
Integrate the Canopy SIF and Its Derived Structural and Physiological Components for Wheat Stripe Rust Stress Monitoring

Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Experimental Areas
2.2. Data Acquisition
2.2.1. Canopy Spectral Measurements
2.2.2. Severity Level Survey
2.2.3. SIFcanopy Retrieval Method
2.2.4. Estimation of NIRVP and SIFtot
3. Results and Analysis
3.1. Effects of NIRVP and SIFtot on the Variations in SIFcanopy under Different Disease Severities
3.2. The Relationships between SIFcanopy, NIRVP, SIFtot, and SL of Different Disease Severities
3.3. Model Accuracy Test
4. Discussion
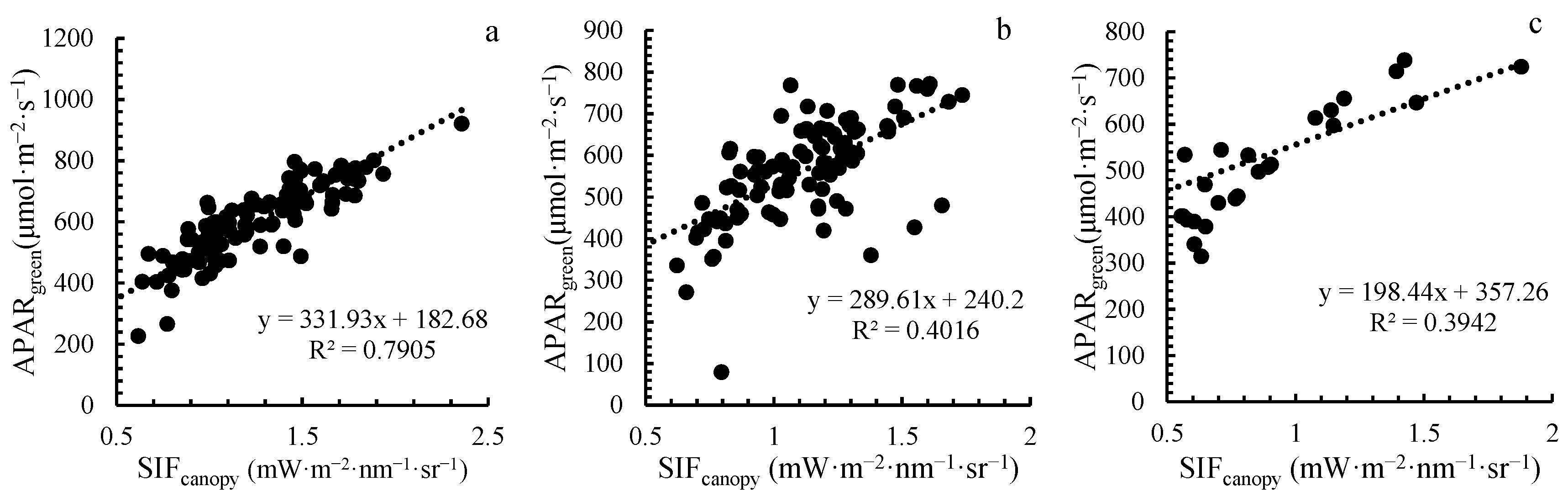
4.1. Response Characteristics between SIFcanopy and APARgreen under Stripe Rust Stress
4.2. Analysis of the Influence of Uncertainty Factors on SIFcanopy and fAPARgreen
4.3. Responses of SIFcanopy and SL to Canopy Structure and Plant Physiological Indicators under Disease Stress
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, Z.; Wang, S.; Qiu, B.; Song, L.; Zhang, Y. Retrieval of sun-induced chlorophyll fluorescence and advancements in carbon cycle application. J. Remote Sens. 2019, 23, 16. [Google Scholar]
- Porcar-Castell, A.; Tyystjärvi, E.; Atherton, J.; Van der Tol, C.; Flexas, J.; Pfündel, E.E.; Moreno, J.; Frankenberg, C.; Berry, J.A. Linking chlorophyll a fluorescence to photosynthesis for remote sensing applications: Mechanisms and challenges. J. Exp. Bot. 2014, 65, 4065–4095. [Google Scholar] [CrossRef]
- Yang, P.; Liu, Z.; Ni, Z.; Wang, Q. Remote Sensing of Chlorophyll Fluorescence at Airborne Level Based on Unmanned Airship Platform and Hyperspectral Sensor. Spectrosc. Spectr. Anal. 2013, 33, 3101–3105. [Google Scholar] [CrossRef]
- Huang, C.; Zhang, L.; Wang, S.; Qiao, N. Automated observation of diurnal solar-induced chlorophyll fluorescence for better understanding of crop photosynthesis. In Proceedings of the Agu Fall Meeting Abstracts, San Francisco, CA, USA, 12–16 December 2016; p. B51B-0406. [Google Scholar]
- Somkuti, P.; Bsch, H.; Liang, F.; Palmer, P.I.; Quaife, T. A new space-borne perspective of crop productivity variations over the US Corn Belt. Agric. For. Meteorol. 2020, 281, 107826. [Google Scholar] [CrossRef]
- Zhao, Y.; Jing, X.; Huang, W.; Dong, Y.; Li, C. Comparison of Sun-Induced Chlorophyll Fluorescence and Reflectance Data on Estimating Severity of Wheat Stripe Rust. Spectrosc. Spectr. Anal. 2019, 39, 2739. [Google Scholar]
- Jing, X.; Lv, X.; Zhang, C.; BAI, Z.-F. Early Detection of Winter Wheat Stripe Rust Based on SIF-PLS Model. Trans. Chin. Soc. Agric. Mach. 2020, 51, 191–197. [Google Scholar] [CrossRef]
