Functional Biomarkers for the Selenium Status in a Human Nutritional Intervention Study


 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Study Design and Diet
- 1
- Adequate intake of energy, carbohydrates, protein and fat according to the guidelines of the German Society of Nutrition [31];
- 2
- Desirable intake of saturated fatty acids (SFA, ≤ 7 % of daily energy), monounsaturated fatty acids (MUFA, ≥ 10 % of daily energy), polyunsaturated fatty acids (PUFA, ≥ 10 % of daily energy) and long-chain n-3 PUFA (≥ 500 mg/d);
- 3
- Encouraged consumption of vegetables, fruits and cereals;
- 4
- Intake of > 40 g/d dietary fiber,;
- 5
- Reduced intake of salt and sugar reduction;
- 6
- Reduced intake of (highly) processed, calorie-rich, nutrient-poor foods (e.g., fast food, convenience products);
- 7
- Optimized intake of vitamins, minerals and trace elements by commercially available foods; and
- 8
- Encouraged physical activity.
2.3. Laboratory Measurements
2.3.1. Total Selenium
2.3.2. GPx Activity
2.3.3. SELENOP
2.4. Statistical Analysis
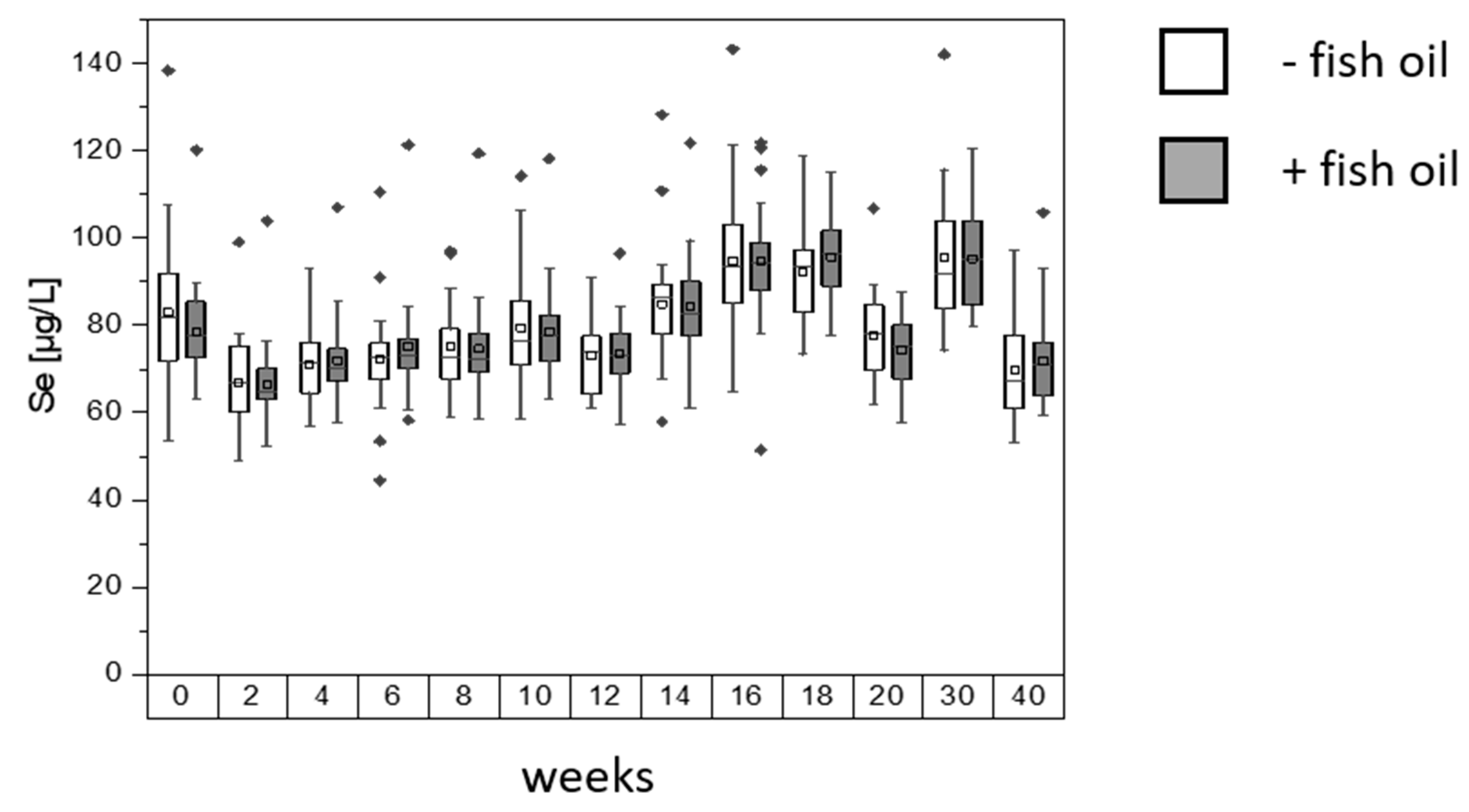
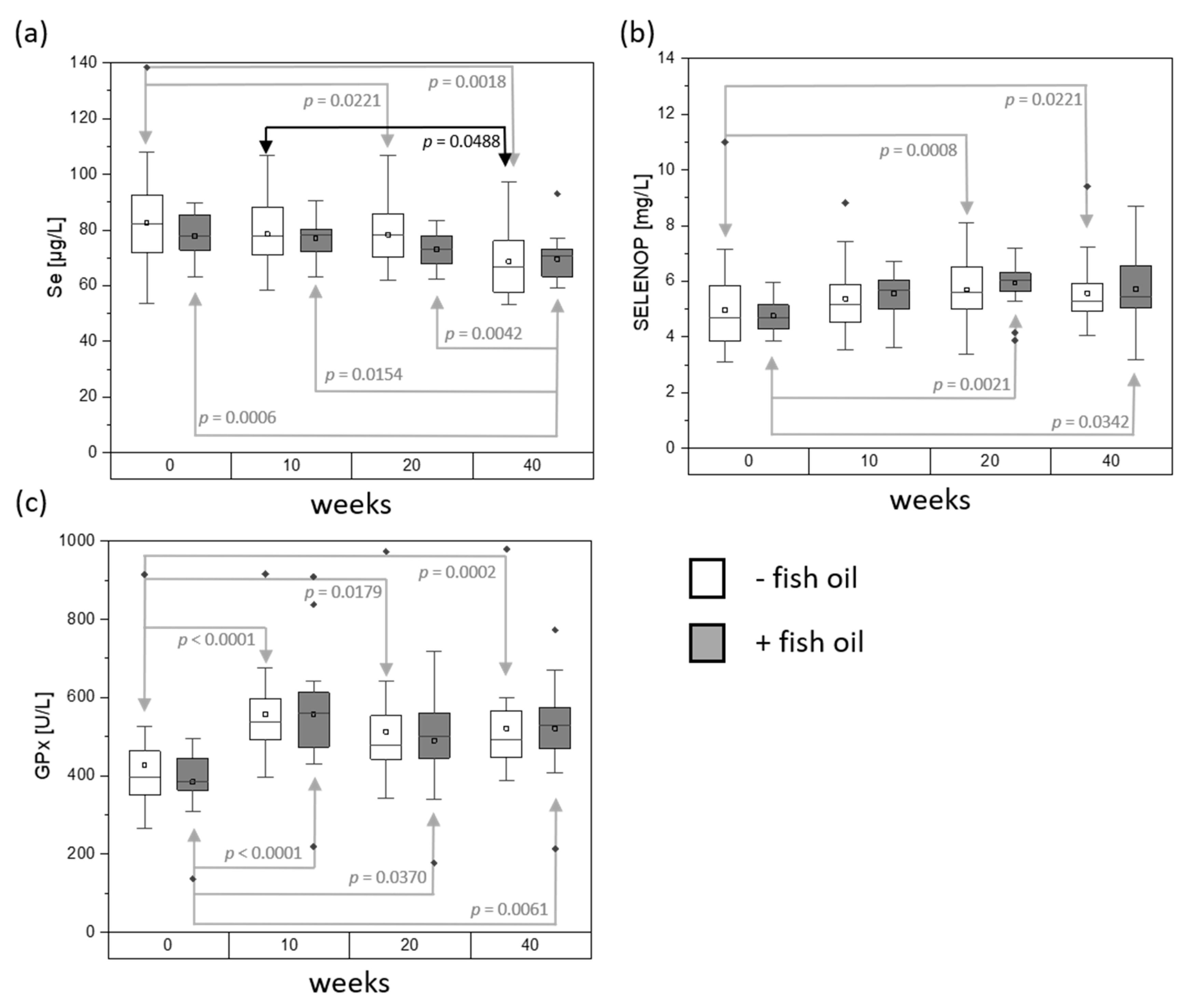
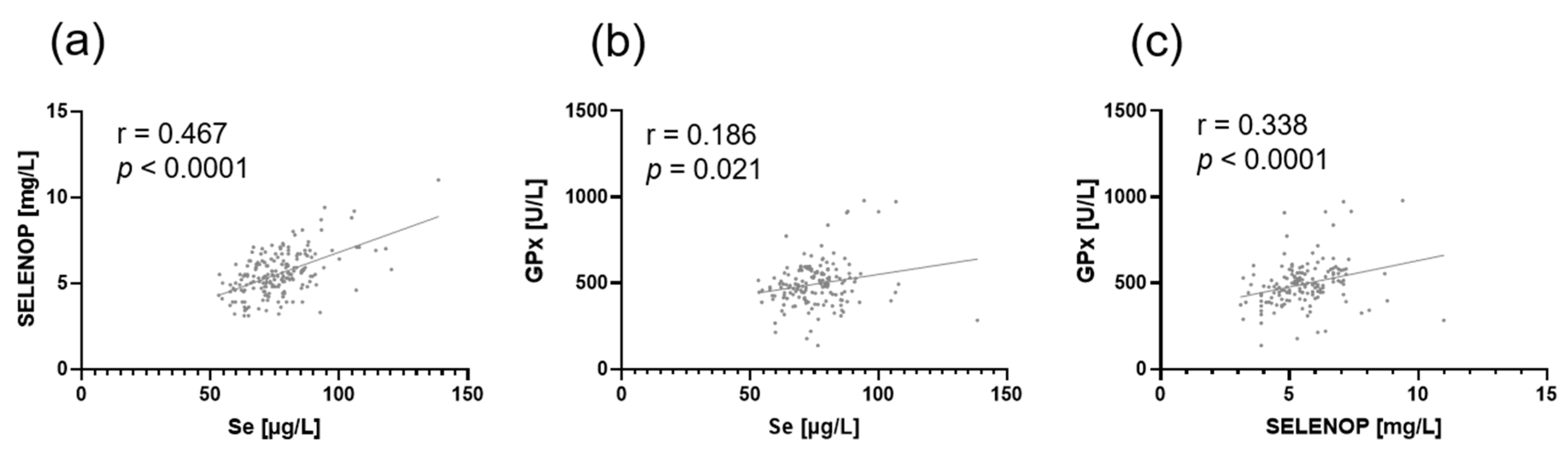
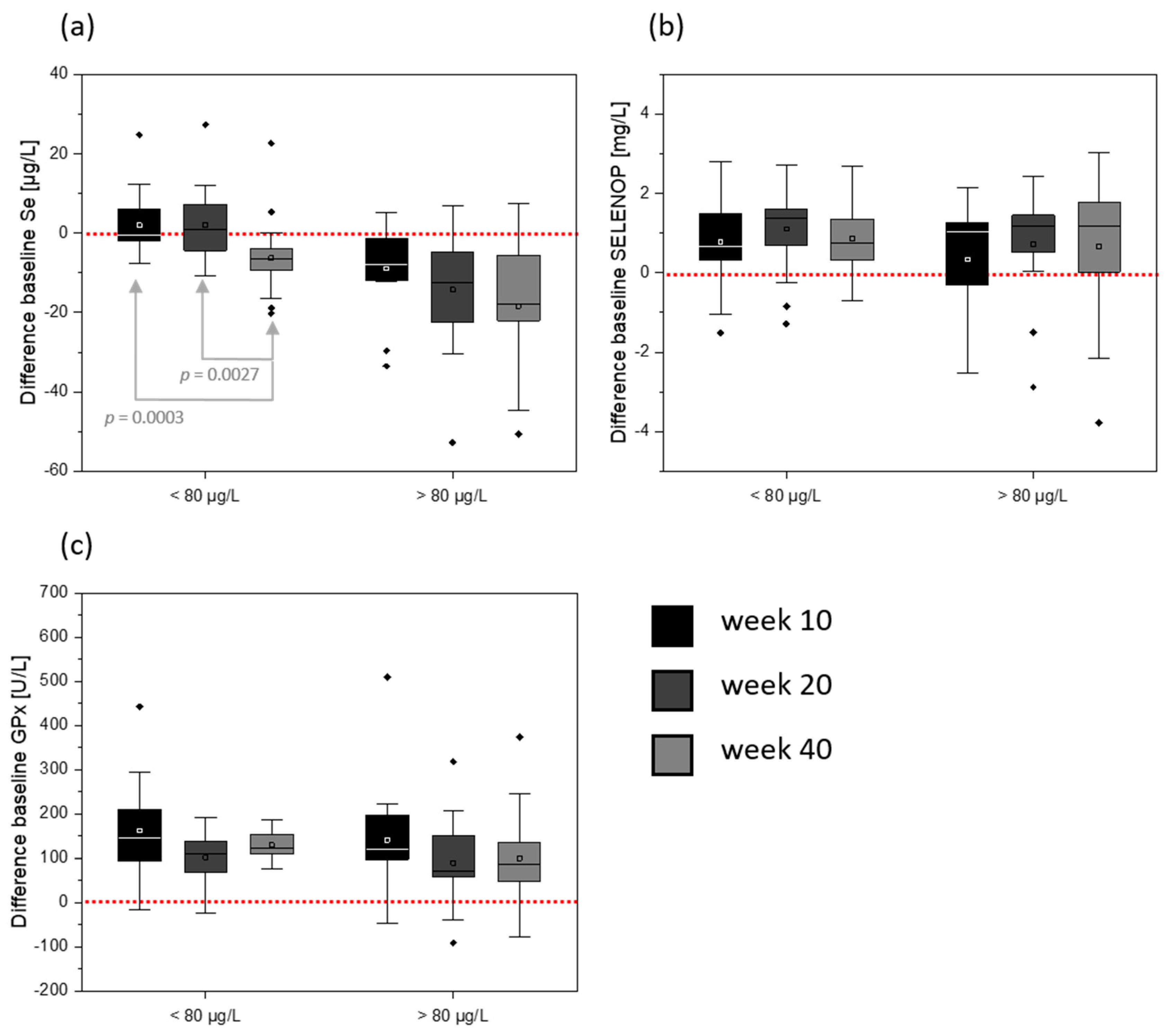
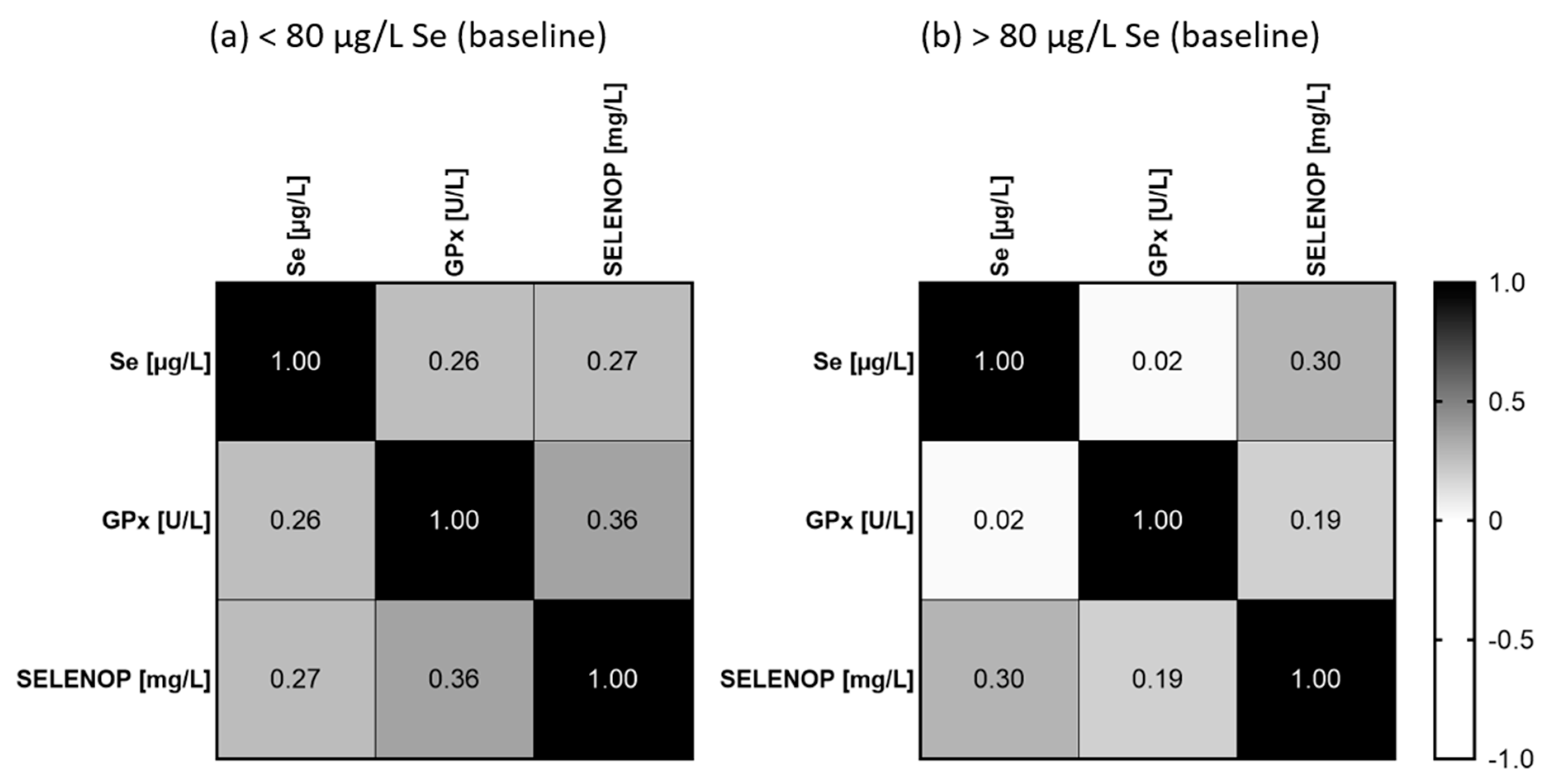
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

References
- Köhrle, J. The deiodinase family: Selenoenzymes regulating thyroid hormone availability and action. Cell. Mol. Life Sci. CMLS 2000, 57, 1853–1863. [Google Scholar] [CrossRef] [PubMed]
- Hatfield, D.L.; Gladyshev, V.N. How selenium has altered our understanding of the genetic code. Mol. Cell Biol. 2002, 22, 3565–3576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arner, E.S.; Holmgren, A. Physiological functions of thioredoxin and thioredoxin reductase. Eur. J. Biochem. 2000, 267, 6102–6109. [Google Scholar] [CrossRef] [PubMed]
- Combs, G.F., Jr. Biomarkers of selenium status. Nutrients 2015, 7, 2209–2236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Combs, G.F., Jr. Selenium in global food systems. Br. J. Nutr. 2001, 85, 517–547. [Google Scholar] [CrossRef] [Green Version]
- Brown, K.; Arthur, J. Selenium, selenoproteins and human health: A review. Public Health Nutr. 2001, 4, 593–599. [Google Scholar] [CrossRef] [Green Version]
- Bornhorst, J.; Kipp, A.P.; Haase, H.; Meyer, S.; Schwerdtle, T. The crux of inept biomarkers for risks and benefits of trace elements. Trac-Trend Anal. Chem. 2018, 104, 183–190. [Google Scholar] [CrossRef]
- Health, N.I.O. Selenium—Fact Sheet for Health Professionals. Available online: https://ods.od.nih.gov/factsheets/Selenium-HealthProfessional/ (accessed on 20 January 2020).
