Twelve-Week Protocatechuic Acid Administration Improves Insulin-Induced and Insulin-Like Growth Factor-1-Induced Vasorelaxation and Antioxidant Activities in Aging Spontaneously Hypertensive Rats

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Resting Blood Pressure and Heart Rate
2.3. Vasoreactivity Experiments
2.4. Blood Collection and Biochemical Analysis
2.5. Insulin Resistance Determination
2.6. Western Immunoblotting
2.7. Statistical Analysis
3. Results
3.1. General Characteristics
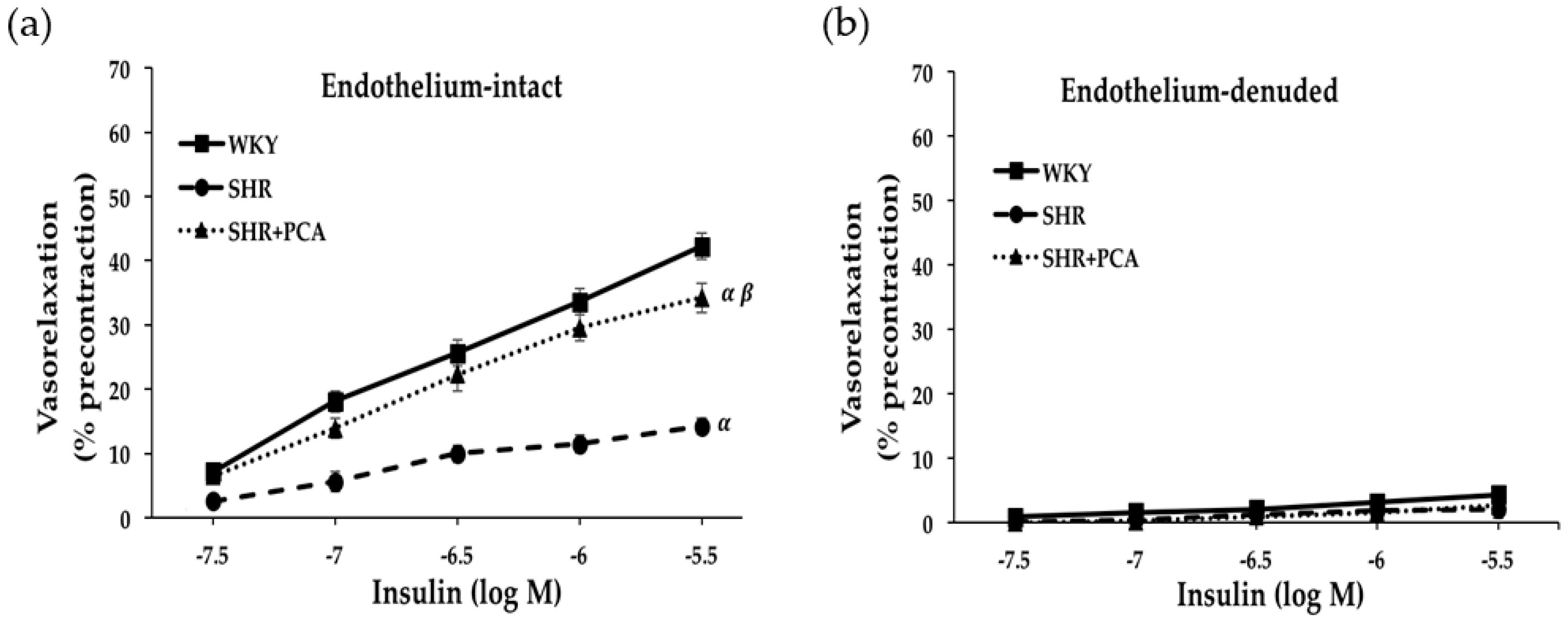
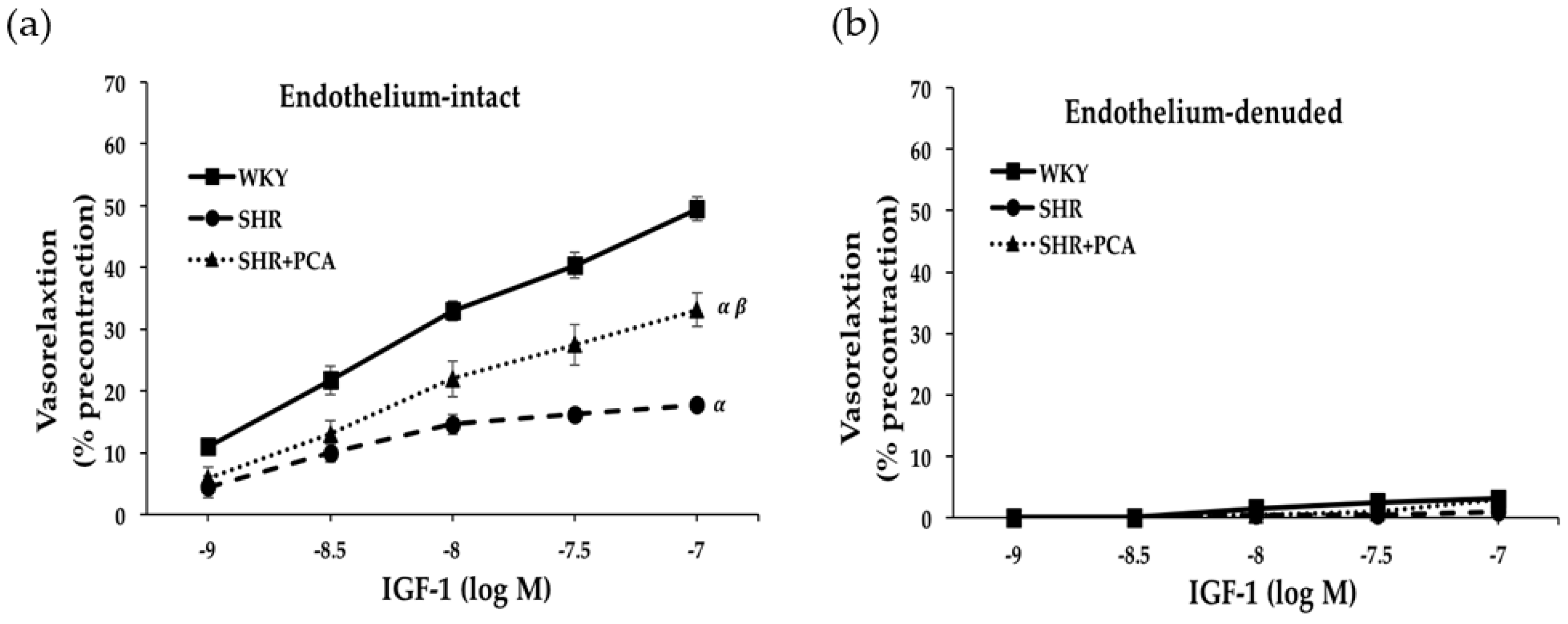
3.2. Insulin-Induced and IGF-1-Induced Vasorelaxation in Aortas
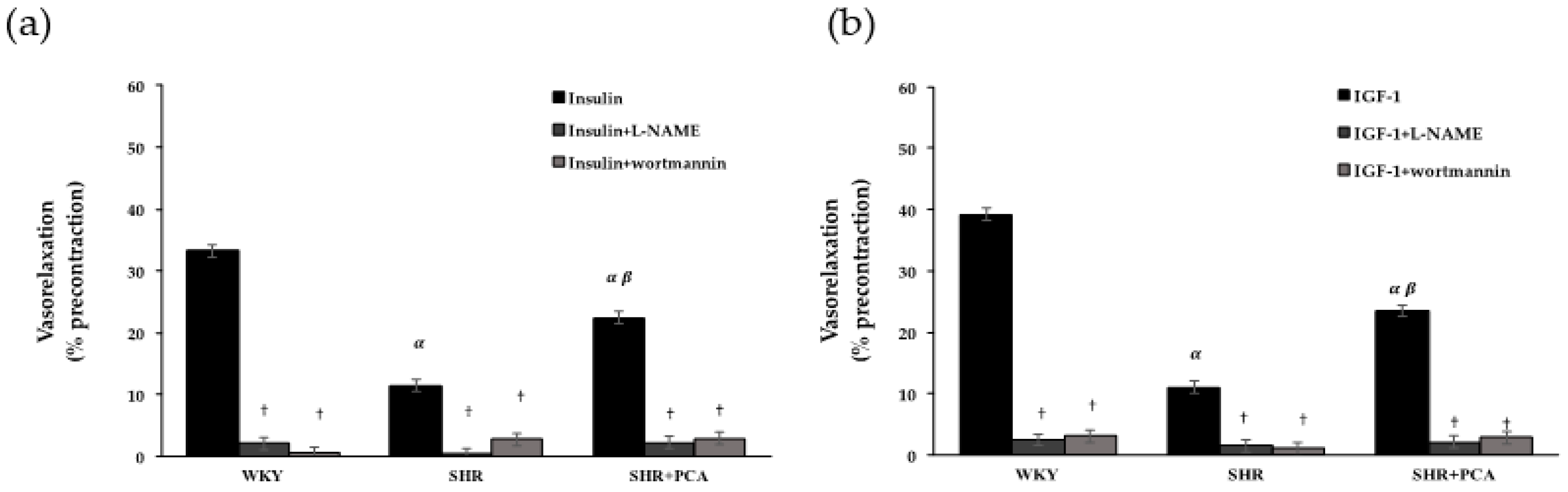
3.3. Roles of PI3K and NOS in Insulin-Induced and IGF-1-Induced Vasorelaxation
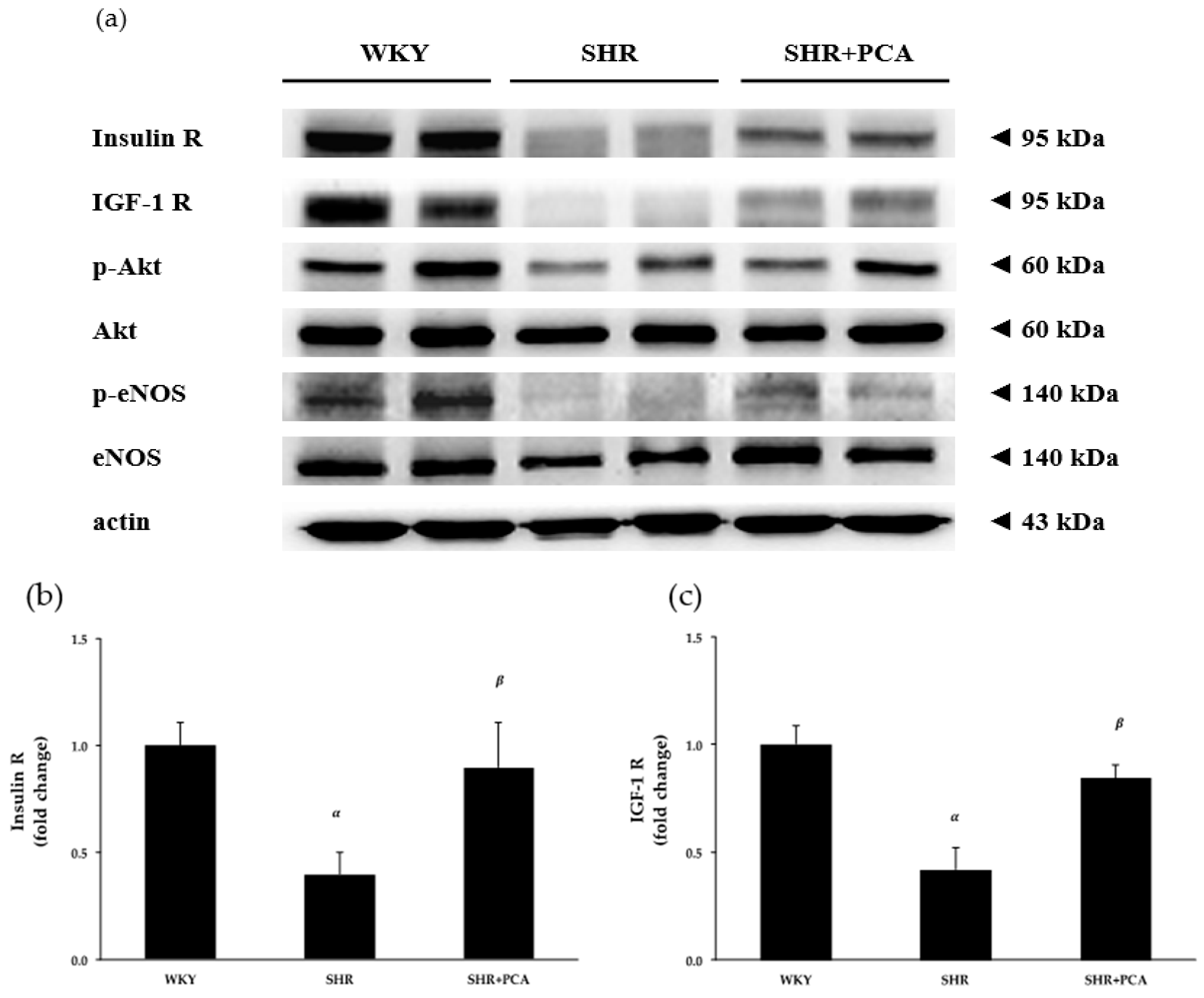
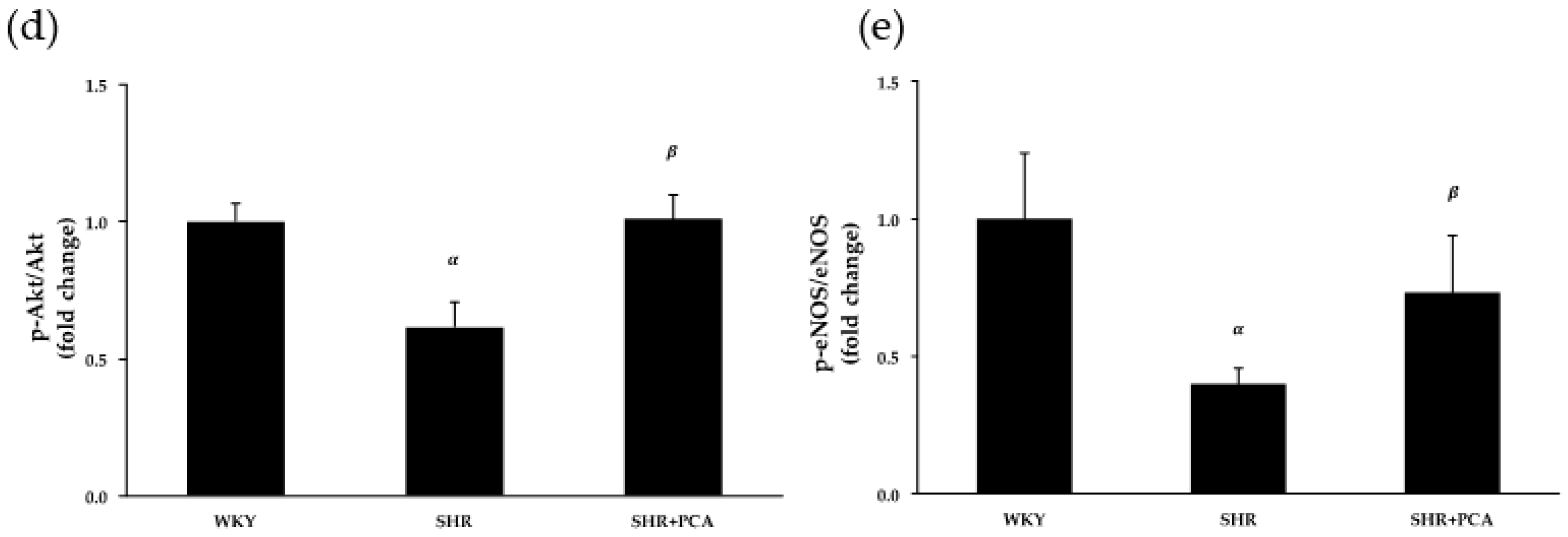
3.4. Aortic Protein Expression
3.5. Serum Nitrate/Nitrite Concentration
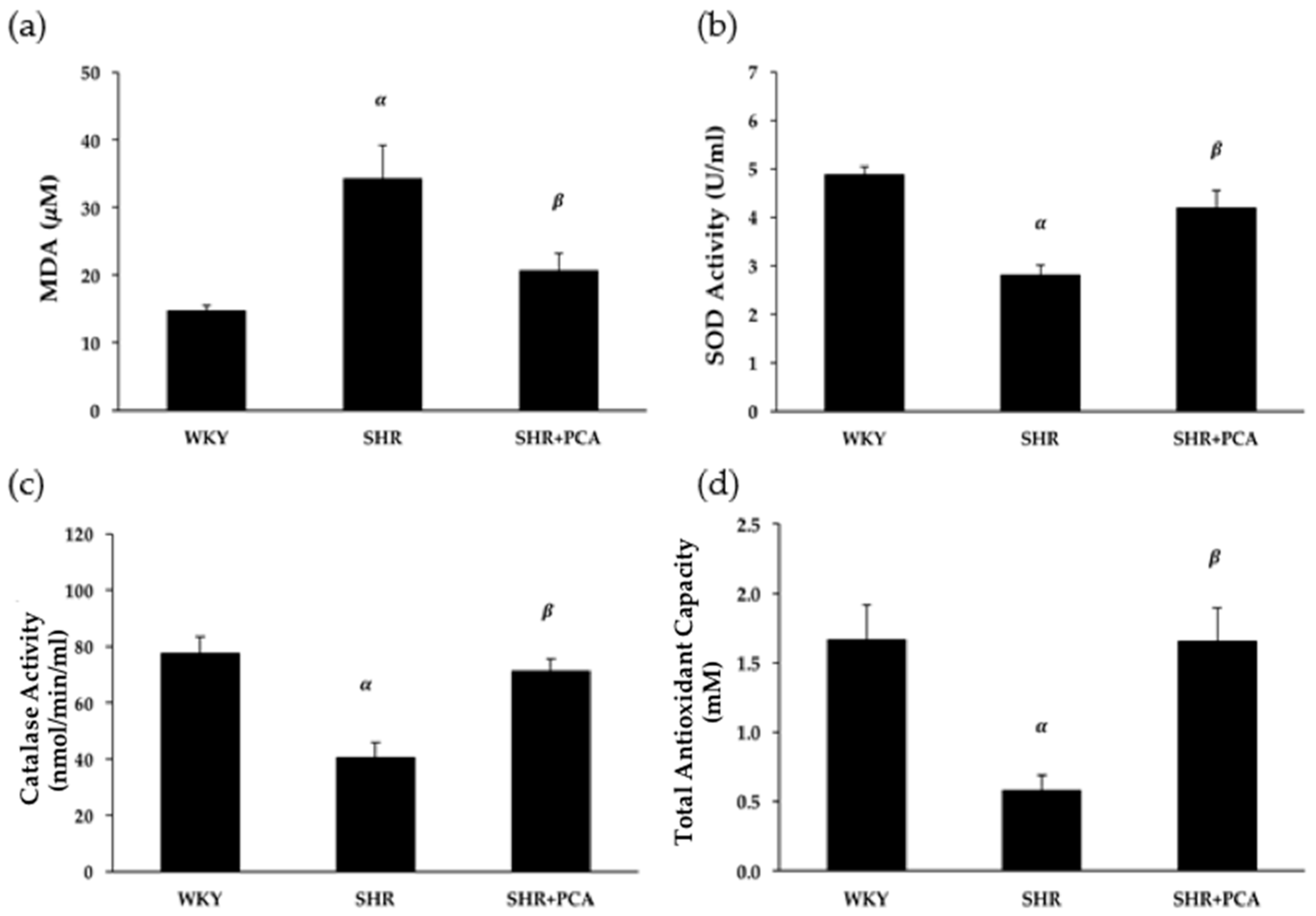
3.6. Serum MDA and Antioxidant Activities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Whelton, P.K.; Carey, R.M.; Aronow, W.S.; Casey, D.E., Jr.; Collins, K.J.; Dennison Himmelfarb, C.; DePalma, S.M.; Gidding, S.; Jamerson, K.A.; Jones, D.W.; et al. 2017 ACC/AHA/AAPA/ABC/ACPM/AGS/APhA/ASH/ASPC/NMA/PCNA Guideline for the Prevention, Detection, Evaluation, and Management of High Blood Pressure in Adults: Executive Summary: A Report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. Hypertension 2017. [Google Scholar] [CrossRef]
