In this section, the results and influence of dense image matching on point cloud generation, the DSM, and detection of vegetation were discussed.
4.1. Analysis of the Influence of Parameters during Image Matching on the Quality of Point Clouds
The impact of selected control parameters was analysed by comparing the following parameters: the number of filtered 3D points, time per generated DSM post, RMS (Root Mean Square) difference (Z) for the control used and adjusted points, estimated internal height accuracy (DSM), and elapsed time for DSM generation. The vertical accuracy did not change significantly regardless of the selected variant. A very large difference was visible in the density of the cloud, where variant w3 had, on average, 9 times more points than w1, w2, or w5, providing a point cloud density theoretically equal to one point per GSD value compared to other variants with image matching every 2, 3, or 5 pixels. This, of course, had an impact on the time needed to generate the cloud, but this was not a large enough effect to consider important. In this research, it was more important to be able to obtain a higher DSM accuracy. Additionally, in order to compare the impact of selecting the cloud parameters on the resulting DSM simply, images generated from different point clouds were subtracted from each other. This way, images of the differences between generated products were obtained.
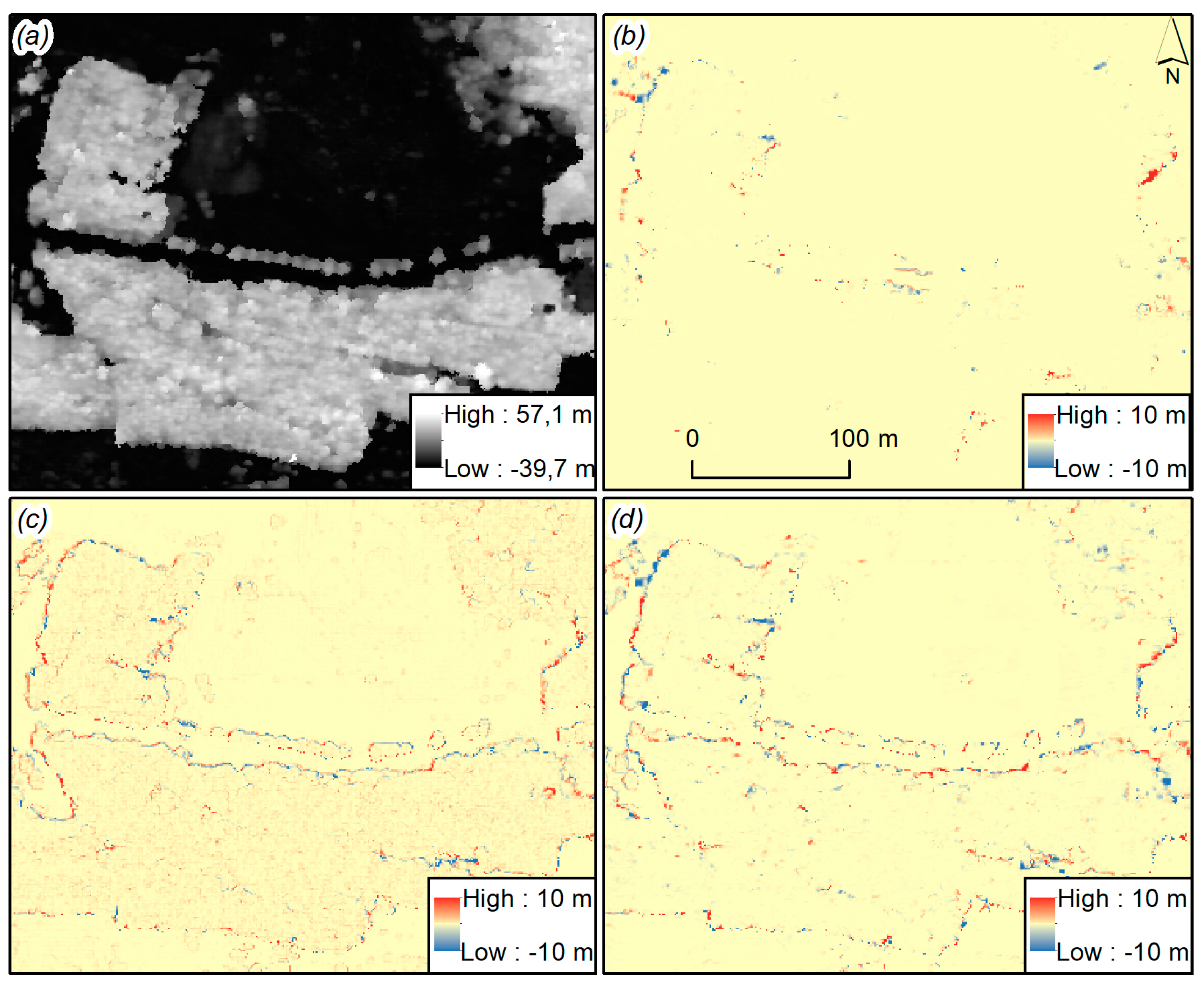
The analysis of these images (
Figure 6) revealed that small differences in the selection of parameters result in significant changes in the resulting cloud obtained and thus in the final rasters.
Changing only the value of parallax (pair w2–w1) resulted mainly in point differences at the boundaries of high objects due to the mapping of an object or lack thereof. Changing the level of smoothing (w5–w1) and cloud density (w3–w1) affects the contours of objects, which differ as a result of smoothing. This effect is caused by stronger interpolation, while in case of cloud condensing, objects grow their perimeters. This increases the probability of tree detection as the cloud is denser and the crown has a larger area. This visual analysis confirms the choice of w3 as the best option when generating a cloud from images in Trimble Inpho software with low smoothing and the densest point cloud generated for points found for each corresponding pixel.
