Dietary Fatty Acids in Postprandial Triglyceride-Rich Lipoproteins Modulate Human Monocyte-Derived Dendritic Cell Maturation and Activation

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Postprandial Study and TRL Isolation
2.2. Fat and TRL Fatty Acid Composition
2.3. Monocyte Isolation
2.4. MonocyteDerived Dendritic Cell Maturation and Activation
2.5. Monocyte-Derived Dendritic Cell Viability
2.6. Triglyceride Quantification
2.7. RNA Isolation and RT-qPCR
2.8. Cytokine Quantification
2.9. Statistical Analysis
3. Results
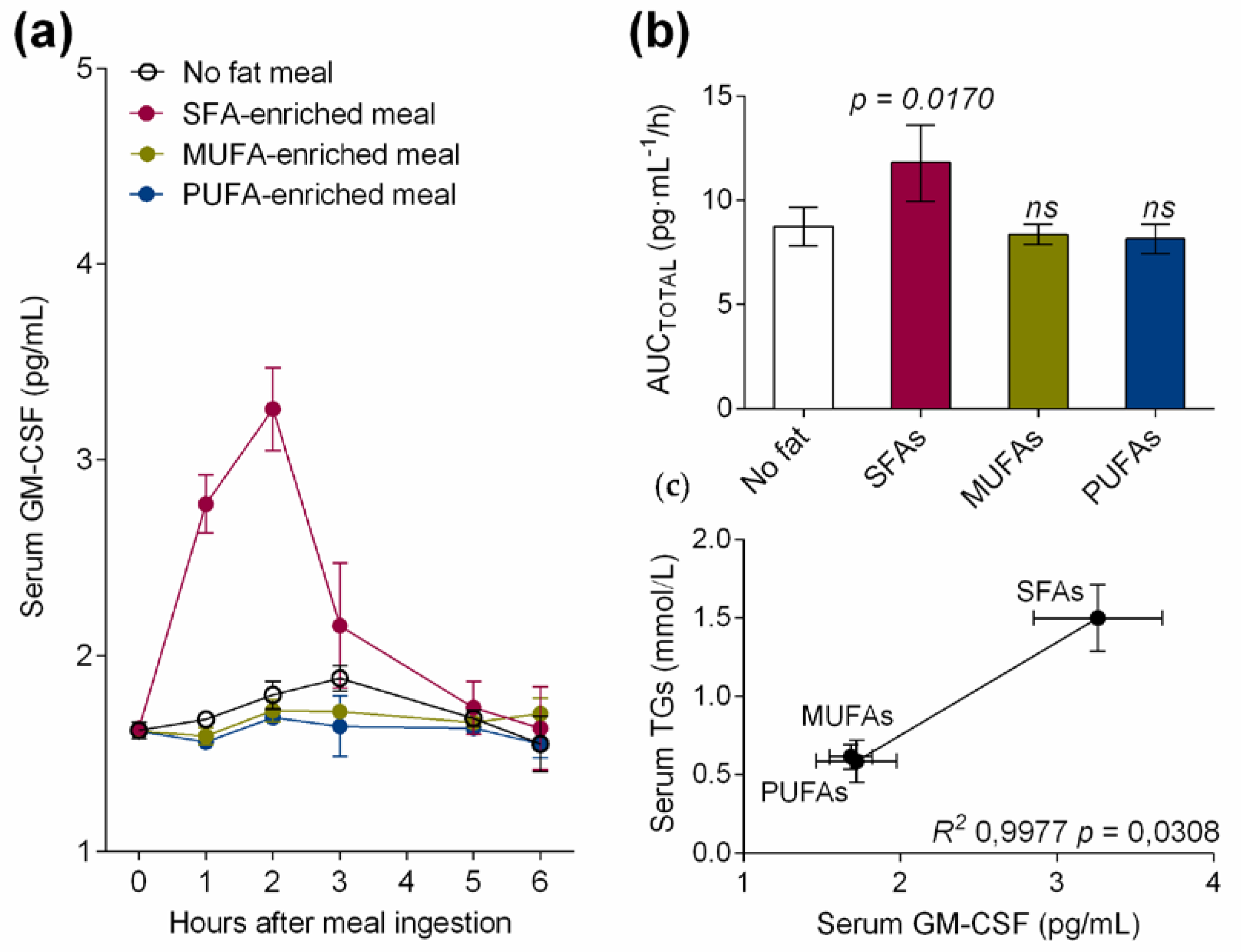
3.1. Dietary Saturated Fatty Acids Acutely Increase Serum GM-CSF Levels in the Postprandial State of Healthy Volunteers
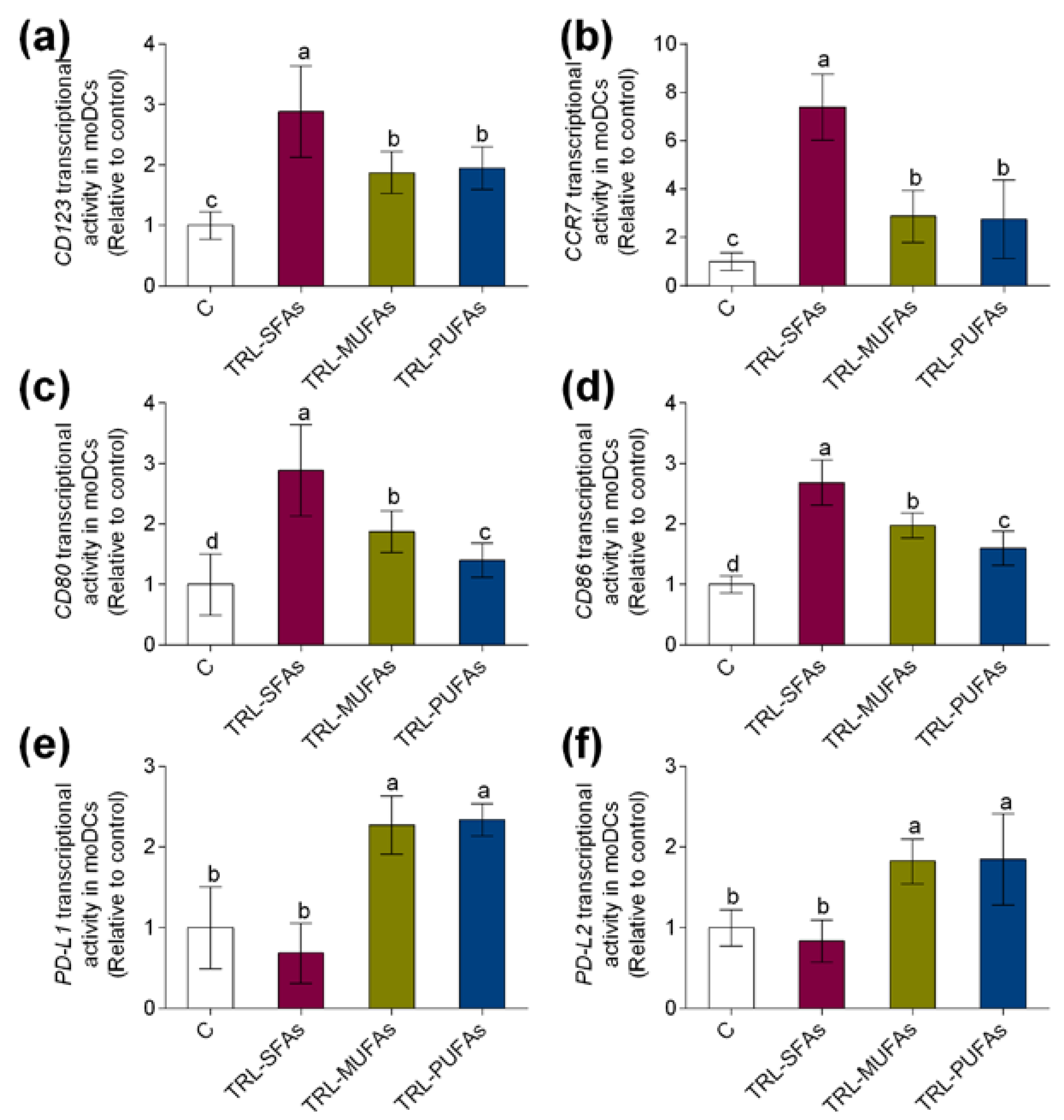
3.2. Triglyceride Rich-Lipoproteins Modulate Maturation and Activation Markers in Human Monocyte-Derived Dendritic Cells
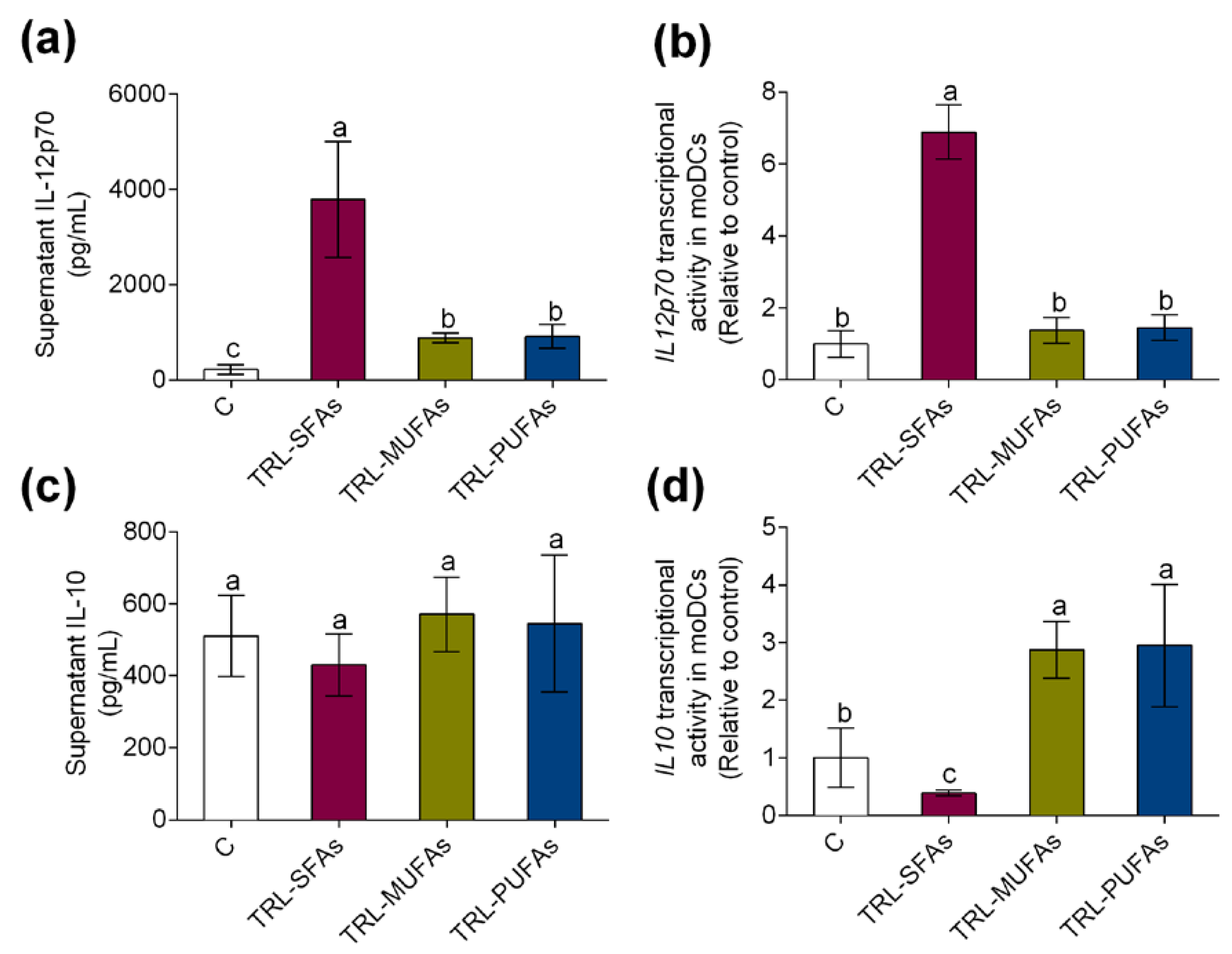
3.3. Fatty Acid-Enriched Meals Modulate Serum IL-12p70 and IL-10 Postprandial Secretion and Triglyceride-Rich Lipoproteins Regulate Cytokine Levels and Gene Expression in Human Monocyte-Derived Dendritic Cells
3.4. Triglyceride-Rich Lipoproteins Induce Lipid Accumulation and ApoB48R Transcriptional Activity in Monocyte-Derived Dendritic Cells in a Fatty Acid-Dependent Manner
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| DC | dendritic cell |
| GM-CSF | granulocyte/macrophage colony stimulating factor |
| IL | interleukin |
| mDC | myeloid dendritic cell |
| moDC | monocyte-derived dendritic cell |
| MUFA | monounsaturated fatty acid |
| pDC, | plasmacytoid dendritic cell |
| SFA | saturated fatty acid |
| TG | triglyceride |
| TRL | triglyceride-rich lipoprotein |
References
- Ayres, J.S. Immunometabolism of infections. Nat. Rev. Immunol. 2020, 20, 79–80. [Google Scholar] [CrossRef] [PubMed]