- Hernandez-Clemente; North, P.R.J.; Hornero, A.; Zarco-Tejada, P.J. Assessing the effects of forest health on sun-induced chlorophyll fluorescence using the FluorFLIGHT 3-D radiative transfer model to account for forest structure. Remote Sens. Environ. 2017, 193, 165–179. [Google Scholar] [CrossRef] [Green Version]
- Saleem, M.; Atta, B.M.; Ali, Z.; Bilal, M. Laser induced Fluorescence spectroscopy for early disease detection in grapefruit plants. Photochem. Photobiol. Sci. 2020, 19, 713–721. [Google Scholar] [CrossRef]
- Song, Y.; Wang, J.; Wang, L.J.R.S. Satellite solar-induced chlorophyll fluorescence reveals heat stress impacts on wheat yield in India. Remote Sens. 2020, 12, 3277. [Google Scholar] [CrossRef]
- Xian, C.; He, L.; He, Z.; Xue, D.; Li, Z. Assessing the Response of Satellite Solar-Induced Chlorophyll Fluorescence and NDVI to Impacts of Heat Waves on Winter Wheat in the North China Plain. Adv. Meteorol. 2020, 2020, 1–14. [Google Scholar] [CrossRef]
- Sun, Y.; Fu, R.; Dickinson, R.; Joiner, J.; Frankenberg, C.; Gu, L.; Xia, Y.; Fernando, N. Drought onset mechanisms revealed by satellite solar-induced chlorophyll fluorescence: Insights from two contrasting extreme events. J. Geophys. Res. Biogeosci. 2016, 120, 2427–2440. [Google Scholar] [CrossRef]
- Xu, S.; Atherton, J.; Riikonen, A.; Zhang, C.; Oivukkamäki, J.; MacArthur, A.; Honkavaara, E.; Hakala, T.; Koivumäki, N.; Liu, Z.; et al. Structural and photosynthetic dynamics mediate the response of SIF to water stress in a potato crop. Remote Sens. Environ. 2021, 263, 112555. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, X.; Wolf, S.; Wu, J.; Wu, X.; Gioli, B.; Wohlfahrt, G.; Cescatti, A.; Christiaan, V.; Zhou, S. Spatio-Temporal Convergence of Maximum Daily Light-Use Efficiency Based on Radiation Absorption by Canopy Chlorophyll. Geophys. Res. Lett. 2018, 45, 3508–3519. [Google Scholar] [CrossRef]
- Van der Tol, C.; Berry, J.A.; Campbell, P.; Rascher, U. Models of fluorescence and photosynthesis for interpreting measurements of solar-induced chlorophyll fluorescence. J. Geophys. Res.-Biogeosci. 2014, 119, 2312–2327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, P.; Christiaan, V. Linking canopy scattering of far-red sun-induced chlorophyll fluorescence with reflectance. Remote Sens. Environ. 2018, 209, 456–467. [Google Scholar] [CrossRef]
- Köhler, P.; Guanter, L.; Kobayashi, H.; Walther, S.; Yang, W. Assessing the potential of sun-induced fluorescence and the canopy scattering coefficient to track large-scale vegetation dynamics in Amazon forests. Remote Sens. Environ. 2018, 204, 769–785. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Luis, G.; Liu, L.; Alexander, D.; Zbyněk, M.; Uwe, R.; Peng, D.; Du, S.; Jean-Philippe, G.E. Downscaling of solar-induced chlorophyll fluorescence from canopy level to photosystem level using a random forest model. Remote Sens. Environ. 2018, 231, 110772. [Google Scholar] [CrossRef]
