Use of an Animal Model to Evaluate Anxiolytic Effects of Dietary Supplementation with Tilia tomentosa Moench Bud Extracts

 ,
,  ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Raw Samples
2.2. Spectroscopic Analysis: UV-Visible Fingerprint
2.3. HPLC Analysis
2.4. Animals
2.5. Dietary Supplementation and Testing Procedure
2.6. Hole Board
2.7. Light/Dark Box
2.8. Release Studies
2.9. Experiments of Release
2.10. Statistical Analysis
2.11. Drugs
3. Results
3.1. Bud-Extracts Characterization: UV-Visible and HPLC Fingerprints
3.2. Evaluation Active Dilutions of TTBEs on Native Brain Targets
3.3. Dietary Supplementation with TTBEs: Control of Daily Intake and Weight Gain
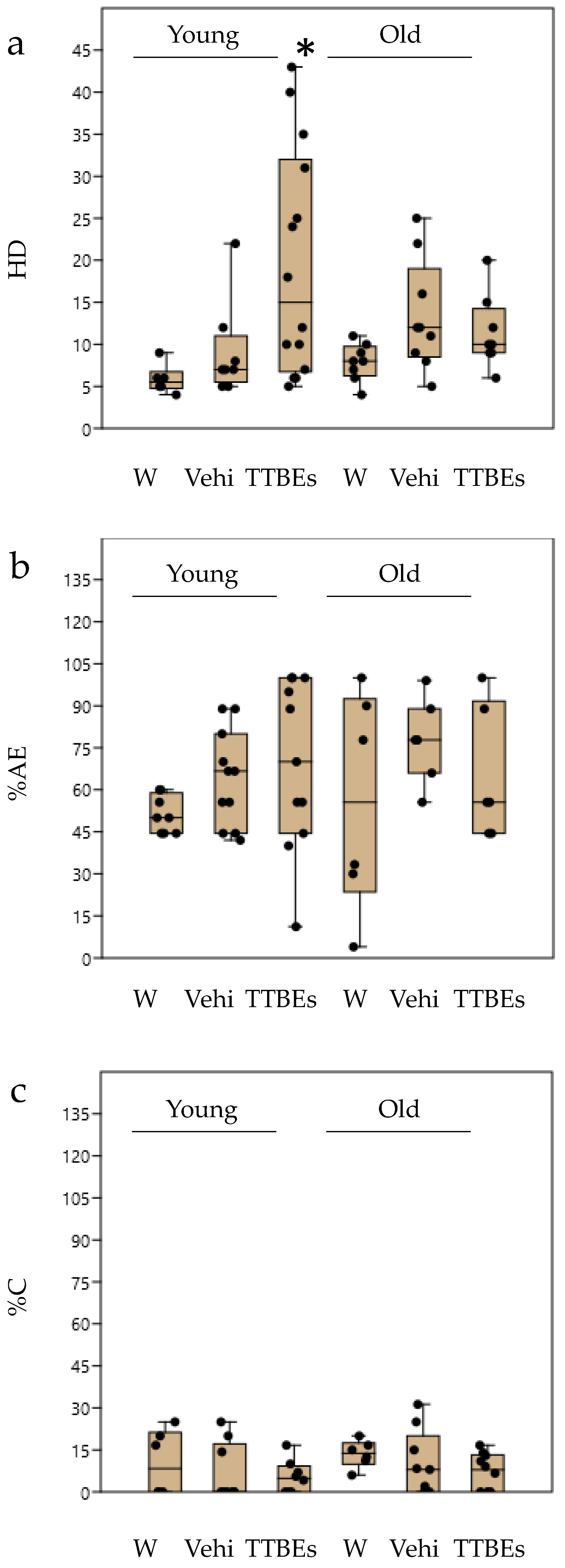
3.4. Dietary Supplementation with TTBEs: Behavioral Scores
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| c1 | c2 | HD_1 | %AE_1 | %C_1 | tL_1 | T_1 | Av Sp_1 | RAE_1 | D_1 | |
|---|---|---|---|---|---|---|---|---|---|---|
| YMW | Y | M | 25.90909 | 56.05949 | −17.1947 | 22.69231 | 1.980769 | 4.915 | −40.85 | 1536.75 |
| YMVehi | Y | M | 18.40909 | 35.68949 | −18.6447 | 12.69231 | 0.900769 | 5.015 | −29.92 | 1561.75 |
| YMTTBEs | Y | M | 13.18909 | 10.99949 | −13.1547 | 2.692308 | −2.21923 | 0.985 | −37.67 | 325.75 |
| OMW | O | M | 26.02294 | 71.24405 | −20.6994 | 24.66667 | 0.72 | 2.865 | 10.05 | 1086 |
| OMVehi | O | M | 25.18294 | 54.57405 | −34.5894 | 44.16667 | 1.55 | 2.915 | 6.58 | 928 |
| OMTTBEs | O | M | 17.06294 | 26.79405 | −12.3794 | 36.66667 | 1.97 | 0.585 | 12.25 | 106 |
| c1 | c2 | HD_1 | %AE_1 | %C_1 | tL_1 | T_1 | Av Sp_1 | RAE_1 | D_1 | |
|---|---|---|---|---|---|---|---|---|---|---|
| YFW | Y | F | 25.58667 | 43.98148 | −2.12525 | 50.125 | 4.25 | 1.895 | −19 | 590.5 |
| YFVehi | Y | F | 22.29667 | 30.09148 | 1.564749 | 11.265 | 1.22 | 1.355 | −18.33 | 434.5 |
| YFTTBEs | Y | F | 11.66667 | 24.54148 | 2.734749 | 20.555 | −1 | 5.085 | −10.1 | 1588.5 |
| OFW | O | F | 20.68294 | 38.46405 | −0.78936 | 43.16667 | −1.45 | 2.755 | 7.25 | 986 |
| OFVehi | O | F | 14.55294 | 17.91405 | 3.120638 | 65.66667 | 2.89 | 2.655 | 8.25 | 852 |
| OFTTBEs | O | F | 17.18294 | 23.09405 | 5.370638 | 60.50667 | 2.62 | 2.265 | 9.25 | 683 |
References
- Foster, J.A.; McVey Neufeld, K.-A. Gut-brain axis: How the microbiome influences anxiety and depression. Trends Neurosci. 2013, 36, 305–312. [Google Scholar] [CrossRef]
- Pandey, M.; Verma, R.K.; Saraf, S.A. Nutraceuticals: New era of medicine and health. Asian J. Pharm. Clin. Res. 2010, 3, 11–15. [Google Scholar]
- Olatunji, B.O.; Wolitzky-Taylor, K.B. Anxiety sensitivity and the anxiety disorders: A meta-analytic review and synthesis. Psychol. Bull. 2009, 135, 974–999. [Google Scholar] [CrossRef]
- Lewinsohn, P.M.; Lewinsohn, M.; Gotlib, I.H.; Seeley, J.R.; Allen, N.B. Gender differences in anxiety disorders and anxiety symptoms in adolescents. J. Abnorm. Psychol. 1998, 107, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Depaola, S.J.; Griffin, M.; Young, J.R.; Neimeyer, R.A. Death anxiety and attitudes toward the elderly among older adults: The role of gender and ethnicity. Death Stud. 2003, 27, 335–354. [Google Scholar] [CrossRef] [PubMed]