- Luque-Sierra, A.; Alvarez-Amor, L.; Kleemann, R.; Martin, F.; Varela, L.M. Extra-Virgin Olive Oil with Natural Phenolic Content Exerts an Anti-Inflammatory Effect in Adipose Tissue and Attenuates the Severity of Atherosclerotic Lesions in Ldlr-/-.Leiden Mice. Mol. Nutr. Food Res. 2018, 62, 1800295. [Google Scholar] [CrossRef] [PubMed]
- Salvatore, G.; Bernoud-Hubac, N.; Bissay, N.; Debard, C.; Daira, P.; Meugnier, E.; Proamer, F.; Hanau, D.; Vidal, H.; Arico, M.; et al. Human monocyte-derived dendritic cells turn into foamy dendritic cells with IL-17A. J. Lipid Res. 2015, 56, 1110–1122. [Google Scholar] [CrossRef] [Green Version]
- Villar, J.; Segura, E. Recent advances towards deciphering human dendritic cell development. Mol. Immunol. 2020, 122, 109–115. [Google Scholar] [CrossRef]
- Sander, J.; Schmidt, S.; Cirovic, B.; McGovern, N.; Papantonopoulou, O.; Hardt, A.L.; Aschenbrenner, A.C.; Kreer, C.; Quast, T.; Xu, A.M.; et al. Cellular Differentiation of Human Monocytes Is Regulated by Time-Dependent Interleukin-4 Signaling and the Transcriptional Regulator NCOR2. Immunity 2017, 47, 1051–1066. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.A.; Lu, Y.B.; Wang, W.D.; Liu, G.B.; Chen, C.; Shen, L.; Luo, H.L.; Xu, H.; Peng, Y.; Luo, H.; et al. BTLA-Expressing Dendritic Cells in Patients with Tuberculosis Exhibit Reduced Production of IL-12/IFN-α and Increased Production of IL-4 and TGF-β, Favoring Th2 and Foxp3+ Treg Polarization. Front. Immunol. 2020, 11, 518. [Google Scholar] [CrossRef] [PubMed]
- Herber, D.L.; Cao, W.; Nefedova, Y.; Novitskiy, S.V.; Nagaraj, S.; Tyurin, V.A.; Corzo, A.; Cho, H.I.; Celis, E.; Lennox, B. Lipid accumulation and dendritic cell dysfunction in cancer. Nat. Med. 2010, 16, 880–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, H.; Kim, J.; Kim, D.; Lee, J.E.; Chung, Y. Cellular and Molecular Links between Autoimmunity and Lipid Metabolism. Mol. Cells 2019, 42, 747–754. [Google Scholar] [CrossRef]
