
What Will Remain? Predicting the Representation in Protected Areas of Suitable Habitat for Endangered Tropical Avifauna in Borneo under a Combined Climate- and Land-Use Change Scenario


_Collins.png)
Abstract
:1. Introduction
2. Materials and Methods
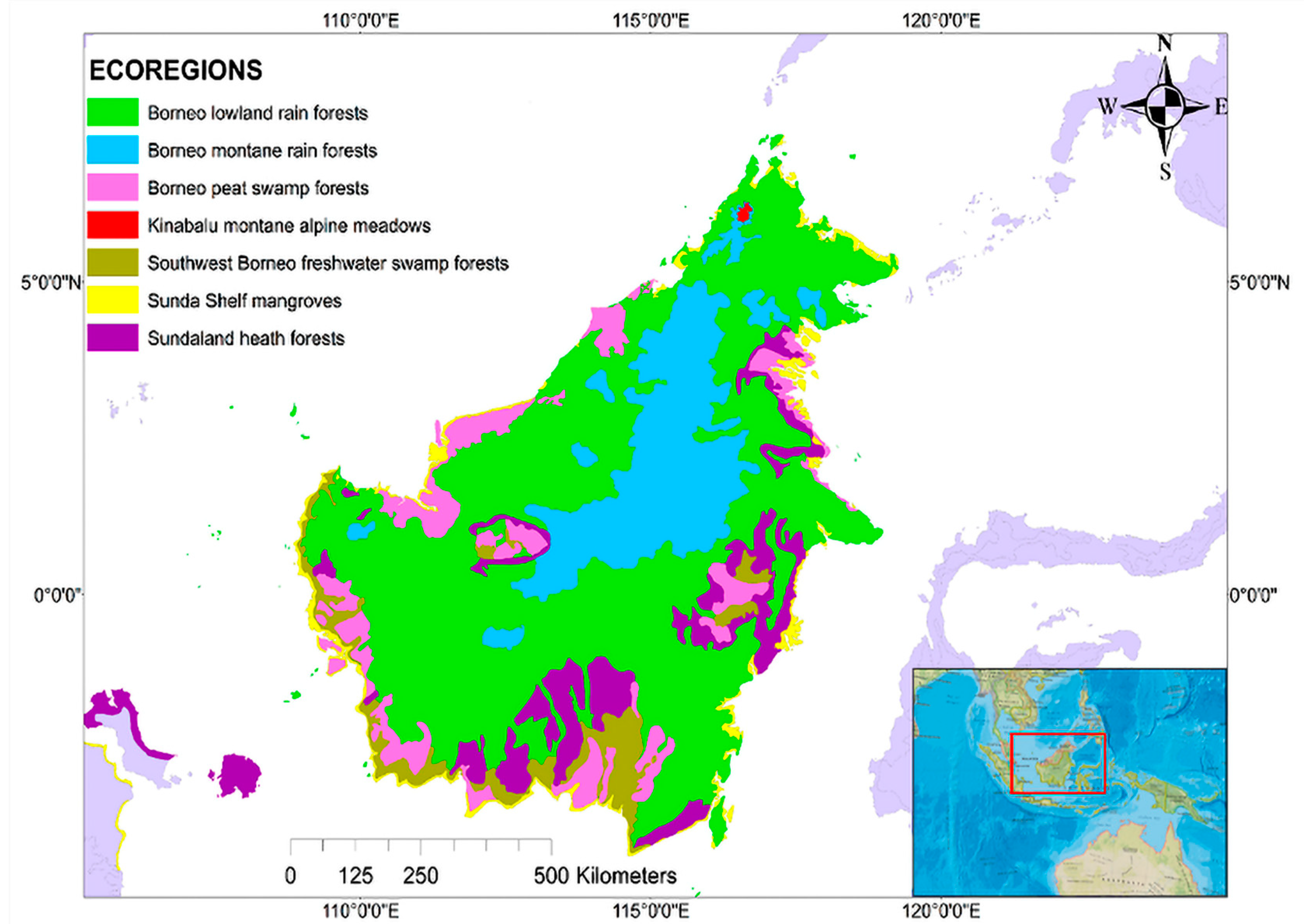
2.1. Study Site
2.2. Focal Species Selection
2.3. Environmental Data
2.3.1. Present Climate Data
2.3.2. Future Climate Data
2.3.3. Topographic Data
2.3.4. Land-Use Data
2.3.5. Layer Preparation
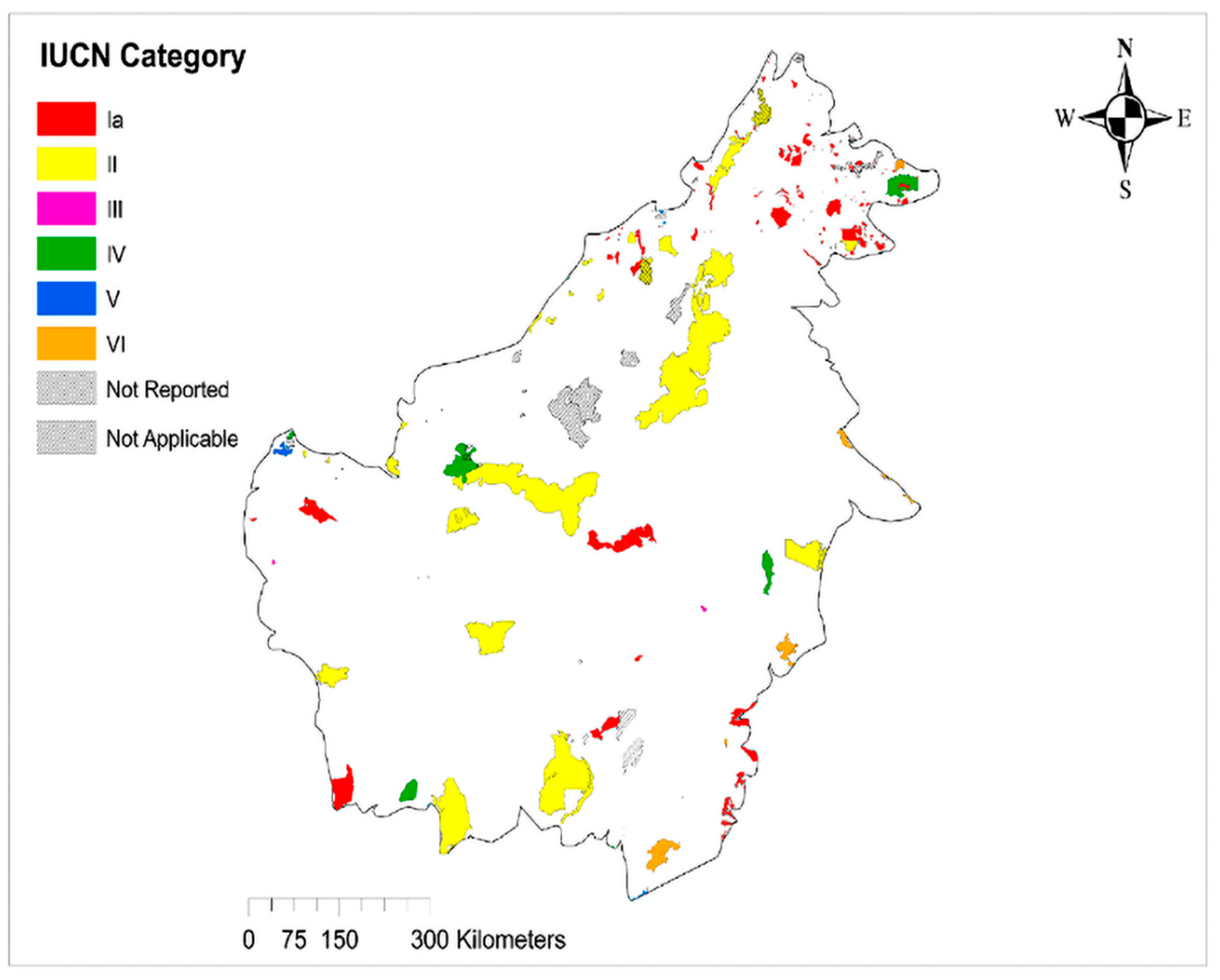
2.3.6. Protected Area Data
2.3.7. Habitat Suitability Modelling
2.3.8. Habitat Suitability and Protected Area Coverage Calculation and Analysis
3. Results
3.1. Current (2010) Habitat Suitability Modelling and Estimated Portion Protected
3.2. Future Habitat Suitability Modelling and Estimated Portion Protected
4. Discussion
4.1. Changes in the Area of Suitable Habitat
4.2. Representation of Threatened Avifauna in the BORNEAN PA Network
4.3. Conservation Implications and Recommendations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sala, O.E.; Chapin, F.S.; Armesto, J.J.; Berlow, E.; Bloomfield, J.; Dirzo, R.; Huber-Sanwald, E.; Huenneke, L.F.; Jackson, R.B.; Kinzig, A.; et al. Global biodiversity scenarios for the year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Foden, W.B.; Butchart, S.H.M.; Stuart, S.N.; Vié, J.-C.; Akçakaya, H.R.; Angulo, A.; DeVantier, L.M.; Gutsche, A.; Turak, E.; Cao, L.; et al. Identifying the World’s Most Climate Change Vulnerable Species: A Systematic Trait-Based Assessment of all Birds, Amphibians and Corals. PLoS ONE 2013, 8, e65427. [Google Scholar] [CrossRef] [Green Version]
- Curran, L.M.; Trigg, S.N.; McDonald, A.K.; Astiani, D.; Hardiono, Y.M.; Siregar, P.; Caniago, I.; Kasischke, E. Lowland Forest Loss in Protected Areas of Indonesian Borneo. Science 2004, 303, 1000–1003. [Google Scholar] [CrossRef] [Green Version]
- Hughes, A.C. Understanding the drivers of Southeast Asian biodiversity loss. Ecosphere 2017, 8, e01624. [Google Scholar] [CrossRef]
- Brodie, J.; Post, E.; Laurance, W.F. Climate change and tropical biodiversity: A new focus. Trends Ecol. Evol. 2012, 27, 145–150. [Google Scholar] [CrossRef]
- Padoa-Schioppa, E.; Baietto, M.; Massa, R.; Bottoni, L. Bird communities as bioindicators: The focal species concept in agricultural landscapes. Ecol. Indic. 2006, 6, 83–93. [Google Scholar] [CrossRef]