- Lynch, S.M. Measurement and Prediction of Aging Anxiety. Res. Aging 2000, 22, 533–558. [Google Scholar] [CrossRef]
- Cazard, F.; Ferreri, F. Troubles bipolaires et troubles anxieux comorbides: Impact pronostique et enjeux therapeutiques. L’encephale 2013, 39, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Meisel, M.K.; Goodie, A.S. Predicting prescription drug misuse in college students’ social networks. Addict. Behav. 2015, 45, 110–112. [Google Scholar] [CrossRef]
- Curran, S.; Musa, S.; Sajjadi, A. Hypnosedatives and Anxiolytics. In Side Effects of Drugs Annual; Elsevier: Amsterdam, The Netherlands, 2015; pp. 57–61. ISBN 9780444635259. [Google Scholar]
- Basile, A.S.; Lippa, A.S.; Skolnick, P. Anxioselective anxiolytics: Can less be more? Eur. J. Pharm. 2004, 500, 441–451. [Google Scholar] [CrossRef]
- Farach, F.J.; Pruitt, L.D.; Jun, J.J.; Jerud, A.B.; Zoellner, L.A.; Roy-Byrne, P.P. Pharmacological treatment of anxiety disorders: Current treatments and future directions. J. Anxiety Disord. 2012, 26, 833–843. [Google Scholar] [CrossRef] [Green Version]
- Alramadhan, E.; Hanna, M.S.; Hanna, M.S.; Goldstein, T.G.; Avila, S.M.; Weeks, B.S. Dietary and botanical anxiolytics. Med. Sci. Monit. 2012, 18, RA40–RA48. [Google Scholar] [CrossRef] [PubMed]
- Calixto, J.B. Efficacy, safety, quality control, marketing and regulatory guidelines for herbal medicines (phytotherapeutic agents). Braz. J. Med. Biol. Res. 2000, 33, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Konik, E.A.; Jungling, R.C.; Bauer, B.A. Herbs and Dietary Supplements in the European Union: A Review of the Regulations with Special Focus on Germany and Poland. J. Diet. Suppl. 2011, 8, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Dell’agli, M.; Di Lorenzo, C.; Badea, M.; Sangiovanni, E.; Dima, L.; Bosisio, E.; Restani, P. Plant Food Supplements with Anti-Inflammatory Properties: A Systematic Review (I). Crit. Rev. Food Sci. Nutr. 2013, 53, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Gulati, O.P.; Berry Ottaway, P.; Coppens, P. Botanical Nutraceuticals, (Food Supplements, Fortified and Functional Foods) in the European Union with Main Focus on Nutrition And Health Claims Regulation. In Nutraceutical and Functional Food Regulations in the United States and Around the World; Elsevier: Amsterdam, The Netherlands, 2014; pp. 221–256. ISBN 9780124058705. [Google Scholar]
- Ieri, F.; Innocenti, M.; Possieri, L.; Gallori, S.; Mulinacci, N. Phenolic composition of “bud extracts” of Ribes nigrum L., Rosa canina L. and Tilia tomentosa M. J. Pharm. Biomed. Anal. 2015, 115, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Donno, D.; Beccaro, G.L.; Mellano, M.G.; Bonvegna, L.; Bounous, G. Castanea spp. buds as a phytochemical source for herbal preparations: Botanical fingerprint for nutraceutical identification and functional food standardisation. J. Sci. Food Agric. 2014, 94, 2863–2873. [Google Scholar] [CrossRef]
