Influence of Glutamine and Branched-Chain Amino Acids Supplementation during Refeeding in Activity-Based Anorectic Mice

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Procedure
2.2. Gastric Emptying
2.3. Body Composition
2.4. Intestinal Permeability
2.5. Protein Extraction and Western Blotting
2.6. Protein Synthesis Analysis by the SUnSET Method
2.7. RT-qPCR
2.8. Plasma Adiponectin and Leptin
2.9. Plasma Biochemical Dosages
2.10. Statistical Analysis
3. Results
3.1. Body Composition and Muscular Protein Synthesis
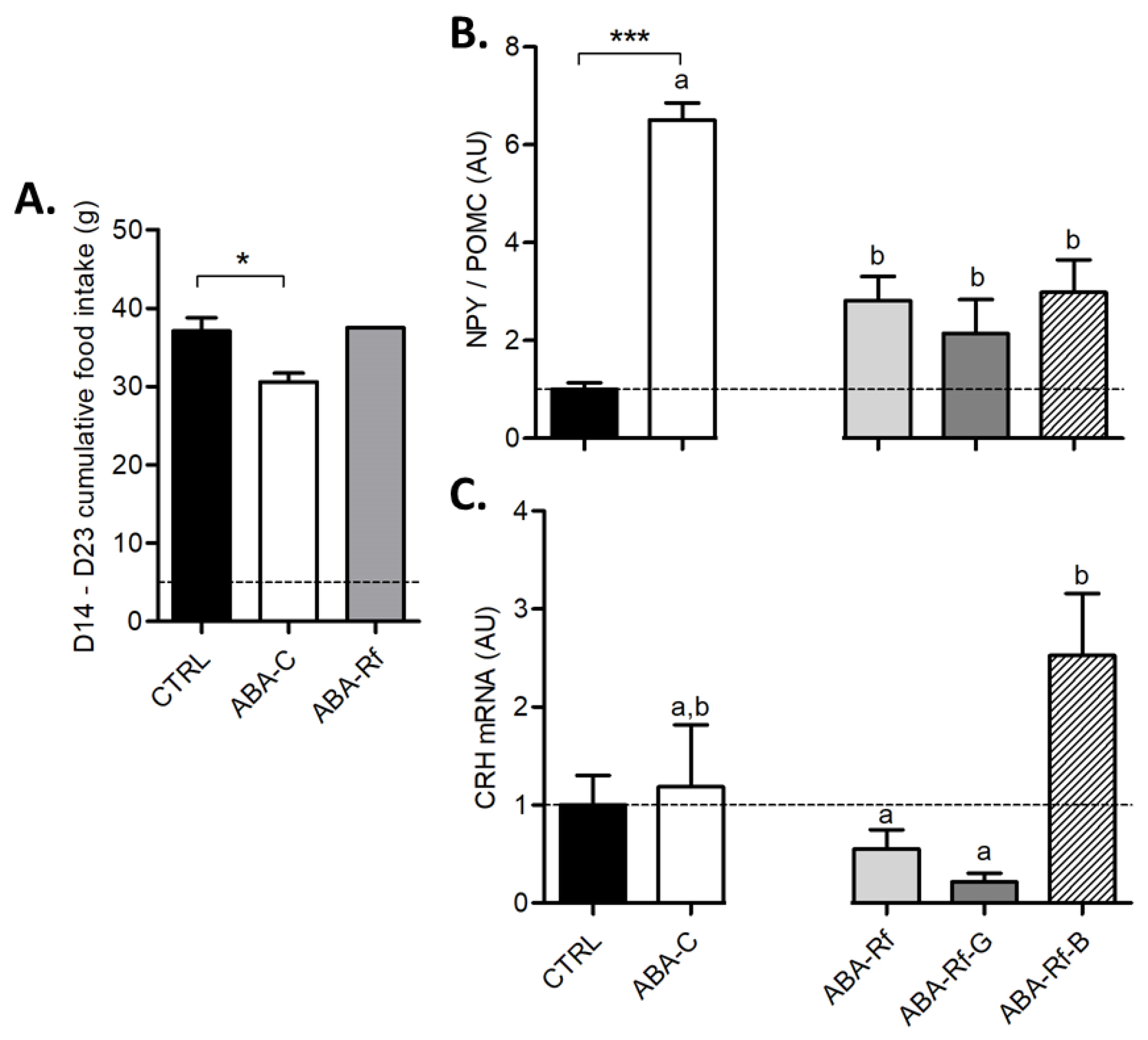
3.2. Gastric Emptying and Hypothalamic Control of Food Intake and Neuropeptides
3.3. Plasma Biochemical Dosages
3.4. Colonic Protein Synthesis, Tight Junction Proteins Expression, and Inflammatory Markers
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- American Psychiatric Association (APA). Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Publishing: Arlington, VA, USA, 2013; ISBN 978-0-89042-554-1. [Google Scholar]
- Volpe, U.; Tortorella, A.; Manchia, M.; Monteleone, A.M.; Albert, U.; Monteleone, P. Eating disorders: What age at onset? Psychiatry Res. 2016, 238, 225–227. [Google Scholar] [CrossRef]
- Galmiche, M.; Déchelotte, P.; Lambert, G.; Tavolacci, M.P. Prevalence of eating disorders over the 2000–2018 period: A systematic literature review. Am. J. Clin. Nutr. 2019, 109, 1402–1413. [Google Scholar] [CrossRef] [PubMed]
- Moskowitz, L.; Weiselberg, E. Anorexia Nervosa/Atypical Anorexia Nervosa. Curr. Probl. Pediatr. Adolesc. Health Care 2017, 47, 70–84. [Google Scholar] [CrossRef] [PubMed]
- Godart, N.T.; Flament, M.F.; Lecrubier, Y.; Jeammet, P. Anxiety disorders in anorexia nervosa and bulimia nervosa: Co-morbidity and chronology of appearance. Eur. Psychiatry 2000, 15, 38–45. [Google Scholar] [CrossRef]
- Lulé, D.; Schulze, U.M.E.; Bauer, K.; Schöll, F.; Müller, S.; Fladung, A.-K.; Uttner, I. Anorexia nervosa and its relation to depression, anxiety, alexithymia and emotional processing deficits. Eat. Weight Disord. Stud. Anorex. Bulim. Obes. 2014, 19, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Rigaud, D.; Pennacchio, H.; Bizeul, C.; Reveillard, V.; Vergès, B. Outcome in AN adult patients: A 13-year follow-up in 484 patients. Diabetes Metab. 2011, 37, 305–311. [Google Scholar] [CrossRef]
- Zipfel, S.; Giel, K.E.; Bulik, C.M.; Hay, P.; Schmidt, U. Anorexia nervosa: Aetiology, assessment, and treatment. Lancet Psychiatry 2015, 2, 1099–1111. [Google Scholar] [CrossRef]
- Berends, T.; Boonstra, N.; Van Elburg, A. Relapse in anorexia nervosa: A systematic review and meta-analysis. Curr. Opin. Psychiatry 2018, 31, 445–455. [Google Scholar] [CrossRef]
- Breton, J.; Déchelotte, P.; Ribet, D. Intestinal microbiota and Anorexia Nervosa. Clin. Nutr. Exp. 2019, 28, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Norris, M.L.; Harrison, M.E.; Isserlin, L.; Robinson, A.; Feder, S.; Sampson, M. Gastrointestinal complications associated with anorexia nervosa: A systematic review: GASTROINTESTINAL COMPLICATIONS IN ANOREXIA NERVOSA. Int. J. Eat. Disord. 2016, 49, 216–237. [Google Scholar] [CrossRef]
