes, M.d.S.S.; Xiong, W.; Kropff, M.; Asseng, S. Climate impact and adaptation to heat and drought stress of regional and global wheat production. Environ. Res. Lett. 2021, 16, 54070. [Google Scholar] [CrossRef]
- Irfan, M.; Hayat, S.; Ahmad, A.; Alyemeni, M.N. Soil cadmium enrichment: Allocation and plant physiological manifestations. Saudi J. Biol. Sci. 2013, 20, 1–10. [Google Scholar] [CrossRef]
- Kelkar, D.S.; Kumar, D.; Kumar, P.; Balakrishnan, L.; Muthusamy, B.; Yadav, A.K.; Shrivastava, P.; Marimuthu, A.; Anand, S.; Sundaram, H. Proteogenomic analysis of Mycobacterium tuberculosis by high resolution mass spectrometry. Mol. Cell. Proteom. 2011, 10, M111.011445. [Google Scholar] [CrossRef] [Green Version]
- Rizwan, M.; Meunier, J.-D.; Davidian, J.-C.; Pokrovsky, O.; Bovet, N.; Keller, C. Silicon alleviates Cd stress of wheat seedlings (Triticum turgidum L. cv. Claudio) grown in hydroponics. Environ. Sci. Pollut. Res. 2016, 23, 1414–1427. [Google Scholar] [CrossRef]
- Ali, A.; Guo, D.; Li, Y.; Shaheen, S.M.; Wahid, F.; Antoniadis, V.; Abdelrahman, H.; Al-Solaimani, S.G.; Li, R.; Tsang, D.C. Streptomyces pactum addition to contaminated mining soils improved soil quality and enhanced metals phytoextraction by wheat in a green remediation trial. Chemosphere 2021, 273, 129692. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, C.; Du, B.; Cui, H.; Fan, X.; Zhou, D.; Zhou, J. Soil and foliar applications of silicon and selenium effects on cadmium accumulation and plant growth by modulation of antioxidant system and Cd translocation: Comparison of soft vs. durum wheat varieties. J. Hazard. Mater. 2021, 402, 123546. [Google Scholar] [CrossRef]
- Hu, B.; Xue, J.; Zhou, Y.; Shao, S.; Fu, Z.; Li, Y.; Chen, S.; Qi, L.; Shi, Z. Modelling bioaccumulation of heavy metals in soil-crop ecosystems and identifying its controlling factors using machine learning. Environ. Pollut. 2020, 262, 114308. [Google Scholar] [CrossRef]
- Greger, M.; Kabir, A.H.; Landberg, T.; Maity, P.J.; Lindberg, S. Silicate reduces cadmium uptake into cells of wheat. Environ. Pollut. 2016, 211, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Rizwan, M.; Hussain, A.; ur Rehman, M.Z.; Ali, B.; Yousaf, B.; Wijaya, L.; Alyemeni, M.N.; Ahmad, P. Silicon nanoparticles enhanced the growth and reduced the cadmium accumulation in grains of wheat (Triticum aestivum L.). Plant Physiol. Biochem. 2019, 140, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Fahad, S.; Hussain, S.; Matloob, A.; Khan, F.A.; Khaliq, A.; Saud, S.; Hassan, S.; Shan, D.; Khan, F.; Ullah, N. Phytohormones and plant responses to salinity stress: A review. Plant Growth Regul. 2015, 75, 391–404. [Google Scholar] [CrossRef]
- Silva, A.; Nascimento, C.; Gouveia-Neto, A. Assessment of cadmium phytotoxicity alleviation by silicon using chlorophyll a fluorescence. Photosynthetica 2017, 55, 648–654. [Google Scholar] [CrossRef]
- Putko, P.; Kwaśny, M. Bioavailable silicon forms in dietary supplements. Biul. Wojsk. Akad. Tech. 2020, 69, 35–41. [Google Scholar]
- Gill, R.A.; Zang, L.; Ali, B.; Farooq, M.A.; Cui, P.; Yang, S.; Ali, S.; Zhou, W. Chromium-induced physio-chemical and ultrastructural changes in four cultivars of Brassica napus L. Chemosphere 2015, 120, 154–164. [Google Scholar] [CrossRef]