- Burnier, M.; Wuerzner, G. Pathophysiology of Hypertension. In Pathophysiology and Pharmacotherapy of Cardiovascular Disease; Jagadeesh, G., Balakumar, P., Maung-U, K., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 655–683. [Google Scholar]
- Hall, J.E.; Granger, J.P.; do Carmo, J.M.; da Silva, A.A.; Dubinion, J.; George, E.; Hamza, S.; Speed, J.; Hall, M.E. Hypertension: Physiology and pathophysiology. Compr. Physiol. 2012, 2, 2393–2442. [Google Scholar] [CrossRef] [PubMed]
- Park, C.G. Hypertension and Vascular Aging. Korean Circ. J. 2006, 36, 477–481. [Google Scholar] [CrossRef]
- Harvey, A.; Montezano, A.C.; Touyz, R.M. Vascular biology of ageing-Implications in hypertension. J. Mol. Cell. Cardiol. 2015, 83, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Papakatsika, S.; Stabouli, S.; Antza, C.; Kotsis, V. Early Vascular Aging: A New Target for Hypertension Treatment. Curr. Pharm. Des. 2016, 22, 122–126. [Google Scholar] [CrossRef]
- Cahill, P.A.; Redmond, E.M. Vascular endothelium—Gatekeeper of vessel health. Atherosclerosis 2016, 248, 97–109. [Google Scholar] [CrossRef]
- Giles, T.D.; Sander, G.E.; Nossaman, B.D.; Kadowitz, P.J. Impaired vasodilation in the pathogenesis of hypertension: Focus on nitric oxide, endothelial-derived hyperpolarizing factors, and prostaglandins. J. Clin. Hypertens. (Greenwich) 2012, 14, 198–205. [Google Scholar] [CrossRef]