- Bargiacchi, A.; Clarke, J.; Paulsen, A.; Leger, J. Refeeding in anorexia nervosa. Eur. J. Pediatr. 2019, 178, 413–422. [Google Scholar] [CrossRef]
- Haas, V.; Kent, D.; Kohn, M.R.; Madden, S.; Clarke, S.; Briody, J.; Fischer, F.; Müller, M.J.; Gaskin, K. Incomplete total body protein recovery in adolescent patients with anorexia nervosa. Am. J. Clin. Nutr. 2018, 107, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Misra, M.; Soyka, L.A.; Miller, K.K.; Grinspoon, S.; Levitsky, L.L.; Klibanski, A. Regional body composition in adolescents with anorexia nervosa and changes with weight recovery. Am. J. Clin. Nutr. 2003, 77, 1361–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaye, W.H.; Bailer, U.F.; Frank, G.K.; Wagner, A.; Henry, S.E. Brain imaging of serotonin after recovery from anorexia and bulimia nervosa. Physiol. Behav. 2005, 86, 15–17. [Google Scholar] [CrossRef] [PubMed]
- Mack, I.; Cuntz, U.; Grämer, C.; Niedermaier, S.; Pohl, C.; Schwiertz, A.; Zimmermann, K.; Zipfel, S.; Enck, P.; Penders, J. Weight gain in anorexia nervosa does not ameliorate the faecal microbiota, branched chain fatty acid profiles, and gastrointestinal complaints. Sci. Rep. 2016, 6, 26752. [Google Scholar] [CrossRef] [Green Version]
- Achamrah, N.; Coëffier, M.; Déchelotte, P. Physical activity in patients with anorexia nervosa. Nutr. Rev. 2016, 74, 301–311. [Google Scholar] [CrossRef]
- Nobis, S.; Morin, A.; Achamrah, N.; Belmonte, L.; Legrand, R.; Chan, P.; do Rego, J.-L.; Vaudry, D.; Gourcerol, G.; Déchelotte, P.; et al. Delayed gastric emptying and altered antrum protein metabolism during activity-based anorexia. Neurogastroenterol. Motil. 2018, 30, e13305. [Google Scholar] [CrossRef]
- Achamrah, N.; Déchelotte, P.; Coëffier, M. Glutamine and the regulation of intestinal permeability: From bench to bedside. Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 86–91. [Google Scholar] [CrossRef]
- Fouré, A.; Bendahan, D. Is Branched-Chain Amino Acids Supplementation an Efficient Nutritional Strategy to Alleviate Skeletal Muscle Damage? A Systematic Review. Nutrients 2017, 9, 1047. [Google Scholar] [CrossRef] [Green Version]
- Platt, K.M.; Charnigo, R.J.; Shertzer, H.G.; Pearson, K.J. Branched-Chain Amino Acid Supplementation in Combination with Voluntary Running Improves Body Composition in Female C57BL/6 Mice. J. Diet. Suppl. 2016, 13, 473–486. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.-H.; Kim, S.-H.; Jeong, W.-S.; Lee, H.-Y. Effect of BCAA intake during endurance exercises on fatigue substances, muscle damage substances, and energy metabolism substances. J. Exerc. Nutr. Biochem. 2013, 17, 169–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahimi, M.H.; Shab-Bidar, S.; Mollahosseini, M.; Djafarian, K. Branched-chain amino acid supplementation and exercise-induced muscle damage in exercise recovery: A meta-analysis of randomized clinical trials. Nutrition 2017, 42, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Fujita, S.; Volpi, E. Amino Acids and Muscle Loss with Aging. J. Nutr. 2006, 136, 277S–280S. [Google Scholar] [CrossRef] [PubMed]