- Hussain, A.; Ali, S.; Rizwan, M.; ur Rehman, M.Z.; Hameed, A.; Hafeez, F.; Alamri, S.A.; Alyemeni, M.N.; Wijaya, L. Role of zinc–lysine on growth and chromium uptake in rice plants under Cr stress. J. Plant Growth Regul. 2018, 37, 1413–1422. [Google Scholar] [CrossRef]
- Ahmad, R.; Ali, S.; Rizwan, M.; Dawood, M.; Farid, M.; Hussain, A.; Wijaya, L.; Alyemeni, M.N.; Ahmad, P. Hydrogen sulfide alleviates chromium stress on cauliflower by restricting its uptake and enhancing antioxidative system. Physiol. Plant. 2020, 168, 289–300. [Google Scholar] [CrossRef] [Green Version]
- Nagata, M.; Yamashita, I. Simple method for simultaneous determination of chlorophyll and carotenoids in tomato fruit. Nippon Shokuhin Kogyo Gakkaishi 1992, 39, 925–928. [Google Scholar] [CrossRef] [Green Version]
- Lutts, S.; Guerrier, G. Peroxidase activities of two rice cultivars differing in salinity tolerance as affected by proline and NaCl. Biol. Plant. 1995, 37, 577–586. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, C.; Lu, T.; Zheng, Y. Cooperation between arbuscular mycorrhizal fungi and earthworms promotes the physiological adaptation of maize under a high salt stress. Plant Soil 2018, 423, 125–140. [Google Scholar] [CrossRef]
- Irigoyen, J.; Einerich, D.; Sánchez-Díaz, M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativd) plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Mukherjee, S.; Choudhuri, M. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant. 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Griffith, O.W. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine. Anal. Biochem. 1980, 106, 207–212. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A. Assay of catalases and peroxidases. Methods Enzymol. 1955, 2, 764–775. [Google Scholar]
- Rady, M.M. Effect of 24-epibrassinolide on growth, yield, antioxidant system and cadmium content of bean (Phaseolus vulgaris L.) plants under salinity and cadmium stress. Sci. Hortic. 2011, 129, 232–237. [Google Scholar] [CrossRef]
- Treder, W.; Cieslinski, G. Effect of silicon application on cadmium uptake and distribution in strawberry plants grown on contaminated soils. J. Plant Nutr. 2005, 28, 917–929. [Google Scholar] [CrossRef]
- Jia, H.; Hou, D.; O’Connor, D.; Pan, S.; Zhu, J.; Bolan, N.S.; Mulder, J. Exogenous phosphorus treatment facilitates chelation-mediated cadmium detoxification in perennial ryegrass (Lolium perenne L.). J. Hazard. Mater. 2020, 389, 121849. [Google Scholar] [CrossRef]
- Bent, E. Silicon Solutions. Bone 2008, 43, 596–606. [Google Scholar]
- Xu, L.; Zeng, W.; Li, J.; Liu, H.; Yan, G.; Si, P.; Yang, C.; Shi, Y.; He, Q.; Zhou, W. Characteristics of membrane-bound fatty acid desaturase (FAD) genes in Brassica napus L. and their expressions under different cadmium and salinity stresses. Environ. Exp. Bot. 2019, 162, 144–156. [Google Scholar] [CrossRef]
- Bharwana, S.A.; Ali, S.; Farooq, M.A.; Iqbal, N.; Hameed, A.; Abbas, F.; Ahmad, M.S.A. Glycine betaine-induced lead toxicity tolerance related to elevated photosynthesis, antioxidant enzymes suppressed lead uptake and oxidative stress in cotton. Turk. J. Bot. 2014, 38, 281–292. [Google Scholar] [CrossRef]
- Li, N.; Feng, A.; Liu, N.; Jiang, Z.; Wei, S. Silicon application improved the yield and nutritional quality while reduced cadmium concentration in rice. Environ. Sci. Pollut. Res. 2020, 27, 20370–20379. [Google Scholar] [CrossRef]