- Viola, H.; Wolfman, C.; de Stein, M.L.; Wasowski, C.; Peña, C.; Medina, J.H.; Paladini, A.C. Isolation of pharmacologically active benzodiazepine receptor ligands from Tilia tomentosa (Tiliaceae). J. Ethnopharmacol. 1994, 44, 47–53. [Google Scholar] [CrossRef]
- Allio, A.; Calorio, C.; Franchino, C.; Gavello, D.; Carbone, E.; Marcantoni, A. Bud extracts from Tilia tomentosa Moench inhibit hippocampal neuronal firing through GABAA and benzodiazepine receptors activation. J. Ethnopharmacol. 2015, 172, 288–296. [Google Scholar] [CrossRef]
- Mohler, H. GABA A Receptors in Central Nervous System Disease: Anxiety, Epilepsy, and Insomnia. J. Recept. Signal Transduct. 2006, 26, 731–740. [Google Scholar] [CrossRef]
- Kalueff, A.V.; Nutt, D.J. Role of GABA in anxiety and depression. Depress. Anxiety 2007, 24, 495–517. [Google Scholar] [CrossRef]
- Goddard, A.W.; Ball, S.G.; Martinez, J.; Robinson, M.J.; Yang, C.R.; Russell, J.M.; Shekhar, A. Current perspectives of the roles of the central norepinephrine system in anxiety and depression. Depress. Anxiety 2010, 27, 339–350. [Google Scholar] [CrossRef]
- Nuss, P. Anxiety disorders and GABA neurotransmission: A disturbance of modulation. Neuropsychiatr. Dis. Treat. 2015, 11, 165–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fung, S.-C.; Fillenz, M. The role of pre-synaptic GABA and benzodiazepine receptors in the control of noradrenaline release in rat hippocampus. Neurosci. Lett. 1983, 42, 61–66. [Google Scholar] [CrossRef]
- Schmid, G.; Chittolini, R.; Raiteri, L.; Bonanno, G. Differential effects of zinc on native GABAA receptor function in rat hippocampus and cerebellum. Neurochem. Int. 1999, 34, 399–405. [Google Scholar] [CrossRef]
- Barnes, R.J.; Dhanoa, M.S.; Lister, S.J. Standard normal variate transformation and de-trending of near-infrared diffuse reflectance spectra. Appl. Spectrosc. 1989, 43, 772–777. [Google Scholar] [CrossRef]
- Mok, D.K.W.; Chau, F.T. Chemical information of Chinese medicines: A challenge to chemist. Chemom. Intell. Lab. Syst. 2006, 82, 210–217. [Google Scholar] [CrossRef]
- Bonfiglio, T.; Olivero, G.; Vergassola, M.; Di Cesare Mannelli, L.; Pacini, A.; Iannuzzi, F.; Summa, M.; Bertorelli, R.; Feligioni, M.; Ghelardini, C.; et al. Environmental training is beneficial to clinical symptoms and cortical presynaptic defects in mice suffering from experimental autoimmune encephalomyelitis. Neuropharmacology 2019. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.; Zhang, H.; Klaminder, J.; Brodin, T.; Andersson, P.L.; Andersson, M. ToxTrac: A fast and robust software for tracking organisms. Methods Ecol. Evol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Bonfiglio, T.; Olivero, G.; Merega, E.; Di Prisco, S.; Padolecchia, C.; Grilli, M.; Milanese, M.; Di Cesare Mannelli, L.; Ghelardini, C.; Bonanno, G.; et al. Prophylactic versus therapeutic fingolimod: Restoration of presynaptic defects in mice suffering from experimental autoimmune encephalomyelitis. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [Green Version]