- Guerrero, F.; Thioub, S.; Goanvec, C.; Theunissen, S.; Feray, A.; Balestra, C.; Mansourati, J. Effect of tetrahydrobiopterin and exercise training on endothelium-dependent vasorelaxation in SHR. J. Physiol. Biochem. 2013, 69, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Versari, D.; Daghini, E.; Virdis, A.; Ghiadoni, L.; Taddei, S. Endothelium-dependent contractions and endothelial dysfunction in human hypertension. Br. J. Pharmacol. 2009, 157, 527–536. [Google Scholar] [CrossRef] [Green Version]
- Abbas, A.; Grant, P.J.; Kearney, M.T. Role of IGF-1 in glucose regulation and cardiovascular disease. Expert Rev. Cardiovasc. Ther. 2008, 6, 1135–1149. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, L.; Horman, S.; Beauloye, C.; Vanoverschelde, J.L. Insulin signalling in the heart. Cardiovasc. Res. 2008, 79, 238–248. [Google Scholar] [CrossRef] [Green Version]
- Muniyappa, R.; Montagnani, M.; Koh, K.K.; Quon, M.J. Cardiovascular actions of insulin. Endocr. Rev. 2007, 28, 463–491. [Google Scholar] [CrossRef]
- Lin, Y.Y.; Lee, S.D.; Su, C.T.; Cheng, T.L.; Yang, A.L. Long-term treadmill training ameliorates endothelium-dependent vasorelaxation mediated by insulin and insulin-like growth factor-1 in hypertension. J. Appl. Physiol. (1985) 2015, 119, 663–669. [Google Scholar] [CrossRef] [Green Version]
- Vecchione, C.; Colella, S.; Fratta, L.; Gentile, M.T.; Selvetella, G.; Frati, G.; Trimarco, B.; Lembo, G. Impaired insulin-like growth factor I vasorelaxant effects in hypertension. Hypertension 2001, 37, 1480–1485. [Google Scholar] [CrossRef]