- Srinivasan, U.; Wilcove, D.S. Interactive impacts of climate change and land-use change on the demography of montane birds. Ecology 2021, 102, e03223. [Google Scholar] [CrossRef]
- Sekercioglu, C.H.; Schneider, S.H.; Fay, J.P.; Loarie, S.R. Climate Change, Elevational Range Shifts, and Bird Extinctions. Conserv. Biol. 2007, 22, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Smeraldo, S.; Bosso, L.; Fraissinet, M.; Bordignon, L.; Brunelli, M.; Ancillotto, L.; Russo, D. Modelling risks posed by wind turbines and power lines to soaring birds: The black stork (Ciconia nigra) in Italy as a case study. Biodivers. Conserv. 2020, 29, 1959–1976. [Google Scholar] [CrossRef]
- Machar, I.; Poprach, K.; Harmacek, J.; Fialova, J. Bird Diversity as a Support Decision Tool for Sustainable Management in Temperate Forested Floodplain Landscapes. Sustainability 2019, 11, 1527. [Google Scholar] [CrossRef] [Green Version]
- Sohl, T.L. The Relative Impacts of Climate and Land-Use Change on Conterminous United States Bird Species from 2001 to 2075. PLoS ONE 2014, 9, e112251. [Google Scholar] [CrossRef] [PubMed]
- Wauchope, H.; Shaw, J.; Varpe, Ø.; Lappo, E.; Boertmann, D.; Lanctot, R.; Fuller, R.A. Rapid climate-driven loss of breeding habitat for Arctic migratory birds. Glob. Chang. Biol. 2017, 23, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Titeux, N.; Henle, K.; Mihoub, J.-B.; Regos, A.; Geijzendorffer, I.R.; Cramer, W.; Verburg, P.H.; Brotons, L. Biodiversity scenarios neglect future land-use changes. Glob. Chang. Biol. 2016, 22, 2505–2515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva, M.X.; Paviolo, A.; Tambosi, L.; Pardini, R. Effectiveness of Protected Areas for biodiversity conservation: Mammal occupancy patterns in the Iguaçu National Park, Brazil. J. Nat. Conserv. 2018, 41, 51–62. [Google Scholar] [CrossRef]
- Di Minin, E.; Toivonen, T. Global Protected Area Expansion: Creating More than Paper Parks. Bioscience 2015, 65, 637–638. [Google Scholar] [CrossRef] [Green Version]
- Tantipisanuh, N.; Savini, T.; Cutter, P.; Gale, G.A. Biodiversity gap analysis of the protected area system of the Indo-Burma Hotspot and priorities for increasing biodiversity representation. Biol. Conserv. 2016, 195, 203–213. [Google Scholar] [CrossRef]