- Zeng, Y.; Chen, M.; Hao, D.; Damm, A.; Badgley, G.; Rascher, U.; Johnson, J.E.; Dechant, B.; Siegmann, B.; Ryu, Y. Combining near-infrared radiance of vegetation and fluorescence spectroscopy to detect effects of abiotic changes and stresses. Remote Sens. Environ. 2022, 270, 112856. [Google Scholar] [CrossRef]
- Zeng, Y.; Badgl Ey, G.; Dechant, B.; Ryu, Y.; Berry, J.A. A practical approach for estimating the escape ratio of near-infrared solar-induced chlorophyll fluorescence. Remote Sens. Environ. 2019, 232, 111209. [Google Scholar] [CrossRef] [Green Version]
- Guanter, L.; Zhang, Y.; Jung, M.; Joiner, J.; Voigt, M.; Berry, J.A.; Frankenberg, C.; Huete, A.R.; Zarco-Tejada, P.; Lee, J.-E.; et al. Global and time-resolved monitoring of crop photosynthesis with chlorophyll fluorescence. Proc. Natl. Acad. Sci. USA 2014, 111, E1327–E1333. [Google Scholar] [CrossRef] [Green Version]
- Dechant, B.; Ryu, Y.; Badgley, G.; Zeng, Y.; Moya, I. Canopy structure explains the relationship between photosynthesis and sun-induced chlorophyll fluorescence in crops. Remote Sens. Environ. 2020, 241, 111733. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Guan, K.; Peng, B.; Chen, M.; Miao, G. Satellite footprint data from OCO-2 and TROPOMI reveal significant spatio-temporal and inter-vegetation type variabilities of solar-induced fluorescence yield in the U.S. Midwest. Remote Sens. Environ. 2020, 241, 111728. [Google Scholar] [CrossRef]
- Zhang, J.; Pu, R.; Huang, W.; Yuan, L.; Luo, J.; Wang, J. Using in-situ hyperspectral data for detecting and discriminating yellow rust disease from nutrient stresses. Field Crops Res. 2012, 134, 165–174. [Google Scholar] [CrossRef]
- Huang, W.; Shi, Y.; Dong, Y.; Ye, H.; Wu, M.; Cui, B.; Liu, L. Progress and prospects of crop diseases and pests monitoring by remote sensing. Smart Agric. 2019, 1, 1–11. [Google Scholar] [CrossRef]
- Jing, X.; Zou, Q.; Bai, Z.-F.; Huang, W.-J. Research progress of crop diseases monitoring based on reflectance and chlorophyll fluorescence data. Acta Agron. Sin. 2021, 47, 2067–2079. [Google Scholar] [CrossRef]
- General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China; China National Standardization Management Committee. Technical Specifications for Forecasting Wheat Stripe Rust. 2011. Available online: https://kns.cnki.net/kcms/detail/detail.aspx?FileName=SCSF00036649&DbName=SCSF (accessed on 29 September 2011).