- Grilli, M.; Pittaluga, A.; Merlo-Pich, E.; Marchi, M. NMDA-mediated modulation of dopamine release is modified in rat prefrontal cortex and nucleus accumbens after chronic nicotine treatment. J. Neurochem. 2009. [Google Scholar] [CrossRef]
- Olivero, G.; Grilli, M.; Vergassola, M.; Bonfiglio, T.; Padolecchia, C.; Garrone, B.; Di Giorgio, F.P.; Tongiani, S.; Usai, C.; Marchi, M.; et al. 5-HT2A-mGlu2/3 receptor complex in rat spinal cord glutamatergic nerve endings: A 5-HT2A to mGlu2/3 signalling to amplify presynaptic mechanism of auto-control of glutamate exocytosis. Neuropharmacology 2018, 133, 429–439. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Jolliffe, I.T.T. Principal Component Analysis, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Wold, S.; Esbensen, K.; Geladi, P. Principal component analysis. Chemom. Intell. Lab. Syst. 1987, 2, 37–52. [Google Scholar] [CrossRef]
- Turrini, F.; Donno, D.; Beccaro, G.L.; Zunin, P.; Pittaluga, A.; Boggia, R. Pulsed Ultrasound-Assisted Extraction as an Alternative Method to Conventional Maceration for the Extraction of the Polyphenolic Fraction of Ribes nigrum Buds: A new category of food supplements proposed by the Finnover project. Foods 2019, 8, 466. [Google Scholar] [CrossRef] [Green Version]
- Turrini, F.; Donno, D.; Boggia, R.; Beccaro, G.L.; Zunin, P.; Leardi, R.; Pittaluga, A.M. An innovative green extraction and re-use strategy to valorize food supplement by-products: Castanea sativa bud preparations as case study. Food Res. Int. 2019. [Google Scholar] [CrossRef] [PubMed]
- Boggia, R.; Turrini, F.; Anselmo, M.; Zunin, P.; Donno, D.; Beccaro, G.L. Feasibility of UV–VIS–Fluorescence spectroscopy combined with pattern recognition techniques to authenticate a new category of plant food supplements. J. Food Sci. Technol. 2017. [Google Scholar] [CrossRef]
- Olivero, G.; Turrini, F.; Vergassola, M.; Boggia, R.; Zunin, P.; Donno, D.; Beccaro, G.L.; Grilli, M.; Pittaluga, A. The 3Rs: Reduction and refinement through a multivariate statistical analysis approach in a behavioural study to unveil anxiolytic effects of natural extracts of Tilia tomentosa. Biomed. Sci. Eng. 2020. [Google Scholar] [CrossRef] [Green Version]
- Donno, D.; Boggia, R.; Zunin, P.; Cerutti, A.K.; Guido, M.; Mellano, M.G.; Prgomet, Z.; Beccaro, G.L. Phytochemical fingerprint and chemometrics for natural food preparation pattern recognition: An innovative technique in food supplement quality control. J. Food Sci. Technol. 2016. [Google Scholar] [CrossRef] [Green Version]
- Leussis, M.P.; Bolivar, V.J. Habituation in rodents: A review of behavior, neurobiology, and genetics. Neurosci. Biobehav. Rev. 2006, 30, 1045–1064. [Google Scholar] [CrossRef]
- Takeda, H.; Tsuji, M.; Matsumiya, T. Changes in head-dipping behavior in the hole-board test reflect the anxiogenic and/or anxiolytic state in mice. Eur. J. Pharm. 1998. [Google Scholar] [CrossRef]
- Berridge, C.W.; Dunn, A.J. Restraint-stress-induced changes in exploratory behavior appear to be mediated by norepinephrine-stimulated release of CRF. J. Neurosci. 1989. [Google Scholar] [CrossRef]