- Hannah, L.; Midgley, G.; Andelman, S.; Araújo, M.; Hughes, G.; Martinez-Meyer, E.; Pearson, R.; Williams, P. Protected area needs in a changing climate. Front. Ecol. Environ. 2007, 5, 131–138. [Google Scholar] [CrossRef]
- Dinerstein, E.; Olson, D.; Joshi, A.; Vynne, C.; Burgess, N.D.; Wikramanayake, E.; Hahn, N.; Palminteri, S.; Hedao, P.; Noss, R.; et al. An Ecoregion-Based Approach to Protecting Half the Terrestrial Realm. Bioscience 2017, 67, 534–545. [Google Scholar] [CrossRef]
- Sodhi, N.S.; Smith, K.G. Conservation of tropical birds: Mission possible? J. Ornithol. 2007, 148, 305–309. [Google Scholar] [CrossRef]
- Radosavljevic, A.; Anderson, R.P. Making better Maxent models of species distributions: Complexity, overfitting and evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]
- Hughes, A.; Satasook, C.; Bates, P.; Bumrungsri, S.; Jones, G. The projected effects of climatic and vegetation changes on the distribution and diversity of Southeast Asian bats. Glob. Chang. Biol. 2012, 18, 1854–1865. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Porfirio, L.; Harris, R.; Lefroy, E.; Hugh, S.; Gould, S.; Lee, G.; Bindoff, N.L.; Mackey, B. Improving the use of species distribution models in con-servation planning and management under climate change. PLoS ONE 2014, 9, e113749. [Google Scholar] [CrossRef] [Green Version]
- Avalos, V.d.R.; Hernández, J. Projected distribution shifts and protected area coverage of range-restricted Andean birds under climate change. Glob. Ecol. Conserv. 2015, 4, 459–469. [Google Scholar] [CrossRef]
- Li, X.; Chen, G.; Liu, X.; Liang, X.; Wang, S.; Chen, Y.; Pei, F.; Xu, X. A New Global Land-Use and Land-Cover Change Product at a 1-km Resolution for 2010 to 2100 Based on Human–Environment Interactions. Ann. Am. Assoc. Geogr. 2017, 107, 1040–1059. [Google Scholar] [CrossRef]
- Singh, M.; Cheyne, S.; Ehlers Smith, D. How conspecific primates use their habitats: Surviving in an anthropogenically-disturbed forest in Central Kalimantan, Indonesia. Ecol. Indic. 2018, 87, 167–177. [Google Scholar] [CrossRef]
- Pasquale, G.D.; Saracino, A.; Bosso, L.; Russo, D.; Moroni, A.; Bonanomi, G.; Allevato, E. Coastal pine-oak glacial refugia in the Mediterranean basin: A biogeographic approach based on charcoal analysis and spatial modelling. Forests 2020, 11, 673. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Srivastava, V.; Roe, A.D.; Keena, M.A.; Hamelin, R.C.; Griess, V.C. Oh the places they’ll go: Improving species distribution modelling for invasive forest pests in an uncertain world. Biol. Invasions 2021, 23, 297–349. [Google Scholar] [CrossRef]