- Yuan, L.; Zhang, J.; Nie, C.; Wei, L.; Yang, G.; Wang, J. Selection of Spectral Channels for Satellite Sensors in Monitoring Yellow Rust Disease of Winter Wheat. Intell. Autom. Soft Comput. 2013, 19, 501–511. [Google Scholar] [CrossRef]
- Maier, S.W.; Günther, K.; Stellmes, M. Sun-Induced Fluorescence: A New Tool for Precision Farming. Digit. Imaging Spectr. Tech. Appl. Precis. Agric. Crop Physiol. 2004, 66, 207–222. [Google Scholar] [CrossRef]
- Liu, X.; Liu, L. Improving Chlorophyll Fluorescence Retrieval Using Reflectance Reconstruction Based on Principal Components Analysis. IEEE Geosci. Remote Sens. Lett. 2015, 12, 1645–1649. [Google Scholar] [CrossRef]
- Liu, X.; Liu, L. Assessing Band Sensitivity to Atmospheric Radiation Transferfor Space-Based Retrieval of Solar-Induced Chlorophyll Fluorescence. Remote Sens. 2014, 6, 10656–10675. [Google Scholar] [CrossRef] [Green Version]
- Gitelson, A.A. Wide Dynamic Range Vegetation Index for Remote Quantification of Biophysical Characteristics of Vegetation. J. Plant Physiol. 2004, 161, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Viña, A.; Gitelson, A.A. New developments in the remote estimation of the fraction of absorbed photosynthetically active radiation in crops. Geophys. Res. Lett. 2005, 32, 17. [Google Scholar] [CrossRef] [Green Version]
- Gitelson, A.A. Remote estimation of fraction of radiation absorbed by photosynthetically active vegetation: Generic algorithm for maize and soybean. Remote Sens. Lett. 2019, 10, 283–291. [Google Scholar] [CrossRef]
- Kimm, H.; Guan, K.; Burroughs, C.H.; Peng, B.; Wu, G. Quantifying high-temperature stress on soybean canopy photosynthesis: The unique role of sun-induced chlorophyll fluorescence. Glob. Change Biol. 2021, 27, 2403–2415. [Google Scholar] [CrossRef] [PubMed]
- Dechant, B.; Ryu, Y.; Badgley, G.; Khler, P.; Berry, J. NIRvP: A robust structural proxy for sun-induced chlorophyll fluorescence and photosynthesis across scales. Remote Sens. Environ. 2022, 268, 112763. [Google Scholar] [CrossRef]
- Shi, Y.; Huang, W.; Luo, J.; Huang, L.; Zhou, X. Detection and discrimination of pests and diseases in winter wheat based on spectral indices and kernel discriminant analysis. Comput. Electron. Agric. 2017, 141, 171–180. [Google Scholar] [CrossRef]
- Chang, C.Y.; Wen, J.; Han, J.; Kira, O.; LeVonne, J.; Melkonian, J.; Sun, Y. Unpacking the drivers of diurnal dynamics of sun-induced chlorophyll fluorescence (SIF): Canopy structure, plant physiology, instrument configuration and retrieval methods. Remote Sens. Environ. 2021, 265, 112672. [Google Scholar] [CrossRef]
- Mahlein, A.K.; Steiner, U.; Hillnhütter, C.; Dehne, H.W.; Oerke, E.C. Hyperspectral imaging for small-scale analysis of symptoms caused by different sugar beet diseases. Plant Methods 2012, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- Martinelli, F.; Scalenghe, R.; Davino, S.; Panno, S.; Scuderi, G.; Ruisi, P.; Villa, P.; Stroppiana, D.; Boschetti, M.; Goulart, L.R. Advanced methods of plant disease detection. A review. Agron. Sustain. Dev. 2015, 35, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Calderón, R.; Navas-Cortés, J.; Lucena, C.; Zarco-Tejada, P.J. High-resolution airborne hyperspectral and thermal imagery for early detection of Verticillium wilt of olive using fluorescence, temperature and narrow-band spectral indices. Remote Sens. Environ. 2013, 139, 231–245. [Google Scholar] [CrossRef]