- Naranjo, M.C.; Garcia, I.; Bermudez, B.; Lopez, S.; Cardelo, M.P.; Abia, R.; Muriana, F.J.G.; Montserrat-de la Paz, S. Acute effects of dietary fatty acids on osteclastogenesis via RANKL/RANK/OPG system. Mol. Nutr. Food Res. 2016, 60, 2505–2513. [Google Scholar] [CrossRef]
- Montserrat-de la Paz, S.; Bermudez, B.; Cardelo, M.P.; Lopez, S.; Abia, R.; Muriana, F.J.G. Olive Oil and postprandial hyperlipidemia: Implications for atherosclerosis and metabolic syndrome. Food Funct. 2016, 7, 4734–4744. [Google Scholar] [CrossRef] [Green Version]
- Montserrat-de la Paz, S.; Bermudez, B.; Lopez, S.; Naranjo, M.C.; Romero, Y.; Bando-Hidalgo, M.J.; Abia, R.; Muriana, F.J.G. Exogenous fatty acids and niacin on acute prostaglandin D2 production in human myeloid cells. J. Nutr. Biochem. 2017, 39, 22–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montserrat-de la Paz, S.; Lopez, S.; Bermudez, B.; Guerrero, J.M.; Abia, R.; Muriana, F.J.G. Effects of immediate-release niacin and dietary fatty acids on acute insulin and lipid status in individuals with metabolic syndrome. J. Sci. Food Agric. 2018, 98, 2194–2200. [Google Scholar] [CrossRef]
- Toscano, R.; Millan-Linares, M.C.; Lemus-Conejo, A.; Claro, C.; Sanchez-Margalet, V.; Montserrat-de la Paz, S. Postprandial triglyceride-rich lipoproteins promote M1/M2 microglia polarization in a fatty-acid-dependent manner. J. Nutr. Biochem. 2020, 75, 108248. [Google Scholar] [CrossRef]
- Alipour, A.; Van Oostrom, A.J.; Izraeljan, A.; Verseyden, C.; Collins, J.M.; Frayn, K.N.; Plokker, T.W.M.; Elte, J.W.F.; Castro Cabezas, M. Leukocyte activation by triglyceride-rich lipoproteins. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 792–797. [Google Scholar] [CrossRef]