- Xia, Y.; Hill, K.E.; Byrne, D.W.; Xu, J.; Burk, R.F. Effectiveness of selenium supplements in a low-selenium area of china. Am. J. Clin. Nutr. 2005, 81, 829–834. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Hill, K.E.; Li, P.; Xu, J.; Zhou, D.; Motley, A.K.; Wang, L.; Byrne, D.W.; Burk, R.F. Optimization of selenoprotein p and other plasma selenium biomarkers for the assessment of the selenium nutritional requirement: A placebo-controlled, double-blind study of selenomethionine supplementation in selenium-deficient chinese subjects. Am. J. Clin. Nutr. 2010, 92, 525–531. [Google Scholar] [CrossRef] [Green Version]
- Ravn-Haren, G.; Krath, B.N.; Overvad, K.; Cold, S.; Moesgaard, S.; Larsen, E.H.; Dragsted, L.O. Effect of long-term selenium yeast intervention on activity and gene expression of antioxidant and xenobiotic metabolising enzymes in healthy elderly volunteers from the danish prevention of cancer by intervention by selenium (precise) pilot study. Br. J. Nutr. 2008, 99, 1190–1198. [Google Scholar] [CrossRef] [Green Version]
- Burk, R.F.; Norsworthy, B.K.; Hill, K.E.; Motley, A.K.; Byrne, D.W. Effects of chemical form of selenium on plasma biomarkers in a high-dose human supplementation trial. Cancer Epidemiol. Biomark. Prev. 2006, 15, 804–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.G.; Hill, K.E.; Burk, R.F. Dietary selenium intake controls rat plasma selenoprotein p concentration. J. Nutr. 1989, 119, 1010–1012. [Google Scholar] [CrossRef] [PubMed]
- Burk, R.F.; Hill, K.E. Selenoprotein p-expression, functions, and roles in mammals. Biochim. Biophys. Acta 2009, 1790, 1441–1447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Read, R.; Bellew, T.; Yang, J.G.; Hill, K.E.; Palmer, I.S.; Burk, R.F. Selenium and amino acid composition of selenoprotein p, the major selenoprotein in rat serum. J. Biol. Chem. 1990, 265, 17899–17905. [Google Scholar] [PubMed]
- Burk, R.F.; Hill, K.E. Regulation of selenoproteins. Annu. Rev. Nutr. 1993, 13, 65–81. [Google Scholar] [CrossRef]
- Hagmar, L.; Persson-Moschos, M.; Akesson, B.; Schutz, A. Plasma levels of selenium, selenoprotein p and glutathione peroxidase and their correlations to fish intake and serum levels of thyrotropin and thyroid hormones: A study on latvian fish consumers. Eur. J. Clin. Nutr. 1998, 52, 796–800. [Google Scholar] [CrossRef] [Green Version]
- Hill, K.E.; Xia, Y.; Akesson, B.; Boeglin, M.E.; Burk, R.F. Selenoprotein p concentration in plasma is an index of selenium status in selenium-deficient and selenium-supplemented chinese subjects. J. Nutr. 1996, 126, 138–145. [Google Scholar] [CrossRef] [Green Version]
- Hambidge, M. Biomarkers of trace mineral intake and status. J. Nutr. 2003, 133, 948S–955S. [Google Scholar] [CrossRef]
- Goen, T.; Schaller, B.; Jager, T.; Brau-Dumler, C.; Schaller, K.H.; Drexler, H. Biological monitoring of exposure and effects in workers employed in a selenium-processing plant. Int. Arch. Occup. Environ. Health 2015, 88, 623–630. [Google Scholar] [CrossRef]
- Greiner, A.; Goen, T.; Hildebrand, J.; Feltes, R.; Drexler, H. Low internal exposure and absence of adverse effects in workers exposed to high air levels of inorganic selenium. Toxicol. Lett. 2018, 298, 141–149. [Google Scholar] [CrossRef]
- Chen, X.; Yang, G.; Chen, J.; Chen, X.; Wen, Z.; Ge, K. Studies on the relations of selenium and keshan disease. Biol. Trace Elem. Res. 1980, 2, 91–107. [Google Scholar] [CrossRef] [PubMed]
- Ge, K.; Yang, G. The epidemiology of selenium deficiency in the etiological study of endemic diseases in china. Am. J. Clin. Nutr. 1993, 57, 259S–263S. [Google Scholar] [CrossRef] [PubMed]