- Yang, A.L.; Chao, J.I.; Lee, S.D. Altered insulin-mediated and insulin-like growth factor-1-mediated vasorelaxation in aortas of obese Zucker rats. Int. J. Obes. 2007, 31, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.L.; Yeh, C.K.; Su, C.T.; Lo, C.W.; Lin, K.L.; Lee, S.D. Aerobic exercise acutely improves insulin- and insulin-like growth factor-1-mediated vasorelaxation in hypertensive rats. Exp. Physiol. 2010, 95, 622–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seals, D.R.; Kaplon, R.E.; Gioscia-Ryan, R.A.; LaRocca, T.J. You’re only as old as your arteries: Translational strategies for preserving vascular endothelial function with aging. Physiology 2014, 29, 250–264. [Google Scholar] [CrossRef] [PubMed]
- Mojiminiyi, F.B.; Dikko, M.; Muhammad, B.Y.; Ojobor, P.D.; Ajagbonna, O.P.; Okolo, R.U.; Igbokwe, U.V.; Mojiminiyi, U.E.; Fagbemi, M.A.; Bello, S.O.; et al. Antihypertensive effect of an aqueous extract of the calyx of Hibiscus sabdariffa. Fitoterapia 2007, 78, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Sarr, M.; Ngom, S.; Kane, M.O.; Wele, A.; Diop, D.; Sarr, B.; Gueye, L.; Andriantsitohaina, R.; Diallo, A.S. In vitro vasorelaxation mechanisms of bioactive compounds extracted from Hibiscus sabdariffa on rat thoracic aorta. Nutr. Metab. 2009, 6, 45. [Google Scholar] [CrossRef]
- Semaming, Y.; Pannengpetch, P.; Chattipakorn, S.C.; Chattipakorn, N. Pharmacological properties of protocatechuic Acid and its potential roles as complementary medicine. Evid.-Based Complement. Altern. Med. 2015, 2015, 593902. [Google Scholar] [CrossRef] [PubMed]