- Hamayun, M.; Sohn, E.-Y.; Khan, S.A.; Shinwari, Z.K.; Khan, A.L.; Lee, I.-J. Silicon alleviates the adverse effects of salinity and drought stress on growth and endogenous plant growth hormones of soybean (Glycine max L.). Pak. J. Bot 2010, 42, 1713–1722. [Google Scholar]
- Adrees, M.; Ali, S.; Rizwan, M.; Zia-ur-Rehman, M.; Ibrahim, M.; Abbas, F.; Farid, M.; Qayyum, M.F.; Irshad, M.K. Mechanisms of silicon-mediated alleviation of heavy metal toxicity in plants: A review. Ecotoxicol. Environ. Saf. 2015, 119, 186–197. [Google Scholar] [CrossRef]
- Ceccarelli, M.; Barthel, F.P.; Malta, T.M.; Sabedot, T.S.; Salama, S.R.; Murray, B.A.; Morozova, O.; Newton, Y.; Radenbaugh, A.; Pagnotta, S.M. Molecular profiling reveals biologically discrete subsets and pathways of progression in diffuse glioma. Cell 2016, 164, 550–563. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Ma, C.; Ma, Y.; Li, H.; Kang, J.; Liu, T.; Polle, A.; Peng, C.; Luo, Z.-B. Cadmium tolerance in six poplar species. Environ. Sci. Pollut. Res. 2013, 20, 163–174. [Google Scholar] [CrossRef]
- Mateos-Naranjo, E.; Andrades-Moreno, L.; Davy, A.J. Silicon alleviates deleterious effects of high salinity on the halophytic grass Spartina densiflora. Plant Physiol. Biochem. 2013, 63, 115–121. [Google Scholar] [CrossRef]
- Zanella, L.; Fattorini, L.; Brunetti, P.; Roccotiello, E.; Cornara, L.; D’Angeli, S.; Della Rovere, F.; Cardarelli, M.; Barbieri, M.; Di Toppi, L.S. Overexpression of AtPCS1 in tobacco increases arsenic and arsenic plus cadmium accumulation and detoxification. Planta 2016, 243, 605–622. [Google Scholar] [CrossRef]
- Rios, J.J.; Martínez-Ballesta, M.C.; Ruiz, J.M.; Blasco, B.; Carvajal, M. Silicon-mediated improvement in plant salinity tolerance: The role of aquaporins. Front. Plant Sci. 2017, 8, 948. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Xu, P.; Zeng, G.; Yang, C.; Huang, D.; Zhang, J. Bioremediation of soils contaminated with polycyclic aromatic hydrocarbons, petroleum, pesticides, chlorophenols and heavy metals by composting: Applications, microbes and future research needs. Biotechnol. Adv. 2015, 33, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Ming, D.; Pei, Z.; Naeem, M.; Gong, H.; Zhou, W. Silicon alleviates PEG-induced water-deficit stress in upland rice seedlings by enhancing osmotic adjustment. J. Agron. Crop Sci. 2012, 198, 14–26. [Google Scholar] [CrossRef]
- Slabbert, M.; Krüger, G. Antioxidant enzyme activity, proline accumulation, leaf area and cell membrane stability in water stressed Amaranthus leaves. S. Afr. J. Bot. 2014, 95, 123–128. [Google Scholar] [CrossRef] [Green Version]
- Kabir, A.H.; Hossain, M.M.; Khatun, M.A.; Mandal, A.; Haider, S.A. Role of silicon counteracting cadmium toxicity in alfalfa (Medicago sativa L.). Front. Plant Sci. 2016, 7, 1117. [Google Scholar] [CrossRef]
- Coskun, D.; Britto, D.T.; Huynh, W.Q.; Kronzucker, H.J. The role of silicon in higher plants under salinity and drought stress. Front. Plant Sci. 2016, 7, 1072. [Google Scholar] [CrossRef] [Green Version]