- Varela, L.M.; Ortega-Gomez, A.; Lopez, S.; Abia, R.; Muriana, F.J.G.; Bermudez, B. The effects of dietary fatty acids on the postprandial triglyceride-rich lipoprotein/apoB48 receptor axis in human monocyte/macrophage cells. J. Nutr. Biochem. 2013, 24, 2031–2039. [Google Scholar] [CrossRef] [PubMed]
- Varela, L.M.; Bermudez, B.; Ortega-Gomez, A.; Lopez, S.; Sanchez, R.; Villar, J.; Muriana, F.J.G.; Abia, R. Postprandial triglyceride-rich lipoproteins promote invasion of human coronary artery smooth muscle cells in a fatty-acid manner through Pl3k-Rac1-JNK signalling. Mol. Nutr. Food Res. 2014, 58, 1349–1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EEC. Comision regulation (EEC) no. 796/02 of 6 May 2002 amending regulation EEC2568/91 on the characteristics of olive oil and olive residue oil and on the relevant methodology analysis. Off. J. Eur. Commun. 2002, 128, 8–28. [Google Scholar]
- Montserrat-de la Paz, S.; Naranjo, M.C.; Bermudez, B.; Lopez, S.; Moreda, W.; Abia, R.; Muriana, F.J.G. Postprandial dietary fatty acids exert divergent inflammatory response in retinal-pigmented epithelium cells. Food Funct. 2016, 7, 1345–1353. [Google Scholar] [CrossRef]
- Montserrat-de la Paz, S.; de la Puerta, R.; Fernandez-Arche, A.; Quilez, A.M.; Muriana, F.J.G.; Garcia-Gimenez, M.D.; Bermudez, B. Pharmacological effects of mitraphylline from Uncaria tomentosa in primary human monocytes: Skew toward M2 macrophages. J. Ethnopharmacol. 2015, 170, 128–135. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Ikeda, K.; Maeda, Y.; Shinagawa, K.; Ohtsuka, A.; Yamamura, H.; Tanimoto, M. Identification of CD123+ myeloid dendritic cells as an early-stage immature subset with strong tumoristatic potential. Cancer Let. 2008, 270, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Collin, M.; Bigley, V. Human dendritic cell subsets: An update. Immunology 2018, 154, 3–20. [Google Scholar] [CrossRef]