- Rodríguez Echandía, E.L.; Broitman, S.T.; Fóscolo, M.R. Effect of the chronic ingestion of chlorimipramine and desipramine on the hole board response to acute stresses in male rats. Pharm. Biochem. Behav. 1987. [Google Scholar] [CrossRef]
- Wang, Y.; Kasper, L.H. The role of microbiome in central nervous system disorders. Brain. Behav. Immun. 2014, 38, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Masana, M.F.; Tyrovolas, S.; Kollia, N.; Chrysohoou, C.; Skoumas, J.; Haro, J.M.; Tousoulis, D.; Papageorgiou, C.; Pitsavos, C.; Panagiotakos, D.B. Dietary Patterns and Their Association with Anxiety Symptoms among Older Adults: The ATTICA Study. Nutrients 2019, 11, 1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Head, K.A.; Kelly, G.S. Nutrients and botanicals for treatment of stress: Adrenal fatigue, neurotransmitter imbalance, anxiety, and restless sleep. Altern. Med. Rev. 2009, 14, 114–140. [Google Scholar] [PubMed]
- Andreescu, C.; Varon, D. New Research on Anxiety Disorders in the Elderly and an Update on Evidence-Based Treatments. Curr. Psychiatry Rep. 2015, 17, 53. [Google Scholar] [CrossRef]
- Andreescu, C.; Lee, S. Anxiety Disorders in the Elderly. In Advances in Experimental Medicine and Biology; Springer: Berlin/Heidelberg, Germany, 2020; pp. 561–576. [Google Scholar]
- Vink, D.; Aartsen, M.J.; Schoevers, R.A. Risk factors for anxiety and depression in the elderly: A review. J. Affect. Disord. 2008, 106, 29–44. [Google Scholar] [CrossRef]
- Kessler, R.C.; Birnbaum, H.G.; Shahly, V.; Bromet, E.; Hwang, I.; McLaughlin, K.A.; Sampson, N.; Andrade, L.H.; de Girolamo, G.; Demyttenaere, K.; et al. Age differences in the prevalence and co-morbidity of DSM-IV major depressive episodes: Results from the WHO World Mental Health Survey Initiative. Depress. Anxiety 2010, 27, 351–364. [Google Scholar] [CrossRef] [Green Version]
- Amico, J.A.; Mantella, R.C.; Vollmer, R.R.; Li, X. Anxiety and stress responses in female oxytocin deficient mice. J. Neuroendocr. 2004. [Google Scholar] [CrossRef]
- Marques, A.A.; Bevilaqua, M.C.D.N.; Da Fonseca, A.M.P.; Nardi, A.E.; Thuret, S.; Dias, G.P. Gender Differences in the Neurobiology of Anxiety: Focus on Adult Hippocampal Neurogenesis. Neural Plast. 2016, 2016, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Puchol, S.; Santafe, A.; Solis, M.; Sanz, V. Properties of four herbal remedies traditionally used to treat anxiety. Basic Clin. Pharm. Toxicol. 2011, 119, 62. [Google Scholar]
- Cavadas, C.; Fontes Ribeiro, C.A.; Santos, M.S.; Cunha, A.P.; Macedo, T.; Caramona, M.M.; Cotrim, M.D. In vitro study of the interaction of Tilia europeae L. aqueous extract with GABA(A) receptors in rat brain. Phyther. Res. 1997. [Google Scholar] [CrossRef]