- Safaeian, L.; Hajhashemi, V.; Haghjoo Javanmard, S.; Sanaye Naderi, H. The Effect of Protocatechuic Acid on Blood Pressure and Oxidative Stress in Glucocorticoid-induced Hypertension in Rat. Iran J. Pharm. Res. 2016, 15, 83–91. [Google Scholar] [PubMed]
- Shi, G.F.; An, L.J.; Jiang, B.; Guan, S.; Bao, Y.M. Alpinia protocatechuic acid protects against oxidative damage in vitro and reduces oxidative stress in vivo. Neurosci. Lett. 2006, 403, 206–210. [Google Scholar] [CrossRef]
- Harini, R.; Pugalendi, K.V. Antihyperglycemic effect of protocatechuic acid on streptozotocin-diabetic rats. J. Basic Clin. Physiol. Pharmacol. 2010, 21, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Semaming, Y.; Kumfu, S.; Pannangpetch, P.; Chattipakorn, S.C.; Chattipakorn, N. Protocatechuic acid exerts a cardioprotective effect in type 1 diabetic rats. J. Endocrinol. 2014, 223, 13–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, R.; Li, H.; Fan, W.; Jin, Q.; Chao, T.; Wu, Y.; Huang, J.; Hao, L.; Yang, X. Leucine Supplementation Differently Modulates Branched-Chain Amino Acid Catabolism, Mitochondrial Function and Metabolic Profiles at the Different Stage of Insulin Resistance in Rats on High-Fat Diet. Nutrients 2017, 9, 565. [Google Scholar] [CrossRef]
- McCallum, R.W.; Hamilton, C.A.; Graham, D.; Jardine, E.; Connell, J.M.; Dominiczak, A.F. Vascular responses to IGF-I and insulin are impaired in aortae of hypertensive rats. J. Hypertens. 2005, 23, 351–358. [Google Scholar] [CrossRef]