- Sugimoto, C.; Hasegawa, A.; Saito, Y.; Fukuyo, Y.; Chiu, K.B.; Cai, Y.; Breed, M.W.; Mori, K.; Roy, C.J.; Lackner, A.A.; et al. Differentiation Kinetics of Blood Monocytes and Dendritic Cells in Macaques: Insights to Understanding Human Myeloid Cell Development. J. Immunol. 2015, 1954, 1774–1781. [Google Scholar] [CrossRef]
- Roncero-Ramos, I.; Rangel-Zuñiga, O.A.; Lopez-Moreno, J.; Alcala-Diaz, J.F.; Perez-Martinez, P.; Jimenez-Lucena, R.; Castaño, J.P.; Roche, H.M.; Delgado-Lista, J.; Ordovas, J.M.; et al. Mediterranean Diet, Glucose Homeostasis, and Inflammasome Genetic Variants: The CORDIOPREV Study. Mol. Nutr. Food Res. 2018, 62, e1700960. [Google Scholar] [CrossRef]
- Hernaez, A.; Sanllorente, A.; Castañer, O.; Martinez-Gonzalez, M.A.; Ros, E.; Pinto, X.; Estruch, R.; Salas-Salvado, J.; Corella, D.; Alonso-Gomez, A.M.; et al. Increased Consumption of Virgin Olive Oil, Nuts, Legumes, Whole Grains, and Fish Promotes HDL Functions in Humans. Mol. Nutr. Food Res. 2019, 63, e1800847. [Google Scholar] [CrossRef]
- Montserrat-de la Paz, S.; Naranjo, M.C.; Millan-Linares, M.C.; Lopez, S.; Abia, R.; Biessen, E.A.L.; Muriana, F.J.G.; Bermudez, B. Monounsaturated Fatty Acids in a High-Fat Diet and Niacin Protect from White Fat Dysfunction in the Metabolic Syndrome. Mol. Nutr. Food Res. 2019, 63, 1900425. [Google Scholar] [CrossRef]
- Lopez, S.; Garcia-Serrano, S.; Gutierrez-Repiso, C.; Rodriguez-Pacheco, F.; Ho-Plagaro, E.; Santiago-Fernandez, C.; Alba, G.; Cejudo-Guillen, M.; Rodrigue-Cañete, A.; Valdes, S.; et al. Tissue-Specific Phenotype and Activation of iNKT Cells in Morbidly Obese Subjects: Interaction with Adipocytes and Effect of Bariatric Surgery. Obes. Surg. 2018, 28, 2774–2782. [Google Scholar] [CrossRef]
- Ganguly, D.; Haak, S.; Sisirak, V.; Reizis, B. The role of dendritic cells in autoimmunity. Nat. Rev. Immunol. 2013, 13, 566–577. [Google Scholar] [CrossRef] [Green Version]