- Achamrah, N.; Nobis, S.; Breton, J.; Jésus, P.; Belmonte, L.; Maurer, B.; Legrand, R.; Bôle-Feysot, C.; do Rego, J.L.; Goichon, A.; et al. Maintaining physical activity during refeeding improves body composition, intestinal hyperpermeability and behavior in anorectic mice. Sci. Rep. 2016, 6, 21887. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Pierre, J.F.; Heneghan, A.F.; Busch, R.; Kudsk, K.A. Glutamine improves innate immunity and prevents bacterial enteroinvasion during parenteral nutrition. J. Parenter. Enter. Nutr. 2015, 39, 688–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertrand, J.; Goichon, A.; Déchelotte, P.; Coëffier, M. Regulation of intestinal protein metabolism by amino acids. Amino Acids 2013, 45, 443–450. [Google Scholar] [CrossRef]
- Coëffier, M. Modulating effect of glutamine on IL-1β-induced cytokine production by human gut. Clin. Nutr. 2003, 22, 407–413. [Google Scholar] [CrossRef]
- Zhou, Q.; Verne, M.L.; Fields, J.Z.; Lefante, J.J.; Basra, S.; Salameh, H.; Verne, G.N. Randomised placebo-controlled trial of dietary glutamine supplements for postinfectious irritable bowel syndrome. Gut 2018, Published Online First. 1–7. [Google Scholar] [CrossRef]
- L’Huillier, C.; Jarbeau, M.; Achamrah, N.; Belmonte, L.; Amamou, A.; Nobis, S.; Goichon, A.; Salameh, E.; Bahlouli, W.; do Rego, J.-L.; et al. Glutamine, but not Branched-Chain Amino Acids, Restores Intestinal Barrier Function during Activity-Based Anorexia. Nutrients 2019, 11, 1348. [Google Scholar] [CrossRef] [Green Version]
- Jésus, P.; Ouelaa, W.; François, M.; Riachy, L.; Guérin, C.; Aziz, M.; Do Rego, J.-C.; Déchelotte, P.; Fetissov, S.O.; Coëffier, M. Alteration of intestinal barrier function during activity-based anorexia in mice. Clin. Nutr. 2014, 33, 1046–1053. [Google Scholar] [CrossRef]
- Belmonte, L.; Achamrah, N.; Nobis, S.; Guérin, C.; Riou, G.; Bôle-Feysot, C.; Boyer, O.; Richard, V.; Rego, J.C.D.; Déchelotte, P.; et al. A role for intestinal TLR4-driven inflammatory response during activity-based anorexia. Sci. Rep. 2016, 6, 35813. [Google Scholar] [CrossRef] [PubMed]
- Coëffier, M.; Miralles-Barrachina, O.; Le Pessot, F.; Lalaude, O.; Daveau, M.; Lavoinne, A.; Lerebours, E.; Déchelotte, P. INFLUENCE OF GLUTAMINE ON CYTOKINE PRODUCTION BY HUMAN GUT IN VITRO. Cytokine 2001, 13, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Nobis, S.; Goichon, A.; Achamrah, N.; Guérin, C.; Azhar, S.; Chan, P.; Morin, A.; Bôle-Feysot, C.; do Rego, J.C.; Vaudry, D.; et al. Alterations of proteome, mitochondrial dynamic and autophagy in the hypothalamus during activity-based anorexia. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nobis, S.; Achamrah, N.; Goichon, A.; L’Huillier, C.; Morin, A.; Guérin, C.; Chan, P.; do Rego, J.L.; do Rego, J.C.; Vaudry, D.; et al. Colonic Mucosal Proteome Signature Reveals Reduced Energy Metabolism and Protein Synthesis but Activated Autophagy during Anorexia-Induced Malnutrition in Mice. Proteomics 2018, 18, 1700395. [Google Scholar] [CrossRef]