- Phillips, S.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Urbina-Cardona, J.; Loyola, R. Applying Niche-Based Models to Predict Endangered-Hylid Potential Distributions: Are Neotropical Protected Areas Effective Enough? Trop. Conserv. Sci. 2008, 1, 417–445. [Google Scholar] [CrossRef]
- Cameron, A.; Trivedi, P.K. Microeconometrics: Methods and Applications; Cambridge University Press: Cambridge, NY, USA, 2005. [Google Scholar]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Peterson, A.; Papeş, M.; Soberón, J. Rethinking receiver operating characteristic analysis applications in ecological niche modeling. Ecol Model. 2008, 213, 63–72. [Google Scholar] [CrossRef]
- Wisz, M.; Hijmans, R.; Li, J.; Peterson, A.; Graham, C.; Guisan, A.; NCEAS Predicting Species Distributions Working Group. Effects of sample size on the performance of species distribution models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- Coxen, C.L.; Frey, J.K.; Carleton, S.A.; Collins, D.P. Species distribution models for a migratory bird based on citizen science and satellite tracking data. Glob. Ecol. Conserv. 2017, 11, 298–311. [Google Scholar] [CrossRef]
- Nazeri, M.; Jusoff, K.; Madani, N.; Mahmud, A.; Bahman, A.; Kumar, L. Predictive modeling and mapping of Malayan Sun Bear (Helarctos malayanus) distribution using maximum entropy. PLoS ONE 2012, 7, e48104. [Google Scholar] [CrossRef] [Green Version]
- Escalante, T.; Rodríguez-Tapia, G.; Linaje, M.; Illoldi-Rangel, P.; González-López, R. Identification of areas of endemism from species distribution models: Threshold selection and Nearctic mammals. TIP 2013, 16, 5–17. [Google Scholar] [CrossRef]
- Freeman, B.; Sunnarborg, J.; Peterson, A.T. Effects of climate change on the distributional potential of three range-restricted West African bird species. Condor 2019, 121. [Google Scholar] [CrossRef]
- R Foundation for Statistical Computing VA. R Foundation for Statistical Computing RCT. R: A Language and Environment for Statistical Computing. Austria: R Foundation for Statistical Computing, Vienna, Austria. 2020. Available online: https://www.R-project.org/ (accessed on 1 March 2020).