- Scazzocchio, B.; Vari, R.; Filesi, C.; Del Gaudio, I.; D’Archivio, M.; Santangelo, C.; Iacovelli, A.; Galvano, F.; Pluchinotta, F.R.; Giovannini, C.; et al. Protocatechuic acid activates key components of insulin signaling pathway mimicking insulin activity. Mol. Nutr. Food Res. 2015, 59, 1472–1481. [Google Scholar] [CrossRef] [PubMed]
- Pagliaro, B.; Santolamazza, C.; Simonelli, F.; Rubattu, S. Phytochemical Compounds and Protection from Cardiovascular Diseases: A State of the Art. Biomed. Res. Int. 2015, 2015, 918069. [Google Scholar] [CrossRef]
- Zhou, M.S.; Wang, A.; Yu, H. Link between insulin resistance and hypertension: What is the evidence from evolutionary biology? Diabetol. Metab. Syndr. 2014, 6, 12. [Google Scholar] [CrossRef]
- Montezano, A.C.; Dulak-Lis, M.; Tsiropoulou, S.; Harvey, A.; Briones, A.M.; Touyz, R.M. Oxidative stress and human hypertension: Vascular mechanisms, biomarkers, and novel therapies. Can. J. Cardiol. 2015, 31, 631–641. [Google Scholar] [CrossRef]
- Seals, D.R.; Jablonski, K.L.; Donato, A.J. Aging and vascular endothelial function in humans. Clin. Sci. (Lond.) 2011, 120, 357–375. [Google Scholar] [CrossRef] [Green Version]
- Guzik, T.J.; Touyz, R.M. Oxidative Stress, Inflammation, and Vascular Aging in Hypertension. Hypertension 2017, 70, 660–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masodsai, K.; Lin, Y.Y.; Lee, S.D.; Yang, A.L. Exercise and Endothelial Dysfunction in Hypertension. Adapt. Med. 2017, 9, 1–14. [Google Scholar] [CrossRef]