- Mraz, M.; Cinkajzlova, A.; Klouckova, J.; Lacinova, Z.; Kratochvilova, H.; Lips, M.; Porizka, M.; Kopecky, P.; Lindner, J.; Kotulak, T.; et al. Dendritic Cells in Subcutaneous and Epicardial Adipose Tissue of Subjects with Type 2 Diabetes, Obesity, and Coronary Artery Disease. Mediators Inflamm. 2019, 2019, 5481725. [Google Scholar] [CrossRef]
- Radzikowska, U.; Rinaldi, A.O.; Sozener, Z.C.; Karaguzel, D.; Wojcik, M.; Cypryk, K.; Akdis, M.; Akdis, C.A.; Sokolowska, M. The Influence of Dietary Fatty Acids on Immune Responses. Nutrients 2019, 11, 2990. [Google Scholar] [CrossRef] [Green Version]
- Nastasi, C.; Candela, M.; Bonefeld, C.M.; Geisler, C.; Hansen, M.; Krejsgaard, T.; Biagi, E.; Andersen, M.H.; Brigidi, P.; Odum, N.; et al. The effect of short-chain fatty acids on human monocyte-derived dendritic cells. Sci. Rep. 2015, 5, 16148. [Google Scholar] [CrossRef]
- Lopez, S.; Bermudez, B.; Ortega, A.; Varela, L.M.; Pacheco, Y.M.; Villar, J.; Abia, R.; Muriana, F.J.G. Effects of meals rich in either monounsaturated or saturated fat on lipid concentrations and on insulin secretion and action in subjects with high fasting triglyceride concentrations. Am. J. Clin. Nutr. 2011, 93, 494–499. [Google Scholar] [CrossRef]
- Khan, I.M.; Pokharel, Y.; Dadu, R.T.; Lewis, D.E.; Hoogeveen, R.C.; Wu, H.; Ballantyne, C.M. Postprandial Monocyte Activation in Individuals with Metabolic Syndrome. J. Clin. Endocrinol. Metab. 2016, 101, 4195–4204. [Google Scholar] [CrossRef] [Green Version]
- Montserrat-de la Paz, S.; Rodriguez, D.; Cardelo, M.P.; Naranjo, M.C.; Bermudez., B.; Abia, R.; Muriana, F.J.G.; Lopez, S. The effects of exogenous fatty acids and niacin on human monocyte-macrophage plasticity. Mol. Nutr. Food Res. 2017, 61, 1600824. [Google Scholar] [CrossRef]