- Mahmood, A.; Malik, R.N. Human health risk assessment of heavy metals via consumption of contaminated vegetables collected from different irrigation sources in Lahore, Pakistan. Arab. J. Chem. 2014, 7, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Ranieri, A.; Castagna, A.; Scebba, F.; Careri, M.; Zagnoni, I.; Predieri, G.; Pagliari, M.; di Toppi, L.S. Oxidative stress and phytochelatin characterisation in bread wheat exposed to cadmium excess. Plant Physiol. Biochem. 2005, 43, 45–54. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Ibrahim, M.; Farid, M.; Adrees, M.; Bharwana, S.A.; Zia-ur-Rehman, M.; Qayyum, M.F.; Abbas, F. Mechanisms of silicon-mediated alleviation of drought and salt stress in plants: A review. Environ. Sci. Pollut. Res. 2015, 22, 15416–15431. [Google Scholar] [CrossRef]
- Li, G.Z.; Chen, S.J.; Li, N.Y.; Wang, Y.Y.; Kang, G.Z. Exogenous Glutathione Alleviates Cadmium Toxicity in Wheat by Influencing the Absorption and Translocation of Cadmium. Bull. Environ. Contam. Toxicol. 2021, 107, 1–7. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Nazir, F.; Asgher, M.; Per, T.S.; Khan, N.A. Selenium and sulfur influence ethylene formation and alleviate cadmium-induced oxidative stress by improving proline and glutathione production in wheat. J. Plant Physiol. 2015, 173, 9–18. [Google Scholar] [CrossRef]
- Anjum, N.A.; Umar, S.; Ahmad, A.; Iqbal, M.; Khan, N.A. Sulphur protects mustard (Brassica campestris L.) from cadmium toxicity by improving leaf ascorbate and glutathione. Plant Growth Regul. 2008, 54, 271–279. [Google Scholar] [CrossRef]
- Liu, J.; Platts-Mills, J.A.; Juma, J.; Kabir, F.; Nkeze, J.; Okoi, C.; Operario, D.J.; Uddin, J.; Ahmed, S.; Alonso, P.L. Use of quantitative molecular diagnostic methods to identify causes of diarrhoea in children: A reanalysis of the GEMS case-control study. Lancet 2016, 388, 1291–1301. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Treatments | Seedling Fresh Weight (g) | Seedling Dry Weight (g) | Leaf Area (cm2) |
|---|---|---|---|
| Heavy Metals Stress (S) | |||
| S1 = Control | 4.03 A | 0.85 A | 25.51 A |
| S2 = Cadmium Stress (20 mg kg−1 of Soil) | 1.80 B | 0.39 B | 13.57 B |
| HSD (S) (p ≤ 0.01) | 1.14 | 0.27 | 2.15 |
| Silicate Chemicals (C) | |||
| C1 = Potassium Silicate | 3.30 | 0.65 | 20.07 |
| C2 = Calcium Silicate | 2.53 | 0.60 | 19.02 |
| HSD (C) (p ≤ 0.01) | 1.14 | 0.27 | 2.15 |
| Foliar Application Treatments (T) | |||
| T1 = Control | 2.49 D | 0.56 D | 16.42 C |
| T2 = 1.50 mM foliar application | 2.79 C | 0.59 C | 18.05 BC |
| T3 = 3.00 mM foliar application | 3.03 B | 0.65 B | 20.86 AB |
| T4 = 4.50 mM foliar application | 3.37 A | 0.70 A | 22.84 A |
| HSD (T) (p ≤ 0.01) | 0.20 | 0.03 | 4.06 |
| Significance Level (S) | 15.20 ** | 11.83 ** | 126.86 ** |
| Significance Level (C) | 1.86 NS | 0.13 NS | 0.97 NS |
| Significance Level (T) | 0.43 ** | 0.23 ** | 7.30 ** |
| Significance Level (S × C) | 0.02 NS | 0.00 NS | 0.20 NS |
| Significance Level (S × T) | 0.06 NS | 0.00 NS | 0.01 NS |
| Significance Level (C × T) | 0.00 NS | 0.00 NS | 0.02 NS |
| Significance Level (S × C × T) | 0.00 NS | 0.00 NS | 0.00 NS |
| Experimental Treatments | SOD (U µg−1 Protein) | POD (µg g−1 Fresh Weight min−1) | CAT (U mg−1 min−1) |
|---|---|---|---|
| Heavy Metals Stress (S) | |||
| S1 = Control | 40.50 B | 79.60 B | 0.36 B |
| S2 = Cadmium Stress (20 mg kg−1 of Soil) | 71.76 A | 140.86 A | 0.75 A |
| HSD (S) (p ≤ 0.01) | 2.37 | 1.91 | 0.07 |
| Silicate Chemicals (C) | |||
| C1 = Potassium Silicate | 56.57 | 110.70 | 0.59 |
| C2 = Calcium Silicate | 55.68 | 109.76 | 0.52 |
| HSD (C) (p ≤ 0.01) | 2.37 | 1.91 | 0.07 |