- Safaeian, L.; Emami, R.; Hajhashemi, V.; Haghighatian, Z. Antihypertensive and antioxidant effects of protocatechuic acid in deoxycorticosterone acetate-salt hypertensive rats. Biomed. Pharmacother. 2018, 100, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Seo, H.W.; Lee, M.H.; Kim, D.K.; Jeon, H.; Cha, D.S. Protocatechuic acid extends lifespan and increases stress resistance in Caenorhabditis elegans. Arch. Pharm. Res. 2014, 37, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shi, G.F.; Liu, X.Z.; An, L.J.; Guan, S. Anti-ageing effects of protocatechuic acid from Alpinia on spleen and liver antioxidative system of senescent mice. Cell Biochem. Funct. 2011, 29, 342–347. [Google Scholar] [CrossRef] [PubMed]
- Varì, R.; D’Archivio, M.; Filesi, C.; Carotenuto, S.; Scazzocchio, B.; Santangelo, C.; Giovannini, C.; Masella, R. Protocatechuic acid induces antioxidant/detoxifying enzyme expression through JNK-mediated Nrf2 activation in murine macrophages. J. Nutr. Biochem. 2011, 22, 409–417. [Google Scholar] [CrossRef]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef]
- Li, Q.X.; Xiong, Z.Y.; Hu, B.P.; Tian, Z.J.; Zhang, H.F.; Gou, W.Y.; Wang, H.C.; Gao, F.; Zhang, Q.J. Aging-associated insulin resistance predisposes to hypertension and its reversal by exercise: The role of vascular vasorelaxation to insulin. Basic Res. Cardiol. 2009, 104, 269–284. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters/Groups | WKY | SHR | SHR+PCA |
|---|---|---|---|
| Body weight (g) | 395.75 ± 8.47 | 393.50 ± 5.15 | 404.50 ± 8.85 |