- Noguerón-Merino, M.C.; Jiménez-Ferrer, E.; Román-Ramos, R.; Zamilpa, A.; Tortoriello, J.; Herrera-Ruiz, M. Interactions of a standardized flavonoid fraction from Tilia americana with Serotoninergic drugs in elevated plus maze. J. Ethnopharmacol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Ortega, G.; Guevara-Fefer, P.; Chávez, M.; Herrera, J.; Martínez, A.L.; González-Trujano, M.E. Sedative and anxiolytic efficacy of Tilia americana var. mexicana inflorescences used traditionally by communities of State of Michoacan, Mexico. J. Ethnopharmacol. 2008. [Google Scholar] [CrossRef]
- Aguirre-Hernández, E.; Martínez, A.L.L.; González-Trujano, M.E.E.; Moreno, J.; Vibrans, H.; Soto-Hernández, M. Pharmacological evaluation of the anxiolytic and sedative effects of Tilia americana L. var. mexicana in mice. J. Ethnopharmacol. 2007, 109, 140–145. [Google Scholar]
- Conley, M.N.; Wong, C.P.; Duyck, K.M.; Hord, N.; Ho, E.; Sharpton, T.J. Aging and serum MCP-1 are associated with gut microbiome composition in a murine model. PeerJ 2016. [Google Scholar] [CrossRef] [Green Version]
- Laukens, D.; Brinkman, B.M.; Raes, J.; De Vos, M.; Vandenabeele, P. Heterogeneity of the gut microbiome in mice: Guidelines for optimizing experimental design. FEMS Microbiol. Rev. 2015, 40, 117–132. [Google Scholar] [CrossRef] [Green Version]
- Vital, M.; Harkema, J.R.; Rizzo, M.; Tiedje, J.; Brandenberger, C. Alterations of the Murine Gut Microbiome with Age and Allergic Airway Disease. J. Immunol. Res. 2015. [Google Scholar] [CrossRef]
- Wax, T.M.; Teena, M.W. Effects of Age, Strain, and Illumination Intensity on Activity and Self-Selection of Light-Dark Schedules in Mice. J. Comp. Physiol. Psychol. 1977. [Google Scholar] [CrossRef]
- Hodge, C.W.; Mehmert, K.K.; Kelley, S.P.; McMahon, T.; Haywood, A.; Olive, M.F.; Wang, D.; Sanchez-Perez, A.M.; Messing, R.O. Supersensitivity to allosteric GABA(A) receptor modulators and alcohol in mice lacking PKCε. Nat. Neurosci. 1999. [Google Scholar] [CrossRef]
- Quoilin, C.; Didone, V.; Tirelli, E.; Quertemont, E. Ontogeny of the stimulant and sedative effects of ethanol in male and female Swiss mice: Gradual changes from weaning to adulthood. Psychopharmacology 2010. [Google Scholar] [CrossRef] [PubMed]
- Yin, M.; Ikejima, K.; Wheeler, M.D.; Bradford, B.U.; Seabra, V.; Forman, D.T.; Sato, N.; Thurman, R.G. Estrogen is involved in early alcohol-induced liver injury in a rat enteral feeding model. Hepatology 2000. [Google Scholar] [CrossRef] [PubMed]
- Iimuro, Y.; Frankenberg, M.V.; Arteel, G.E.; Bradford, B.U.; Wall, C.A.; Thurman, R.G. Female rats exhibit greater susceptibility to early alcohol-induced liver injury than males. Am. J. Physiol. Gastrointest. Liver Physiol. 1997. [Google Scholar] [CrossRef] [PubMed]
- Ikejima, K.; Enomoto, N.; Iimuro, Y.; Ikejima, A.; Fang, D.; Xu, J.; Forman, D.T.; Brenner, D.A.; Thurman, R.G. Estrogen increases sensitivity of hepatic Kupffer cells to endotoxin. Am. J. Physiol. Gastrointest. Liver Physiol. 1998. [Google Scholar] [CrossRef]
- Morgan, M.A.; Pfaff, D.W. Effects of estrogen on activity and fear-related behaviors in mice. Horm. Behav. 2001. [Google Scholar] [CrossRef] [PubMed]
- Shansky, R.M.; Glavis-Bloom, C.; Lerman, D.; McRae, P.; Benson, C.; Miller, K.; Cosand, L.; Horvath, T.L.; Arnsten, A.F.T. Estrogen mediates sex differences in stress-induced prefrontal cortex dysfunction. Mol. Psychiatry 2004. [Google Scholar] [CrossRef]