- McFarlin, B.K.; Carpenter, K.C.; Henning, A.L.; Venable, A.S. Consumption of a high-fat breakfast on consecutive days alters preclinical biomarkers for atherosclerosis. Eur. J. Clin. Nutr. 2017, 71, 239–244. [Google Scholar] [CrossRef]
- Nicholas, D.A.; Zhang, K.; Hung, C.; Glasgow, S.; Aruni, A.W.; Unternaehrer, J.; Payne, K.J.; Langridge, W.H.R.; De Leon, M. Palmitic acid is a toll-like receptor 4 ligand that induces human dendritic cell secretion of IL-1beta. PLoS ONE 2017, 12, e0176793. [Google Scholar] [CrossRef]
- Kong, W.; Yen, J.H.; Vassiliou, E.; Adhikary, S.; Toscano, M.G.; Ganea, D. Docosahexaenoic acid prevents dendritic cell maturation and in vitro and in vivo expression of the IL-12 cytokine family. Lipids Health Dis. 2010, 9, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortega-Gomez, A.; Varela, L.M.; Lopez, S.; Montserrat-de la Paz, S.; Sanchez, R.; Muriana, F.J.G.; Bermudez, B.; Abia, R. Postprandial triglyceride-rich lipoproteins promote lipid accumulation and apolipoprotein B-48 receptor transcriptional activity in human circulating and murine bone marrow neutrophils in a fatty acid-dependent manner. Mol. Nutr. Food Res. 2017, 61, 1600879. [Google Scholar] [CrossRef]
- Varela, L.M.; Ortega, A.; Bermudez, B.; Lopez, S.; Pacheco, Y.M.; Villar, J.; Abia, R.; Muriana, F.J.G. A high-fat meal promotes lipid-load and apolipoprotein B-48 receptor transcriptional activity in circulating monocytes. Am. J. Clin. Nutr. 2011, 93, 918–925. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid | TRL-SFAs | TRL-MUFAs | TRL-PUFAs |
|---|---|---|---|
| g/100 g of Fatty Acid | |||
| 4:0, butyric | 0.26 ± 0.03 | - | - |
| 6:0, caproic | 0.19 ± 0.02 | - | - |
| 8:0, caprylic | 0.38 ± 0.14 | - | - |
| 10:0, capric | 1.62 ± 0.52 | - | - |
| 12:0, lauric | 3.52 ± 1.01 | - | - |
| 14:0, myristic | 8.76 ± 1.63 | - | - |
| 16:0, palmitic | 38.10 ± 1.87 | 11.2 ± 1.52 | 11.98 ± 1.21 |