- Singh, M. Evaluating the impact of future climate and forest cover change on the ability of Southeast (SE) Asia’s protected areas to provide coverage to the habitats of threatened avian species. Ecol. Indic. 2020, 114, 106307. [Google Scholar] [CrossRef]
- Brus, J.; Pechanec, V.; Machar, I. Depiction of uncertainty in the visually interpreted land cover data. Ecol. Inform. 2018, 47, 10–13. [Google Scholar] [CrossRef]
- Struebig, M.J.; Fischer, M.; Gaveau, D.L.; Meijaard, E.; Wich, S.A.; Gonner, C.; Sykes, R.; Wilting, A.; Kramer-Schadt, S. Anticipated climate and land-cover changes reveal refuge areas for Borneo’s orang-utans. Glob. Chang. Biol. 2015, 21, 2891–2904. [Google Scholar] [CrossRef]
- Alamgir, M.; Campbell, M.J.; Sloan, S.; Suhardiman, A.; Supriatna, J.; Laurance, W.F. High-risk infrastructure projects pose imminent threats to forests in Indonesian Borneo. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Mantyka-Pringle, C.; Visconti, P.; Di Marco, M.; Martin, T.; Rondinini, C.; Rhodes, J. Climate change modifies risk of global bio-diversity loss due to land-cover change. Biol. Conserv. 2015, 187, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Scriven, S.; Hodgson, J.; McClean, C.; Hill, J. Protected areas in Borneo may fail to conserve tropical forest biodiversity under climate change. Biol. Conserv. 2015, 184, 414–423. [Google Scholar] [CrossRef] [Green Version]
- Freeman, B.; Class Freeman, A. Rapid upslope shifts in New Guinean birds illustrate strong distributional responses of tropical montane species to global warming. Proc. Natl. Acad. Sci. USA 2014, 111, 4490–4494. [Google Scholar] [CrossRef] [Green Version]
- Freeman, B.; Scholer, M.; Ruiz-Gutierrez, V.; Fitzpatrick, J. Climate change causes upslope shifts and mountaintop extirpa-tions in a tropical bird community. Proc. Natl. Acad. Sci. USA 2018, 115, 11982–11987. [Google Scholar] [CrossRef]
- La Sorte, F.; Jetz, W. Projected range contractions of montane biodiversity under global warming. Proc. Biol. Sci. 2010, 277, 3401–3410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colwell, R.; Brehm, G.; Cardelús, C.; Gilman, A.; Longino, J. Global warming, elevational range shifts, and lowland biotic attrition in the wet tropics. Science 2008, 322, 258–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez-Villegas, J.; Cuesta, F.; Devenish, C.; Peralvo, M.; Jarvis, A.; Arnillas, C. Using species distributions models for designing conservation strategies of Tropical Andean biodiversity under climate change. J. Nat. Conserv. 2014, 22, 391–404. [Google Scholar] [CrossRef] [Green Version]
- Jose, V.S.; Nameer, P. The expanding distribution of the Indian Peafowl (Pavo cristatus) as an indicator of changing climate in Kerala, southern India: A modelling study using Maxent. Ecol. Indic. 2020, 110, 105930. [Google Scholar] [CrossRef]
- Velásquez-Tibatá, J.; Salaman, P.; Graham, C. Effects of climate change on species distribution, community structure, and conservation of birds in protected areas in Colombia. Reg. Environ. Chang. 2013, 13, 235–248. [Google Scholar] [CrossRef]
- Rodrigues, A.; Akçakaya, H.; Andelman, S.; Bakarr, M.; Boitani, L.; Brooks, T.; Chanson, J.S.; Fishpool, L.D.C.; Da Fonseca, G.A.B.; Gaston, K.J.; et al. Global Gap Analysis: Priority Regions for Expanding the Global Protected-Area Network. Bioscience 2004, 54, 1092. [Google Scholar] [CrossRef]