- Lebron-Milad, K.; Milad, M.R. Sex differences, gonadal hormones and the fear extinction network: Implications for anxiety disorders. Biol. Mood Anxiety Disord. 2012. [Google Scholar] [CrossRef] [Green Version]
- Rai, A.; Gill, M.; Kinra, M.; Shetty, R.; Krishnadas, N.; Rao, C.; Sumalatha, S.; Kumar, N. Catechin ameliorates depressive symptoms in Sprague Dawley rats subjected to chronic unpredictable mild stress by decreasing oxidative stress. Biomed. Rep. 2019. [Google Scholar] [CrossRef] [Green Version]
- Kalshetti, P.; Alluri, R.; Thakurdesai, P. Antidepressant and anti-anxiety effect of ellagic acid from Punica granatum L. rind in olfactory bulbectomy model in rats. Int. J. Pharm. Sci. Rev. Res. 2015, 34, 197–204. [Google Scholar]
- Mansouri, M.T.; Soltani, M.; Naghizadeh, B.; Farbood, Y.; Mashak, A.; Sarkaki, A. A possible mechanism for the anxiolytic-like effect of gallic acid in the rat elevated plus maze. Pharm. Biochem. Behav. 2014, 117, 40–46. [Google Scholar] [CrossRef]
- Lee, B.; Sur, B.; Kwon, S.; Yeom, M.; Shim, I.; Lee, H.; Hahm, D.H. Chronic administration of catechin decreases depression and anxiety-like behaviors in a rat model using chronic corticosterone injections. Biomol. Ther. 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samad, N.; Saleem, A.; Yasmin, F.; Shehzad, M.A. Quercetin protects against stress-induced anxiety- and depression-like behavior and improves memory in male mice. Physiol. Res. 2018, 67, 795–808. [Google Scholar] [CrossRef] [PubMed]
- Bhutada, P.; Mundhada, Y.; Bansod, K.; Ubgade, A.; Quazi, M.; Umathe, S.; Mundhada, D. Reversal by quercetin of corticotrophin releasing factor induced anxiety- and depression-like effect in mice. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2010, 34, 955–960. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Yang, L.; Liu, T.; Wang, J.; Wen, A.; Ding, Y. Ellagic acid protects mice against sleep deprivation-induced memory impairment and anxiety by inhibiting TLR4 and activating Nrf2. Aging 2020. [Google Scholar] [CrossRef]









| Chemical Class | Phytochemical | mg/100 g of Fresh Weight (FW) |
|---|---|---|
| Cinnamic acids | Caffeic acid | n.d. |
| Chlorogenic acid | n.d. | |
| Coumaric acid | n.d. | |
| Ferulic acid | 32.02 ± 2.80 | |
| Flavonols | Hyperoside | 32.98 ± 3.46 |
| Isoquercitrin | 10.45 ± 3.67 | |
| Quercetin | 116.75 ± 4.95 | |
| Quercitrin | 4.38 ± 1.59 | |
| Rutin | 1.78 ± 0.36 | |
| Benzoic acids | Ellagic acid | 440.21 ± 21.02 |
| Gallic acid | 120.40 ± 10.90 | |
| Catechins | (+)-Catechin | 203.56 ± 31.39 |
| (−)-Epicatechin | 220.64 ± 10.03 |
| Animals | Treatment | Water Consumption | Weight | n | ||
|---|---|---|---|---|---|---|
| Start | End | Variation | ||||
| YM | W | 6.38 ± 0.63 | 26.4 ± 0.3 | 27.1 ± 0.3 | (+2.8%) | 15 |
| Vehi | 6.12 ± 0.55 | 27.9 ± 0.5 | 29 ± 0.6 | (+4.3%) | 26 | |
| TTBEs | 6.91 ± 0.26 | 27 ± 0.2 | 28.3 ± 0.3 | (+5%) | 24 | |
| YF | W | 5.94 ± 0.45 | 22 ± 0.2 | 22.5 ± 0.3 | (+2.3%) | 14 |