| 16:1(n-7), palmitoleic | 1.03 ± 0.10 | 0.79 ± 0.21 | 1.42 ± 0.61 |
| 18:0, stearic | 18.8 ± 1.32 | 5.71 ± 0.73 | 5.54 ± 0.78 |
| 18:1(n-9), oleic | 20.7 ± 1.76 | 67.2 ± 2.97 | 61.3 ± 3,87 |
| 18:2(n-6), linoleic | 4.04 ± 0.98 | 8.95 ± 1.32 | 9,06 ± 1.03 |
| 18:3(n-3), α-linolenic | 1.96 ± 0.57 | 3.29 ± 0.74 | 3.03 ± 0.98 |
| 20:4(n-6), arachidonic | 0.49 ± 0.09 | 1.18 ± 0.37 | 1.79 ± 0.42 |
| 20:5(n-3), eicosapentaenoic | - | 0.78 ± 0.19 | 2.82 ± 0.29 |
| 22:6(n-3), docosahexaenoic | - | 0.70 ± 0.28 | 2.63 ± 0.14 |
| Others | 0.51 ± 0.13 | 0.34 ± 0.07 | 0.56 ± 0.07 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vazquez-Madrigal, C.; Lopez, S.; Grao-Cruces, E.; Millan-Linares, M.C.; Rodriguez-Martin, N.M.; Martin, M.E.; Alba, G.; Santa-Maria, C.; Bermudez, B.; Montserrat-de la Paz, S. Dietary Fatty Acids in Postprandial Triglyceride-Rich Lipoproteins Modulate Human Monocyte-Derived Dendritic Cell Maturation and Activation. Nutrients 2020, 12, 3139. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12103139
Vazquez-Madrigal C, Lopez S, Grao-Cruces E, Millan-Linares MC, Rodriguez-Martin NM, Martin ME, Alba G, Santa-Maria C, Bermudez B, Montserrat-de la Paz S. Dietary Fatty Acids in Postprandial Triglyceride-Rich Lipoproteins Modulate Human Monocyte-Derived Dendritic Cell Maturation and Activation. Nutrients. 2020; 12(10):3139. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12103139
Chicago/Turabian StyleVazquez-Madrigal, Carlos, Soledad Lopez, Elena Grao-Cruces, Maria C. Millan-Linares, Noelia M. Rodriguez-Martin, Maria E. Martin, Gonzalo Alba, Consuelo Santa-Maria, Beatriz Bermudez, and Sergio Montserrat-de la Paz. 2020. "Dietary Fatty Acids in Postprandial Triglyceride-Rich Lipoproteins Modulate Human Monocyte-Derived Dendritic Cell Maturation and Activation" Nutrients 12, no. 10: 3139. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12103139