- Cazalis, V.; Princé, K.; Mihoub, J.B.; Kelly, J.; Butchart, S.; Rodrigues, A. Effectiveness of protected areas in conserving tropical forest birds. Nat. Commun. 2020, 11, 4461. [Google Scholar] [CrossRef] [PubMed]
- Bosso, L.; Smeraldo, S.; Rapuzzi, P.; Sama, G.; Garonna, A.P.; Russo, D. Nature protection areas of Europe are insufficient to preserve the threatened beetle Rosalia alpina (Coleoptera: Cerambycidae): Evidence from species distribution models and conservation gap analysis. Ecol. Entomol. 2018, 43, 192–203. [Google Scholar] [CrossRef]
- Hickcox, R.P.; Jara, M.; Deacon, L.A.K.; Harvey, L.P.; Pincheira-Donoso, D. Global terrestrial distribution of penguins (Spheniscidae) and their conservation by protected areas. Biodivers. Conserv. 2019, 28, 2861–2876. [Google Scholar] [CrossRef]
- De Klerk, H.; Fjeldså, J.; Blyth, S.; Burgess, N. Gaps in the protected area network for threatened Afrotropical birds. Biol. Conserv. 2004, 117, 529–537. [Google Scholar] [CrossRef] [Green Version]
- Gaüzère, P.; Jiguet, F.; Devictor, V. Can protected areas mitigate the impacts of climate change on bird’s species and communities? Divers. Distrib. 2016, 22, 625–637. [Google Scholar] [CrossRef] [Green Version]
- Garden, J.; O’Donnell, T.; Catterall, C. Changing habitat areas and static reserves: Challenges to species protection under climate change. Landsc. Ecol. 2015, 30, 1959–1973. [Google Scholar] [CrossRef]
- Wan, J.; Wang, C.; Han, S.; Yu, J. Planning the priority protected areas of endangered orchid species in northeastern China. Biodivers. Conserv. 2014, 23, 1395–1409. [Google Scholar] [CrossRef]
- Newbold, T. Future effects of climate and land-use change on terrestrial vertebrate community diversity under different scenarios. Proc. Biol. Sci. 2018, 285, 1881. [Google Scholar] [CrossRef]
- Sharma, S.; Baral, H.; Laumonier, Y.; Okarda, B.; Komarudin, H.; Purnomo, H.; Pacheco, P. Ecosystem services under future oil palm expansion scenarios in West Kalimantan, Indonesia. Ecosyst. Serv. 2019, 39, 100978. [Google Scholar] [CrossRef]
- Savolainen, V.; Clottey, V.A.; Doubi, B.T.S.; Konan, J.L.; Quain, M.; Bezeng, B.S.; Logah, V.; Wireko-Kena, A.; Osekre, E.A.; Atuah, L.; et al. Systems thinking creates opportunities for a circular economy and sustainable palm agriculture in Africa. Curr. Res. Environ. Sustain. 2020, 1, 31–34. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species (Latin) | Species (Common) | IUCN Category | Habitat Preference |
|---|---|---|---|
| Aceros corrugatus | WrinkledHornbill | EN | Lowland evergreen forest extends into the selectively logged forest (Kemp, A.C, Boesman, P and Sharpe, 2019) |
| Anthracoceros malayanus | Black hornbill | VU | Lowland primary forest, mostly <200 m, very occasionally up to 600 m (Kemp, Boesman and Sharpe, 2019) |
| Berenicornis comatus | White-crowned hornbill | EN | Primary lowland and sometimes montane forest up to 1675 m (A. C. Kemp and Boesman, 2019b) |
| Buceros rhinoceros | Rhinoceros hornbill | VU | Primary lowland forest, extending into the secondary forest up to 1400 m (A. C. Kemp and Boesman, 2019a) |
| Caprimulgus concretus | Bonaparte’s Nightjar | VU | Lowland dipterocarp forest and secondary growth, sea level to <500 m (Cleere and Sharpe, 2019) |