4.2. The Accuracy Assessment of the DSMs Obtained using Dense Image Matching Technique
Table 4 presents the accuracy assessment of the heights obtained based on archival photographs using dense image matching techniques versus stereo measurements. The visual presentation of these results is shown in
Figure 7. The highest accuracy was obtained with the DSM from 2015, which was created from digital colour images of the best radiometric and geometric quality (GSD = 25 cm). These aerial photos were collected during the fully developed growing season; therefore, all species of trees and shrubs were visualised correctly. The standard deviation of the height difference determined by both methods equals 1.27 m (
Table 4). Analysing the differences obtained for particular species, it was noticed that conifer species are relatively small (generally 0.2–0.4 m, standard deviation of the height difference determined by both methods is 0.19 m), while for deciduous species the variation is much higher (standard deviation of the difference in height determined by both methods is 1.45 m, 0.54 after eliminating
Populus balsamifera with 10% crown density). The largest differences in height were observed for individuals with open canopy closure (10%–20% of height). Analysing the data from 2015, it can be noticed that the size and the closure of its crown have a notable impact on the accuracy of the tree heights obtained using the DIM algorithm. In the case of less dense crowns, the contrast between the crown and its surroundings significantly affects the operation of DIM algorithms.
The least accurate results were obtained based on aerial photographs from 2012, 2009, 1996, and 1982. In 2012 and 2009, this was caused by the image acquisition date (March and April), which was at the beginning of the growing season in Poland when most of the trees and shrubs were still leafless or in the early stages of development. This issue limited the effective detection of trees and shrubs. Most values of Δ are positive, which means that these trees in DSM were not contained with their height. Only in the case of coniferous trees was high accuracy obtained for most individuals (problems only occur in the case of pines with irregular crowns). The R2 coefficient for coniferous trees was for 1971: 0.51, 1996: 0.36, 2003: 0.89, 2009: 0.64, 2012: 0.93, and 2015: 0.99.