- Heruc, G.A.; Little, T.J.; Kohn, M.; Madden, S.; Clarke, S.; Horowitz, M.; Feinle-Bisset, C. Disturbed in Adolescent Females with Anorexia Nervosa and Only Partially Restored with Short-Term Refeeding. Nutrients 2018, 11, 59. [Google Scholar] [CrossRef] [Green Version]
- Dinan, T.G.; Cryan, J.F. Brain-Gut-Microbiota Axis and Mental Health. Psychosom. Med. 2017, 79, 920–926. [Google Scholar] [CrossRef]
- Bindels, L.B.; Neyrinck, A.M.; Loumaye, A.; Catry, E.; Walgrave, H.; Cherbuy, C.; Leclercq, S.; Van Hul, M.; Plovier, H.; Pachikian, B.; et al. Increased gut permeability in cancer cachexia: Mechanisms and clinical relevance. Oncotarget 2018, 9, 18224–18238. [Google Scholar] [CrossRef] [Green Version]
- Orphanidou, C.I.; McCargar, L.J.; Birmingham, C.L.; Belzberg, A.S. Changes in body composition and fat distribution after short-term weight gain in patients with anorexia nervosa. Am. J. Clin. Nutr. 1997, 65, 1034–1041. [Google Scholar] [CrossRef] [Green Version]
- Bluemel, S.; Menne, D.; Milos, G.; Goetze, O.; Fried, M.; Schwizer, W.; Fox, M.; Steingoetter, A. Relationship of body weight with gastrointestinal motor and sensory function: Studies in anorexia nervosa and obesity. BMC Gastroenterol. 2017, 17, 4. [Google Scholar] [CrossRef] [Green Version]
- Zgheib, S.; Méquinion, M.; Lucas, S.; Leterme, D.; Ghali, O.; Tolle, V.; Zizzari, P.; Bellefontaine, N.; Legroux-Gérot, I.; Hardouin, P.; et al. Long-Term Physiological Alterations and Recovery in a Mouse Model of Separation Associated with Time-Restricted Feeding: A Tool to Study Anorexia Nervosa Related Consequences. PLoS ONE 2014, 9, e103775. [Google Scholar] [CrossRef] [Green Version]
- Haluzík, M.; Papezová, H.; Nedvídková, J.; Kábrt, J. Serum leptin levels in patients with anorexia nervosa before and after partial refeeding, relationships to serum lipids and biochemical nutritional parameters. Physiol. Res. 1999, 48, 197–202. [Google Scholar] [PubMed]
- Ruscica, M.; Macchi, C.; Gandini, S.; Morlotti, B.; Erzegovesi, S.; Bellodi, L.; Magni, P. Free and bound plasma leptin in anorexia nervosa patients during a refeeding program. Endocrine 2016, 51, 380–383. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, J.P.; Valéro, R.; Maraninchi, M.; Lorec, A.M.; Samuelian-Massat, C.; Bégu-Le Corroller, A.; Nicolay, A.; Gaudart, J.; Portugal, H.; Vialettes, B. Growth hormone level at admission and its evolution during refeeding are predictive of short-term outcome in restrictive anorexia nervosa. Br. J. Nutr. 2013, 109, 2175–2181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holeček, M. Branched-chain amino acids in health and disease: Metabolism, alterations in blood plasma, and as supplements. Nutr. Metab. (Lond) 2018, 15, 332018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeanandin, G.; Balage, M.; Schneider, S.M.; Dupont, J.; Hébuterne, X.; Mothe-Satney, I.; Dardevet, D. Differential effect of long-term leucine supplementation on skeletal muscle and adipose tissue in old rats: An insulin signaling pathway approach. Age (Dordr) 2012, 34, 371–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vianna, D.; Resende, G.F.T.; Torres-Leal, F.L.; Pantaleão, L.C.; Donato, J.; Tirapegui, J. Long-term leucine supplementation reduces fat mass gain without changing body protein status of aging rats. Nutrition 2012, 28, 182–189. [Google Scholar] [CrossRef]