- Wang, L.; Liu, J.; Yang, F.; Yang, L.; Yao, B.; Gao, J. Winter wheat stripe rust remote sensing monitoring based on GF-1/WFV data. Trans. Chin. Soc. Agric. Eng. 2017, 33, 153–160. [Google Scholar] [CrossRef]
- Esmail, S.M.; Omara, R.I.; Abdelaal, K.; Hafez, Y.M. Histological and biochemical aspects of compatible and incompatible wheat-Puccinia striiformis interactions. Physiol. Mol. Plant Pathol. 2018, 106, 120–128. [Google Scholar] [CrossRef]
- Zhang, J. Methods for Information Extraction of Wheat Disease Based on Multi-Source Remote Sensing Data. Ph.D. Thesis, ZheJiang University, Hangzhou, China, 2012. [Google Scholar]
- Chen, W.; Wellings, C.; Chen, X.; Kang, Z.; Liu, T. Wheat stripe (yellow) rust caused by Puccinia striiformis f. sp.tritici. Mol. Plant Pathol 2014, 15, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Miao, G.; Guan, K.; Suyker, A.E.; Yang, X.; Arkebauer, T.J.; Walter-Shea, E.A.; Kimm, H.; Hmimina, G.Y.; Gamon, J.A.; Franz, T.E. Varying Contributions of Drivers to the Relationship Between Canopy Photosynthesis and Far-Red Sun-Induced Fluorescence for Two Maize Sites at Different Temporal Scales. J. Geophys. Res. Biogeosci. 2020, 125, e2019JG005051. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.; Ryu, Y.; Dechant, B.; Berry, J.A.; Hwang, Y.; Jiang, C.; Kang, M.; Kim, J.; Kimm, H.; Kornfeld, A.; et al. Sun-induced chlorophyll fluorescence is more strongly related to absorbed light than to photosynthesis at half-hourly resolution in a rice paddy. Remote Sens. Environ. 2018, 216, 658–673. [Google Scholar] [CrossRef]
- Shanshan, D.; Liangyun, L.; Xinjie, L.; Jiaochan, H. Response of Canopy Solar-Induced Chlorophyll Fluorescence to the Absorbed Photosynthetically Active Radiation Absorbed by Chlorophyll. Remote Sens. 2017, 9, 911. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Hou, X.; Wang, M.; Wang, L.; Liu, F. Study on Relationship Between Photosynthetic Rate and Hyperspectral Indexes of Wheat Under Stripe Rust Stress. Spectrosc. Spectr. Anal. 2022, 42, 940–946. [Google Scholar]
- Ashraf, M.; Harris, P. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Christiaan, V.; Rossini, M.; Cogliati, S.; Verhoef, W.; Colombo, R.; Rascher, U.; Mohammed, G. A model and measurement comparison of diurnal cycles of sun-induced chlorophyll fluorescence of crops. Remote Sens. Environ. 2016, 186, 663–677. [Google Scholar] [CrossRef]
- Micol, R.; Michele, M.; Marco, C.; Sergio, C.; Tommaso, J.; Cinzia, P.; Uwe, R.; Christiaan, V.D.T.; Roberto, C. Analysis of Red and Far-Red Sun-Induced Chlorophyll Fluorescence and Their Ratio in Different Canopies Based on Observed and Modeled Data. Remote Sens. 2016, 8, 412. [Google Scholar] [CrossRef] [Green Version]
- Atherton, J.; Nichol, C.J.; Porcar-Castell, A. Using spectral chlorophyll fluorescence and the photochemical reflectance index to predict physiological dynamics. Remote Sens. Environ. 2016, 176, 17–30. [Google Scholar] [CrossRef]
- Liu, L.; Liu, X.; Wang, Z.; Zhang, B. Measurement and Analysis of Bidirectional SIF Emissions in Wheat Canopies. IEEE Trans. Geosci. Remote Sens. 2016, 54, 2640–2651. [Google Scholar] [CrossRef]