Aerial photographs from 1996 and 1982 were made in smaller scales, 1:26,000 and 1:32,000, respectively, which lowered the accuracy of the DSMs generated, for both deciduous and coniferous species. The standard deviation of the height differences is 3.57 m for 1982 and 3.88 m for 1996 (
Table 4). In both these terms, significant outliers (up to 13 m) can be observed for a few individuals. This applies particularly to deciduous trees (the biggest errors were obtained for
Acer spp.). For photographs from 1971 and 1982, the poor radiometric quality and visible film damage (
Figure 8) caused significant errors in the elevation model.
Summing up the performed accuracy analysis, the DSMs with the highest accuracy were obtained using images acquired in the fully developed growing season, with a scale of at least 1:13,000 (analogue cameras) or with a GSD less than or equal to 25 cm (digital cameras).
4.3. Analysis of the Effectiveness of the Tree and Shrub Detection using DIM
In order to evaluate the effectiveness of the DIM technique with respect to its use in the analysis of the succession process dynamics, an experiment with multitemporal data, various acquisition terms, and varied image qualities was carried out to determine what features of the examined objects affect the results of their detection. These analyses were conducted for nDSMs obtained using aerial photos from 2015 because the date of their acquisition was the nearest to the date of the field botanical campaigns (2016–2017) and the registration date of ALS data (2016). In addition, aerial photos from 2015 were acquired during the growing season, with GSD = 25 cm, therefore producing the best parameters from the analysed set of archival data.
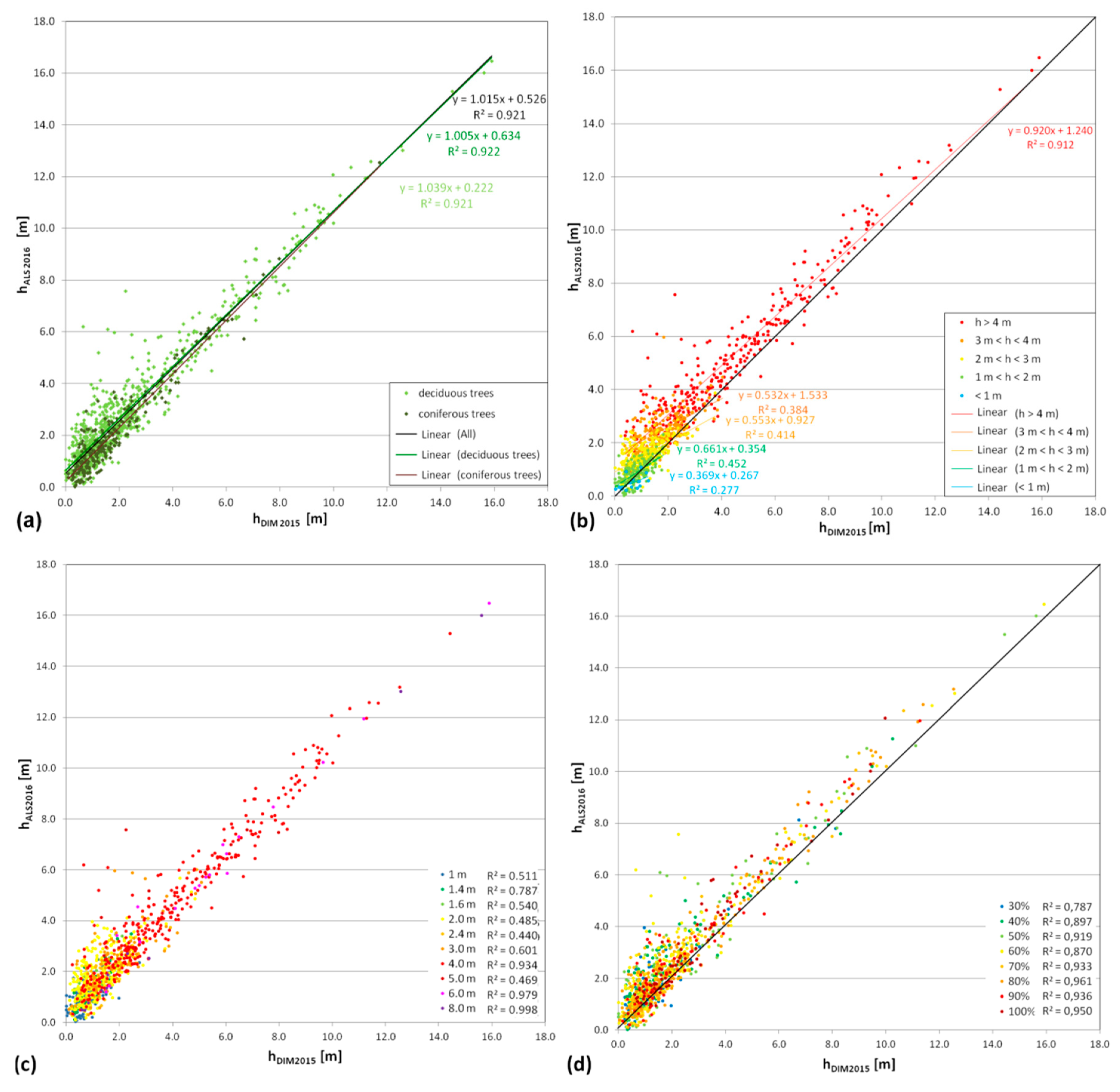
The effectiveness analysis included an impact analysis of the parameters of trees and shrubs (height, crown diameter, crown density) on the tree and shrub detection using the DIM technique. The comparison of the height of trees and shrubs obtained based on ALS data and as a result of DIM techniques depending on the species, height, size, and crown density of the examined trees and shrubs are presented in
Table 5,
Table 6,
Table 7 and
Table 8, and in the sub-figures in
Figure 9. The influence of crown diameter and density (by species) on the accuracy of the height of trees and shrubs using DIM techniques are illustrated in
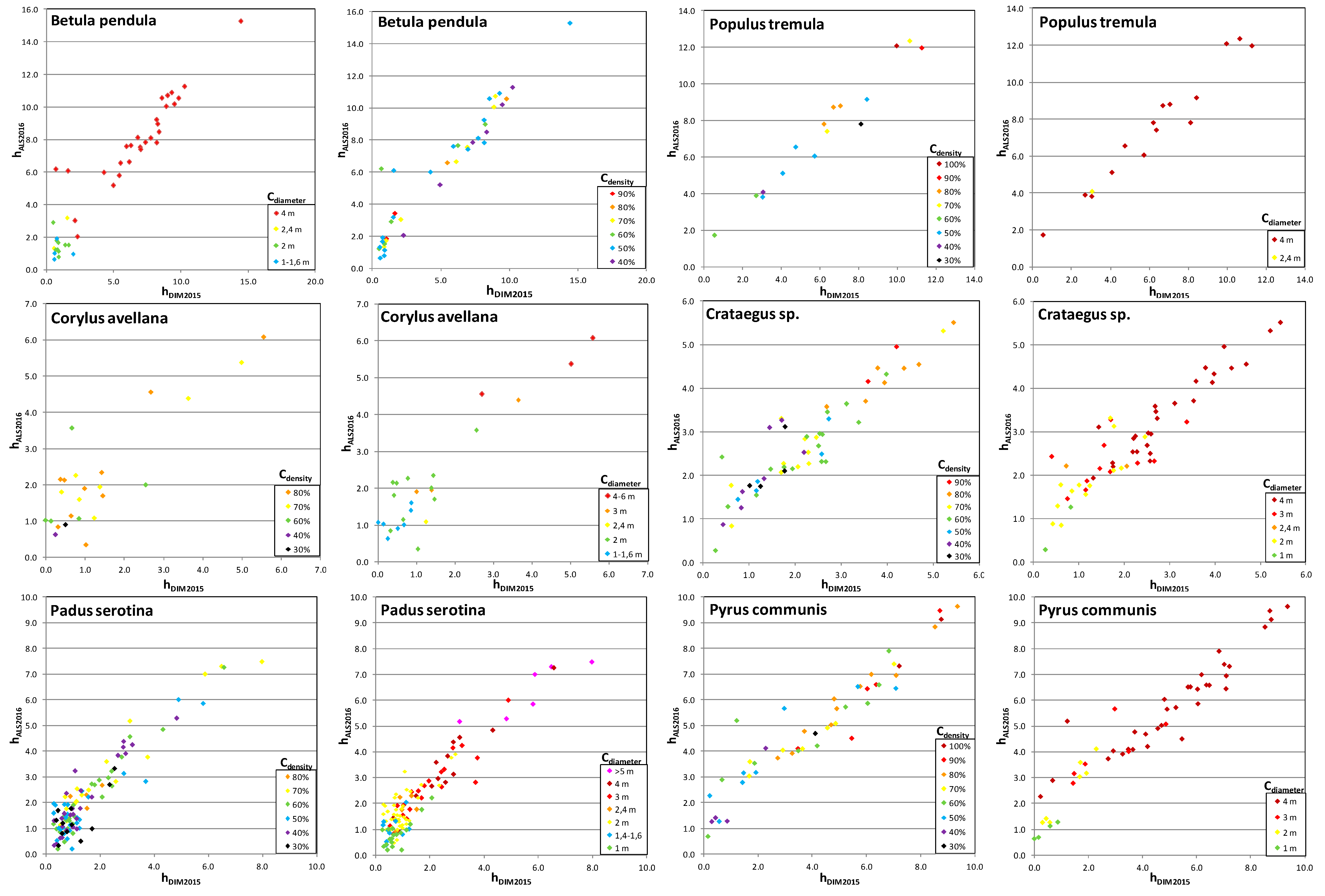