| Heart rate (bpm) | 284.06 ± 3.90 | 383.19 ± 9.83 α | 363.38 ± 6.19 α,β |
| SBP (mmHg) | 121.75 ± 2.15 | 192.88 ± 2.28 α | 173.63 ± 0.88 α,β |
| Insulin (μg/L) | 0.34 ± 0.07 | 0.66 ± 0.11 α | 0.37 ± 0.09 β |
| Blood glucose (mg/dL) | 96.63 ± 2.63 | 121.25 ± 4.76 α | 110.13 ± 4.33 α,β |
| HOMA-IR | 2.02 ± 0.41 | 4.89 ± 0.82 α | 2.50 ± 0.54 β |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masodsai, K.; Lin, Y.-Y.; Chaunchaiyakul, R.; Su, C.-T.; Lee, S.-D.; Yang, A.-L. Twelve-Week Protocatechuic Acid Administration Improves Insulin-Induced and Insulin-Like Growth Factor-1-Induced Vasorelaxation and Antioxidant Activities in Aging Spontaneously Hypertensive Rats. Nutrients 2019, 11, 699. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11030699
Masodsai K, Lin Y-Y, Chaunchaiyakul R, Su C-T, Lee S-D, Yang A-L. Twelve-Week Protocatechuic Acid Administration Improves Insulin-Induced and Insulin-Like Growth Factor-1-Induced Vasorelaxation and Antioxidant Activities in Aging Spontaneously Hypertensive Rats. Nutrients. 2019; 11(3):699. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11030699
Chicago/Turabian StyleMasodsai, Kunanya, Yi-Yuan Lin, Rungchai Chaunchaiyakul, Chia-Ting Su, Shin-Da Lee, and Ai-Lun Yang. 2019. "Twelve-Week Protocatechuic Acid Administration Improves Insulin-Induced and Insulin-Like Growth Factor-1-Induced Vasorelaxation and Antioxidant Activities in Aging Spontaneously Hypertensive Rats" Nutrients 11, no. 3: 699. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11030699