| Centropus rectunguis | Centropus rectunguis | VU | The undergrowth of lowland forest (Payne and de Juana, 2019) |
| Chloropsis sonnerati | Greater green leafbird | VU | Lowland evergreen forest, extending up to 1100 m (Wells and Sharpe, 2019) |
| Ciconia stormi | Storm’s stork | EN | Undisturbed lowland forest |
| Cyornis caerulatus | Sunda Blue Flycatcher | VU | Lowland dipterocarp forest, sea-level to <500 m (Clement, 2019) |
| Hydrornis baudii | blue-headed pitta | VU | Lowland primary forest extends up to 600 m (Erritzoe, 2019) |
| Leptoptilos javanicus | Lesser adjutant | VU | Mangroves, coastland wetlands adjacent to lowland forest |
| Mulleripicus pulverulentus | Great slaty woodpecker | VU | Semi-open tropical lowland forests, extending up to 2000 m (Winkler, Christie and Kirwan, 2019) |
| Nisaetus nanus | Wallace’s hawk-eagle | VU | Lowland forest, rarely up to 1000 m (Clark and Kirwan, 2019) |
| Psittacula longicauda | Long-tailed parakeet | VU | Extreme lowland, <300 m (Collar and Sharpe, 2019) |
| Ptilocichla leucogrammica | Bornean wren-babbler | VU | Lowland dipterocarp forest extends into the montane forest (Collar and Robson, 2019) |
| Rhinoplax vigil | Helmeted hornbill | CR | Extends up to 1500 m (Kemp, Sharpe and Boesman, 2019) |
| Rhyticeros undulatus | Wreathed hornbill | VU | Occasionally lowland forest extends up to 2560 m (A. Kemp and Boesman, 2019) |
| Setornis criniger | hook-billed bulbul | VU | Lowland forest, extending up to 1000 m (Myers, 2016b) |
| Spilornis kinabaluensis | Mountain serpent eagle | VU | Montane and sub-montane forests, preferably ridgetops, 750–2900 m (Clark, Kirwan and Christie, 2019) |
| Treron capellei | large green pigeon | VU | Lowland forest, preferably primary forest (Baptista et al., 2019) |
| Available Range Size | IUCN Status | Species | Habitat Type | Suitable Available Habitat (km2) | Habitat within a PA (%) |
|---|---|---|---|---|---|
| Large | VU | Buceros rhinoceros | Lowland/Montane | 36,400 | 11.1 |
| VU | Ptilocichla leucogrammica | Lowland/Montane | 44,616 | 6.16 | |
| CR | Rhinoplax vigil | Lowland | 50,166 | 5.9 | |
| VU | Rhyticeros undulatus | Lowland/Montane | 41,200 | 18.2 | |
| Medium | VU | Anthracoceros malayanus | Lowland | 25,554 | 9.6 |
| EN | Berenicornis comatus | Lowland/Montane | 12,721 | 13.4 | |
| VU | Caprimulgus concretus | Lowland | 10,431 | 20.5 | |
| VU | Centropus rectunguis | Lowland | 15,404 | 14.2 | |
| VU | Cyornis caerulatus | Lowland | 21,760 | 15.5 | |
| VU | Hydrornis baudii | Lowland | 25,542 | 14.9 | |
| VU | Mulleripicus pulverulentus | Lowland/Montane | 22,292 | 8.6 | |
| VU | Psittacula longicauda | Lowland | 21,394 | 6.7 | |
| VU | Treron capellei | Lowland | 15,702 | 3.4 | |
| Small | VU | Aceros corrugatus | Lowland | 8844 | 8.2 |
| EN | Ciconia stormi | Lowland | 3950 | 11.5 | |
| VU | Leptoptilos javanicus | Lowland | 5957 | 6.5 | |
| VU | Nisaetus nanus | Lowland | 9639 | 16.4 | |
| VU | Spilornis kinabaluensis | Lowland/Montane | 7881 | 30.2 |
| Available Range Size | IUCN Status | Species | Predicted Change in Range Size over the 2010–2050 Window (%) | Predicted Change in the Portion of the Range Found within a PA over the 2010–2050 Window (%) | ||
|---|---|---|---|---|---|---|
| Scenario A1-B | Scenario B1 | Scenario A1-B | Scenario B1 | |||
| Large | VU | Buceros rhinoceros | −10.41 | −7.10 | 0.67 | −2.53 |