| Vehi | 5.67 ± 0.38 | 21.7 ± 0.2 | 22.6 ± 0.1 | (+4.1%) | 20 | |
| TTBEs | 5.39 ± 0.58 | 20.5 ± 0.1 | 20.9 ± 0.1 | (+2.2%) | 26 | |
| OM | W | 6.65 ± 0.13 | 29.2 ± 0.5 | 29.2 ± 0.4 | (−0.1%) | 12 |
| Vehi | 6.12 ± 0.48 | 30.6 ± 0.5 | 30.5 ± 0.5 | (−0.2%) | 12 | |
| TTBEs | 6.78 ± 0.19 | 33.6 ± 0.4 | 33.7 ± 0.3 | (+0.3%) | 12 | |
| OF | W | 4.91 ± 0.62 | 25.6 ± 0.5 | 25.8 ± 0.5 | (+0.8%) | 14 |
| Vehi | 4.37 ± 0.52 | 26.5 ± 0.3 | 26.2 ± 0.2 | (−1.4%) | 18 | |
| TTBEs | 3.83 ± 0.33 | 26.7 ± 0.3 | 26.3 ± 0.4 | (−1.7%) | 18 | |
| c1 | c2 | %HD | %AE | %C | tL | T | Av Sp | RAE | D | |
|---|---|---|---|---|---|---|---|---|---|---|
| YMW | Y | M | 4.50 | 38.89 | 22.92 | 36.00 | 6.25 | 7.62 | 87.60 | 2375.00 |
| YMVehi | Y | M | 12.00 | 59.26 | 24.37 | 46.00 | 7.33 | 7.52 | 76.67 | 2350.00 |
| YMTTBEs | Y | M | 17.22 | 83.95 | 18.88 | 56.00 | 10.45 | 11.55 | 84.42 | 3586.00 |
| YFW | Y | F | 5.83 | 50.00 | 10.28 | 21.25 | 3.75 | 9.34 | 79.50 | 2918.00 |
| YFVehi | Y | F | 9.12 | 63.89 | 6.59 | 60.11 | 6.78 | 9.88 | 78.83 | 3074.00 |
| YFTTBEs | Y | F | 19.75 | 69.44 | 5.42 | 50.82 | 9.00 | 6.15 | 70.60 | 1920.00 |
| OMW | O | M | 2.33 | 22.22 | 33.33 | 71.00 | 5.50 | 4.10 | 45.20 | 1124.00 |
| OMVehi | O | M | 3.17 | 38.89 | 47.22 | 51.50 | 4.67 | 4.05 | 48.67 | 1282.00 |
| OMTTBEs | O | M | 11.29 | 66.67 | 25.01 | 59.00 | 4.25 | 6.38 | 43.00 | 2104.00 |
| OFW | O | F | 7.67 | 55.00 | 13.42 | 52.50 | 7.67 | 4.21 | 48.00 | 1224.00 |
| OFVehi | O | F | 13.80 | 75.55 | 9.51 | 30.00 | 3.33 | 4.31 | 47.00 | 1358.00 |
| OFTTBEs | O | F | 11.17 | 70.37 | 7.26 | 35.16 | 3.60 | 4.70 | 46.00 | 1527.00 |
| ZeroYMW | Y | M | 30.41 | 94.95 | 5.73 | 58.69 | 8.23 | 12.54 | 46.75 | 3911.75 |
| ZeroYFW | Y | F | 31.42 | 93.98 | 8.15 | 71.38 | 8.00 | 11.24 | 60.50 | 3508.50 |
| ZeroOMW | O | M | 28.35 | 93.46 | 12.63 | 95.67 | 6.22 | 6.97 | 55.25 | 2210.00 |
| ZeroOFW | O | F | 27.00 | 82.64 | 13.87 | 58.31 | 7.07 | 7.19 | 50.67 | 2309.17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turrini, F.; Vallarino, G.; Cisani, F.; Donno, D.; Beccaro, G.L.; Zunin, P.; Boggia, R.; Pittaluga, A.; Grilli, M. Use of an Animal Model to Evaluate Anxiolytic Effects of Dietary Supplementation with Tilia tomentosa Moench Bud Extracts. Nutrients 2020, 12, 3328. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12113328
Turrini F, Vallarino G, Cisani F, Donno D, Beccaro GL, Zunin P, Boggia R, Pittaluga A, Grilli M. Use of an Animal Model to Evaluate Anxiolytic Effects of Dietary Supplementation with Tilia tomentosa Moench Bud Extracts. Nutrients. 2020; 12(11):3328. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12113328
Chicago/Turabian StyleTurrini, Federica, Giulia Vallarino, Francesca Cisani, Dario Donno, Gabriele Loris Beccaro, Paola Zunin, Raffaella Boggia, Anna Pittaluga, and Massimo Grilli. 2020. "Use of an Animal Model to Evaluate Anxiolytic Effects of Dietary Supplementation with Tilia tomentosa Moench Bud Extracts" Nutrients 12, no. 11: 3328. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12113328