- Fournier, A.; Goulas, Y.; Daumard, F.; Ounis, A.; Champagne, S.; Moya, I. Effects of vegetation directional reflectance on sun-induced fluorescence retrieval in the oxygen absorption bands. In Proceedings of the 5th International Workshop on Remote Sensing of Vegetation Fluorescence, Paris, France, 22–24 April 2014. [Google Scholar]
- Damm, A.; Guanter, L.; Verhoef, W.; Schläpfer, D.; Garbari, S.; Schaepman, M.E. Impact of varying irradiance on vegetation indices and chlorophyll fluorescence derived from spectroscopy data. Remote Sens. Environ. 2015, 156, 202–215. [Google Scholar] [CrossRef]
- Cartwright, D.; Russell, G. Development of Puccinia striiformis in a susceptible winter wheat variety. Trans. Br. Mycol. Soc. 1981, 76, 197–204. [Google Scholar] [CrossRef]
- Bushnell, W.R. Structural and Physiological Alterations in Susceptible Host Tissue. Cereal Rusts 1984, 1, 477–507. [Google Scholar]
- Liu, Q. Remote Sensing Monitoring and Duplex Real-Time PCR Quantitative Determination of Wheat Stripe Rust in Latent Period; China Agricultural University: Beijing, China, 2016. [Google Scholar]
- Chang, Q.; Liu, J.; Wang, Q.; Han, L.; Liu, J.; Li, M.; Huang, L.; Yang, J.; Kang, Z. The effect of Puccinia striiformis f. sp. tritici on the levels of water-soluble carbohydrates and the photosynthetic rate in wheat leaves. Physiol. Mol. Plant Pathol. 2013, 84, 131–137. [Google Scholar] [CrossRef]
- Kimm, H.; Guan, K.; Jiang, C. A physiological signal derived from sun-induced chlorophyll fluorescence quantifies crop physiological response to environmental stresses in the U.S. Corn Belt. Environ. Res. Lett. 2021, 16, 124051. [Google Scholar] [CrossRef]
- Zhang, J.; Huang, Y.; Pu, R.; Gonzalez-Moreno, P.; Yuan, L.; Wu, K.; Huang, W. Monitoring plant diseases and pests through remote sensing technology: A review. Comput. Electron. Agric. 2019, 165, 104943. [Google Scholar] [CrossRef]
- Ma, Z. Epidemiology of Plant Diseases; Science Press: Beijing, China, 2010. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable Name | Mild Condition (n = 72) | Moderate Condition (n = 69) | Severe Condition (n = 20) | |||
|---|---|---|---|---|---|---|
| R2 | RMSE | R2 | RMSE | R2 | RMSE | |
| SIFcanopy | 0.50 | 0.037 | 0.46 * | 0.046 | 0.34 | 0.101 |
| SIFtot | 0.53 * | 0.036 | 0.31 | 0.051 | 0.28 | 0.109 |
| NIRVP | 0.28 | 0.044 | 0.23 | 0.055 | 0.59 * | 0.068 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jing, X.; Li, B.; Ye, Q.; Zou, Q.; Yan, J.; Du, K. Integrate the Canopy SIF and Its Derived Structural and Physiological Components for Wheat Stripe Rust Stress Monitoring. Remote Sens. 2022, 14, 3427. https://0-doi-org.brum.beds.ac.uk/10.3390/rs14143427
Jing X, Li B, Ye Q, Zou Q, Yan J, Du K. Integrate the Canopy SIF and Its Derived Structural and Physiological Components for Wheat Stripe Rust Stress Monitoring. Remote Sensing. 2022; 14(14):3427. https://0-doi-org.brum.beds.ac.uk/10.3390/rs14143427
Chicago/Turabian StyleJing, Xia, Bingyu Li, Qixing Ye, Qin Zou, Jumei Yan, and Kaiqi Du. 2022. "Integrate the Canopy SIF and Its Derived Structural and Physiological Components for Wheat Stripe Rust Stress Monitoring" Remote Sensing 14, no. 14: 3427. https://0-doi-org.brum.beds.ac.uk/10.3390/rs14143427