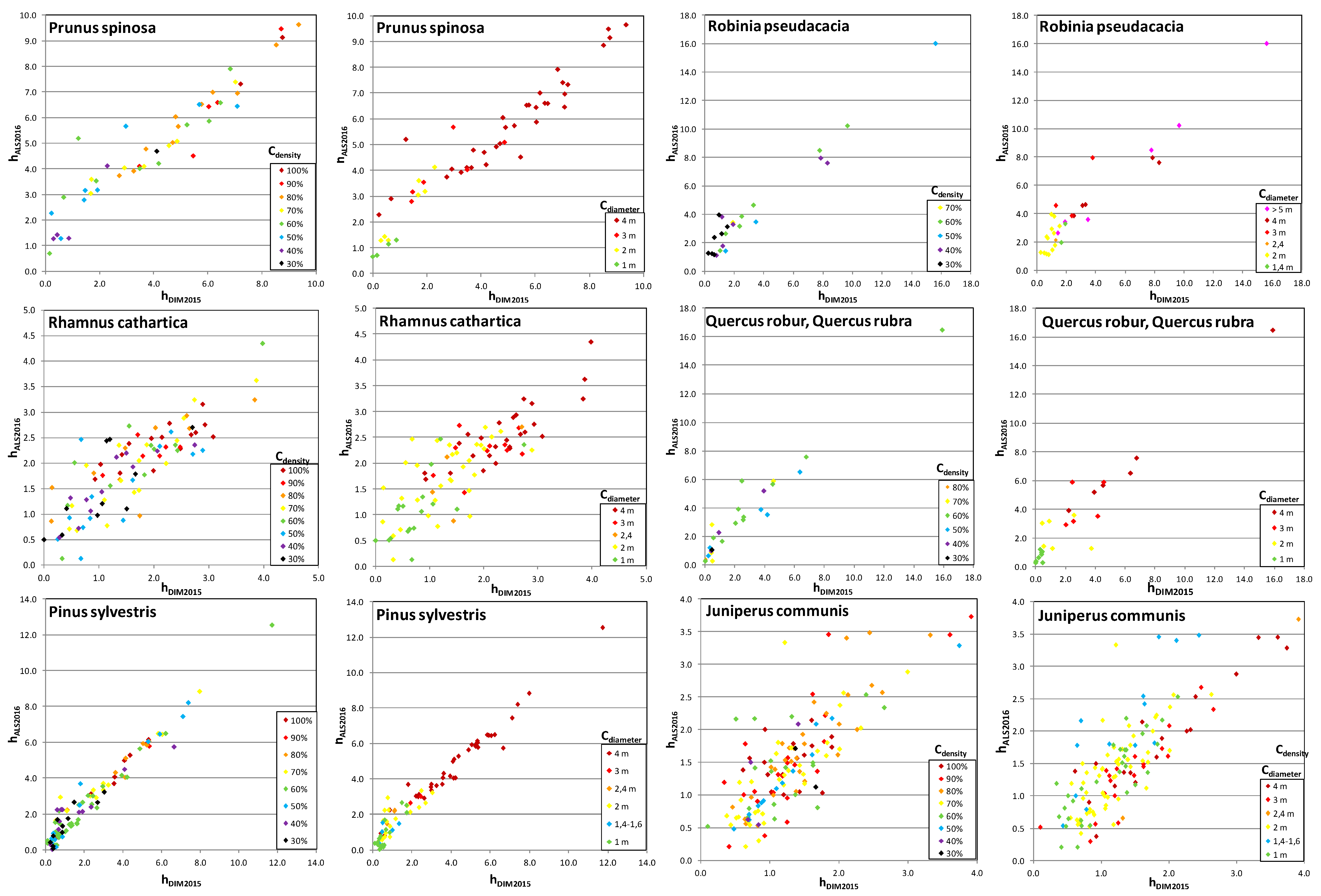
Figure 10.
The height differences indicate that a higher accuracy for determining the height of trees and shrubs is acquired for conifers (
Table 5, μ
|ΔALS-DIM| = 0.43 m). In the case of deciduous trees and shrubs, the mean value of height differences is 0.73 m; it is strongly differentiated (from 0.37 m to 1.64 m, and μ
|ΔALS-DIM| = 0.73 m) and depends on the species and size of the measured object. In general, it was found that tree species, especially those with loose crowns (
Betula pendula, Populus tremula), were characterized by higher values of height differences (
Figure 10). In the case of
Fraxinus sp., Robinia pseudoacacia, and
Quercus rubra, larger differences arise from the specifics of objects present in the study area—some of them were characterized by a small crown density (30%–50%) (
Robinia pseudoacacia) or smaller crown diameter (
Figure 10), moreover
Fraxinus sp. and
Quercus rubra did not occur in large numbers.
Shrubs are characterized by a much smaller error in determining the height than is the case for woody species (the exception here is
Corylus avellana). This applies to both deciduous and coniferous shrubs (
Juniperus communis). The average height differences of shrubs obtained based on LIDAR data and using DIM techniques are 0.49 m for deciduous shrubs and 0.34 m for coniferous, respectively. However, considering their lower heights compared to woody species, the relative error of determining the height is quite high, because it reaches 20%–28% of the height of the shrub (
Table 6). In the case of trees, this error is generally less than 16%.
Analysing the effect of object features on the accuracy of the tree and shrub detection, greater variation in the height difference, μ
|ALS-DIM|, occurs in deciduous trees and increases with decreasing tree height and reducing crown density (
Figure 9a–d). Coniferous trees are generally characterized by a better height mapping than is the case for deciduous trees (
Table 5,
Figure 9a). It can also be noticed that the larger the tree crown (diameter over 3–4 m), the better the height mapping using DIM techniques (
Figure 9c). The graphs in
Figure 9 show outliers that represent deciduous trees with a height of approx. 5–6 m. These are
Betula pendula, Fraxinus sp., and
Quercus rubra with tree crowns with 3–4 meter diameters and 50%–60% density (
Figure 9c,d).