| Foliar Application Treatments (T) | |||
| T1 = Control | 50.60 B | 98.40 C | 0.43 C |
| T2 = 1.50 mM foliar application | 52.57 B | 100.82 C | 0.49 BC |
| T3 = 3.00 mM foliar application | 58.65 A | 111.80 B | 0.61 AB |
| T4 = 4.50 mM foliar application | 62.70 A | 129.90 A | 0.73 A |
| HSD (T) (p ≤ 0.01) | 4.47 | 3.61 | 0.14 |
| Significance Level (S) | 716.85 ** | 4213.51 ** | 102.91 ** |
| Significance Level (C) | 0.58 NS | 0.99 NS | 3.69 NS |
| Significance Level (T) | 22.67 ** | 231.19 ** | 14.16 ** |
| Significance Level (S × C) | 0.00 NS | 0.01 NS | 0.00 NS |
| Significance Level (S × T) | 7.30 ** | 117.14 ** | 0.07 NS |
| Significance Level (C × T) | 0.01 NS | 0.00 NS | 0.00 NS |
| Significance Level (S × C × T) | 0.01 NS | 0.00 NS | 0.00 NS |
| Experimental Treatments | Cd Content in Root (mg kg−1 Dry Weight) | Cd Content in Leaf (mg kg−1 Dry Weight) | Si Content in Root (mg g−1 Dry Weight) | Si Content in Leaf (mg g−1 Dry Weight) |
|---|---|---|---|---|
| Heavy Metals Stress (S) | ||||
| S1 = Control | 0.98 B | 0.56 B | 0.44 A | 2.25 A |
| S2 = Cadmium Stress (20 mg kg−1 of Soil) | 62.56 A | 34.23 A | 0.12 B | 0.96 B |
| HSD (S) (p ≤ 0.01) | 2.34 | 3.12 | 0.19 | 1.23 |
| Silicate Chemicals (C) | ||||
| C1 = Potassium Silicate | 43.76 | 27.45 | 0.39 | 2.23 |
| C2 = Calcium Silicate | 44.56 | 29.34 | 0.37 | 2.12 |
| HSD (C) (p ≤ 0.01) | 2.34 | 3.12 | 0.19 | 1.23 |
| Foliar Application Treatments (T) | ||||
| T1 = Control | 67.72 A | 43.26 A | 0.15 D | 0.48 D |
| T2 = 1.50 mM foliar application | 55.34 B | 30.23 B | 0.28 C | 1.97 C |
| T3 = 3.00 mM foliar application | 29.89 C | 14.56 C | 0.39 B | 3.23 B |
| T4 = 4.50 mM foliar application | 14.23 D | 5.50 D | 0.54 A | 5.12 A |
| HSD (T) (p ≤ 0.01) | 4.34 | 4.67 | 0.07 | 1.12 |
| Significance Level (S) | 99.91 ** | 87.91 ** | 102.91 ** | 67.91 ** |
| Significance Level (C) | 2.69 NS | 3.59 NS | 3.69 NS | 3.29 NS |
| Significance Level (T) | 11.16 ** | 4.16 ** | 14.16 ** | 1.16 ** |
| Significance Level (S × C) | 0.03 NS | 0.02 NS | 0.00 NS | 0.03 NS |
| Significance Level (S × T) | 0.05 NS | 0.07 NS | 0.07 NS | 0.07 NS |
| Significance Level (C × T) | 0.00 NS | 0.01 NS | 0.00 NS | 0.02 NS |
| Significance Level (S × C × T) | 0.00 NS | 0.01 NS | 0.00 NS | 0.05 NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heile, A.O.; Zaman, Q.u.; Aslam, Z.; Hussain, A.; Aslam, M.; Saleem, M.H.; Abualreesh, M.H.; Alatawi, A.; Ali, S. Alleviation of Cadmium Phytotoxicity Using Silicon Fertilization in Wheat by Altering Antioxidant Metabolism and Osmotic Adjustment. Sustainability 2021, 13, 11317. https://0-doi-org.brum.beds.ac.uk/10.3390/su132011317
Heile AO, Zaman Qu, Aslam Z, Hussain A, Aslam M, Saleem MH, Abualreesh MH, Alatawi A, Ali S. Alleviation of Cadmium Phytotoxicity Using Silicon Fertilization in Wheat by Altering Antioxidant Metabolism and Osmotic Adjustment. Sustainability. 2021; 13(20):11317. https://0-doi-org.brum.beds.ac.uk/10.3390/su132011317
Chicago/Turabian StyleHeile, Abdihakim Osman, Qamar uz Zaman, Zubair Aslam, Afzal Hussain, Mustansar Aslam, Muhammad Hamzah Saleem, Muyassar H. Abualreesh, Aishah Alatawi, and Shafaqat Ali. 2021. "Alleviation of Cadmium Phytotoxicity Using Silicon Fertilization in Wheat by Altering Antioxidant Metabolism and Osmotic Adjustment" Sustainability 13, no. 20: 11317. https://0-doi-org.brum.beds.ac.uk/10.3390/su132011317