- Kitajima, Y.; Takahashi, H.; Akiyama, T.; Murayama, K.; Iwane, S.; Kuwashiro, T.; Tanaka, K.; Kawazoe, S.; Ono, N.; Eguchi, T.; et al. Supplementation with branched-chain amino acids ameliorates hypoalbuminemia, prevents sarcopenia, and reduces fat accumulation in the skeletal muscles of patients with liver cirrhosis. J. Gastroenterol. 2018, 53, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Ban, H.; Shigemitsu, K.; Yamatsuji, T.; Haisa, M.; Nakajo, T.; Takaoka, M.; Nobuhisa, T.; Gunduz, M.; Tanaka, N.; Naomoto, Y. Arginine and Leucine regulate p70 S6 kinase and 4E-BP1 in intestinal epithelial cells. Int. J. Mol. Med. 2004, 13, 537–543. [Google Scholar] [CrossRef] [Green Version]
- Coëffier, M.; Claeyssens, S.; Bensifi, M.; Lecleire, S.; Boukhettala, N.; Maurer, B.; Donnadieu, N.; Lavoinne, A.; Cailleux, A.-F.; Déchelotte, P. Influence of leucine on protein metabolism, phosphokinase expression, and cell proliferation in human duodenum. Am. J. Clin. Nutr. 2011, 93, 1255–1262. [Google Scholar] [CrossRef]
- Achamrah, N.; Nobis, S.; Goichon, A.; Breton, J.; Legrand, R.; do Rego, J.L.; do Rego, J.C.; Déchelotte, P.; Fetissov, S.O.; Belmonte, L.; et al. Sex differences in response to activity-based anorexia model in C57Bl/6 mice. Physiol. Behav. 2017, 170, 1–5. [Google Scholar] [CrossRef]
- Solmi, M.; Collantoni, E.; Meneguzzo, P.; Tenconi, E.; Favaro, A. Network analysis of specific psychopathology and psychiatric symptoms in patients with anorexia nervosa. Eur. Eat. Disord. Rev. 2019, 27, 24–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
L’Huillier, C.; Jarbeau, M.; Pingeon, F.; Bahlouli, W.; Salameh, E.; Amamou, A.; Goichon, A.; Tirelle, P.; do Rego, J.-L.; Déchelotte, P.; et al. Influence of Glutamine and Branched-Chain Amino Acids Supplementation during Refeeding in Activity-Based Anorectic Mice. Nutrients 2020, 12, 3510. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12113510
L’Huillier C, Jarbeau M, Pingeon F, Bahlouli W, Salameh E, Amamou A, Goichon A, Tirelle P, do Rego J-L, Déchelotte P, et al. Influence of Glutamine and Branched-Chain Amino Acids Supplementation during Refeeding in Activity-Based Anorectic Mice. Nutrients. 2020; 12(11):3510. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12113510
Chicago/Turabian StyleL’Huillier, Clément, Marine Jarbeau, Floriane Pingeon, Wafa Bahlouli, Emmeline Salameh, Asma Amamou, Alexis Goichon, Pauline Tirelle, Jean-Luc do Rego, Pierre Déchelotte, and et al. 2020. "Influence of Glutamine and Branched-Chain Amino Acids Supplementation during Refeeding in Activity-Based Anorectic Mice" Nutrients 12, no. 11: 3510. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12113510