| VU | Ptilocichla leucogrammica | −71.22 | −66.20 | 6.16 | 4.14 | |
| CR | Rhinoplax vigil | −32.15 | −37.45 | 2.01 | 0.79 | |
| VU | Rhyticeros undulatus | −12.12 | −3.91 | 1.10 | −2.82 | |
| Medium | VU | Anthracoceros malayanus | 19.21 | 37.74 | −3.59 | −4.11 |
| EN | Berenicornis comatus | 22.26 | 51.09 | −3.80 | −6.50 | |
| VU | Caprimulgus concretus | 292.22 | 207.51 | −11.90 | −13.86 | |
| VU | Centropus rectunguis | 13.49 | 26.09 | −7.44 | −8.44 | |
| VU | Cyornis caerulatus | −5.58 | −25.75 | −0.27 | 0.88 | |
| VU | Hydrornis baudii | −31.47 | −30.46 | 2.39 | 1.09 | |
| VU | Mulleripicus pulverulentus | −6.57 | 2.08 | −2.96 | −4.41 | |
| VU | Psittacula longicauda | 9.32 | 27.34 | −2.97 | −3.88 | |
| VU | Treron capellei | −16.57 | −2.01 | 1.90 | 0.93 | |
| Small | VU | Aceros corrugatus | 67.31 | 39.15 | −2.52 | −3.99 |
| EN | Ciconia stormi | 16.63 | 19.18 | −4.64 | −5.14 | |
| VU | Leptoptilos javanicus | 19.25 | 21.67 | −2.38 | −3.63 | |
| VU | Nisaetus nanus | 79.49 | 84.81 | −10.12 | −10.65 | |
| VU | Spilornis kinabaluensis | −20.94 | −24.49 | −6.73 | −5.01 | |
| Scenario B1 Predictions (Low Emissions) | Scenario A1B Predictions (Medium–High Emissions) | |||||||
|---|---|---|---|---|---|---|---|---|
| Predicted Change in Suitable Habitat between 2010 and 2050 | ||||||||
| F | d.f. | p | effect | F | d.f. | p | Effect | |
| IUCN Category | 0.09 | 1,13 | 0.77 | none detected | 0.00 | 1,13 | 0.97 | none detected |
| Habitat | 0.28 | 1,14 | 0.60 | none detected | 1.10 | 1,14 | 0.31 | none detected |
| Current (2010) area available | 9.35 | 1,15 | 0.008 | Greater loss for large ranged-species | 11.24 | 1,15 | 0.004 | Greater loss for large ranged- species |
| Predicted change in the proportion of the suitable habitat within a currently protected area between 2010 and 2050 | ||||||||
| F | d.f. | p | effect | F | d.f. | p | Effect | |
| IUCN Category | 0.08 | 1,13 | 0.78 | None detected | 0.08 | 1,13 | 0.77 | None detected |
| Habitat | 0.11 | 1,14 | 0.77 | None detected | 1.04 | 1,14 | 0.33 | None detected |
| Current (2010) coverage in PAs | 2.45 | 1,15 | 0.14 | None detected | 4.39 | 1,15 | 0.05 | None detected |
| Current (2010) area available | 8.93 | 1,16 | 0.008 | Most loss at higher initial coverage | 13.24 | 1,16 | 0.002 | Most loss at higher initial coverage |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, M.; Badcock-Scruton, J.; Collins, C.M. What Will Remain? Predicting the Representation in Protected Areas of Suitable Habitat for Endangered Tropical Avifauna in Borneo under a Combined Climate- and Land-Use Change Scenario. Sustainability 2021, 13, 2792. https://0-doi-org.brum.beds.ac.uk/10.3390/su13052792
Singh M, Badcock-Scruton J, Collins CM. What Will Remain? Predicting the Representation in Protected Areas of Suitable Habitat for Endangered Tropical Avifauna in Borneo under a Combined Climate- and Land-Use Change Scenario. Sustainability. 2021; 13(5):2792. https://0-doi-org.brum.beds.ac.uk/10.3390/su13052792
Chicago/Turabian StyleSingh, Minerva, Jessamine Badcock-Scruton, and C. Matilda Collins. 2021. "What Will Remain? Predicting the Representation in Protected Areas of Suitable Habitat for Endangered Tropical Avifauna in Borneo under a Combined Climate- and Land-Use Change Scenario" Sustainability 13, no. 5: 2792. https://0-doi-org.brum.beds.ac.uk/10.3390/su13052792