Betula pendula is common in the analysed area. During the secondary succession process,
Betula pendula enters abandoned fields and unused grasslands as one of the first tree species. Most individuals have well-developed, slender crowns. Over a dozen-year-olds, medium height individuals of
Betula pendula dominate, which is associated with the massive abandonment of cultivation and grazing over the last 20 years.
Fraxinus sp. and Quercus rubra are rarely found in the analysed area. Fraxinus sp. is usually planted as a roadside tree, Quercus rubra grows spontaneously in abandoned fields and grasslands, and sometimes is planted in forests. Most individuals of both Fraxinus sp. and Quercus rubra growing in abandoned fields and grasslands have well-developed, spherical crowns. But individuals of Fraxinus sp. are usually adult, and individuals of Quercus rubra are mostly young.
Analysis of the similarity degree of heights obtained based on LIDAR data from 2016 and using DIM algorithms (
Figure 9c), depending on the size of crowns, showed that trees and shrubs with a crown size above 4 m are characterized by the highest compatibility of both heights (R
2 above 0.9, with the exception of 5 m crown diameter, which is due to the small sample number of such objects).
Analysing the influence of species on the results, for smaller shrubs and brushwood (with a crown diameter less than 2.4 m (
Table 7) and height less than 2 m (
Table 6)), there was no influence of type, species, or other traits on the value of μ
|ALS-DIM|. Both coniferous and deciduous species are at a similar level. For larger trees, μ
|ALS-DIM| values are reduced for coniferous species. No influence was observed for the density of crowns of trees and shrubs on the obtained mean differences in height, μ
|ALS-DIM (
Table 8). Because the analysed area has physiognomically different grades with different heights and crowns, an analysis was carried out with differences in height depending on: heights and crown diameters (
Table 9), heights and crown density (
Table 10), crown diameter and density (
Table 11).
The results summarized in
Table 9 indicate that the mean height differences, μ
| ALS-DIM |, for trees and shrubs from a certain height range slightly increase when the crown diameter of a tree or bush decreases. Only in the case of crowns with a larger diameter is there no such a clear tendency, which may result from other features, such as species or shortening of the crown.
Figure 10 clearly shows that shrub species (especially
Rhamnus cathartica,
Juniperus communis,
Crataegus spp.) are characterized by large differences in height depending on the diameter and density of the crown. For
Crataegus spp., the smallest height deviations were noted for higher crown densities. The largest deviations in height for
Juniperus communis were from shrubs with a smaller crown diameter (<1.6 m). The differences observed in height obtained using DIM vs. ALS results mainly from the specificity of these species and their habit.
For smaller shrubs and trees (up to a height of 2 m), there is no significant relationship between crown density and the value of μ
|ALS-DIM| (
Table 10). Clear trends are visible only for trees and shrubs higher than 2 m. In most cases, for lower crown density (below 60%), differences in height with respect to ALS data are higher, but two exceptions were found here. For example, high values for trees with a height over 10 m and 100% crowns are for individuals with narrow crowns, which affects the results of the DIM algorithm.
A similar situation is seen for the analysis of the relationship between the crown diameter and crown density (
Table 11). In the case of small trees and shrubs with crown diameters below 1.6 m, the influence of the crown density on the correctness of vegetation height cannot be noticed. For larger individuals with less than 60% crown density, generally higher differences with the ALS data in the value of μ
|ALS-DIM| were found. These results were confirmed with R
2 determination coefficients obtained to determine the degree of similarity of the heights based on the ALS data from 2016 and as a result of the DIM algorithms (
Figure 9d). Lower coefficient values were obtained for trees and shrubs with crown densification up to 60%. The Pearson correlation coefficient of trees heights obtained using the DIM and ALS (for all analysed trees and shrubs) is 0.96 (R
2 = 0.92).

 ,
, 












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}