- Hurst, R.; Siyame, E.W.; Young, S.D.; Chilimba, A.D.; Joy, E.J.; Black, C.R.; Ander, E.L.; Watts, M.J.; Chilima, B.; Gondwe, J.; et al. Soil-type influences human selenium status and underlies widespread selenium deficiency risks in malawi. Sci. Rep. 2013, 3, 1425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fradejas-Villar, N. Consequences of mutations and inborn errors of selenoprotein biosynthesis and functions. Free Radic. Biol. Med. 2018, 127, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.J.; Duarte-Salles, T.; Hybsier, S.; Trichopoulou, A.; Stepien, M.; Aleksandrova, K.; Overvad, K.; Tjonneland, A.; Olsen, A.; Affret, A.; et al. Prediagnostic selenium status and hepatobiliary cancer risk in the european prospective investigation into cancer and nutrition cohort. Am. J. Clin. Nutr. 2016, 104, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.J.; Fedirko, V.; Jenab, M.; Schomburg, L.; Meplan, C.; Freisling, H.; Bueno-de-Mesquita, H.B.; Hybsier, S.; Becker, N.P.; Czuban, M.; et al. Selenium status is associated with colorectal cancer risk in the european prospective investigation of cancer and nutrition cohort. Int. J. Cancer 2015, 136, 1149–1161. [Google Scholar] [CrossRef] [PubMed]
- Benstoem, C.; Goetzenich, A.; Kraemer, S.; Borosch, S.; Manzanares, W.; Hardy, G.; Stoppe, C. Selenium and its supplementation in cardiovascular disease—What do we know? Nutrients 2015, 7, 3094–3118. [Google Scholar] [CrossRef] [Green Version]
- Bomer, N.; Beverborg, N.G.; Hoes, M.F.; Streng, K.W.; Vermeer, M.; Dokter, M.M.; IJmker, J.; Anker, S.D.; Cleland, J.G.F.; Hillege, H.L.; et al. Selenium and outcome in heart failure. Eur. J. Heart Fail. 2019. [Google Scholar] [CrossRef] [Green Version]
- Schomburg, L.; Orho-Melander, M.; Struck, J.; Bergmann, A.; Melander, O. Selenoprotein-p deficiency predicts cardiovascular disease and death. Nutrients 2019, 11, 1852. [Google Scholar] [CrossRef] [Green Version]
- Deutsche Gesellschaft für Ernährung, Österreichische Gesellschaft für Ernärhung; Schweizerische Gesellschaft für Ernährungsforschung, Schweizerische Vereinigung für Ernährung. Referenzwerte für die Nährstoffzufuhr; Deutsche Gesellschaft für Ernährung: Bonn, Germany, 2019. [Google Scholar]
- Baudry, J.; Kopp, J.F.; Boeing, H.; Kipp, A.P.; Schwerdtle, T.; Schulze, M.B. Changes of trace element status during aging: Results of the epic-potsdam cohort study. Eur. J. Nutr. 2019. [Google Scholar] [CrossRef] [Green Version]
- Florian, S.; Krehl, S.; Loewinger, M.; Kipp, A.; Banning, A.; Esworthy, S.; Chu, F.F.; Brigelius-Flohe, R. Loss of gpx2 increases apoptosis, mitosis, and gpx1 expression in the intestine of mice. Free Radic. Biol. Med. 2010, 49, 1694–1702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravn-Haren, G.; Olsen, A.; Tjønneland, A.; Dragsted, L.O.; Nexø, B.A.; Wallin, H.; Overvad, K.; Raaschou-Nielsen, O.; Vogel, U. Associations between gpx1 pro198leu polymorphism, erythrocyte gpx activity, alcohol consumption and breast cancer risk in a prospective cohort study. Carcinogenesis 2005, 27, 820–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forsberg, L.; de Faire, U.; Marklund, S.L.; Andersson, P.M.; Stegmayr, B.; Morgenstern, R. Phenotype determination of a common pro-leu polymorphism in human glutathione peroxidase 1. Blood Cells Mol. Dis. 2000, 26, 423–426. [Google Scholar] [CrossRef] [PubMed]
- Arsova-Sarafinovska, Z.; Matevska, N.; Eken, A.; Petrovski, D.; Banev, S.; Dzikova, S.; Georgiev, V.; Sikole, A.; Erdem, O.; Sayal, A.; et al. Glutathione peroxidase 1 (gpx1) genetic polymorphism, erythrocyte gpx activity, and prostate cancer risk. Int. Urol. Nephrol. 2008, 41, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voetsch, B.; Jin, R.C.; Bierl, C.; Benke, K.S.; Kenet, G.; Simioni, P.; Ottaviano, F.; Damasceno, B.P.; Annichino-Bizacchi, J.M.; Handy, D.E.; et al. Promoter polymorphisms in the plasma glutathione peroxidase gpx-3 gene. Stroke 2007, 38, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Letsiou, S.; Nomikos, T.; Panagiotakos, D.B.; Pergantis, S.A.; Fragopoulou, E.; Pitsavos, C.; Stefanadis, C.; Antonopoulou, S. Gender-specific distribution of selenium to serum selenoproteins: Associations with total selenium levels, age, smoking, body mass index, and physical activity. BioFactors 2014, 40, 524–535. [Google Scholar] [CrossRef]
- Olson, G.E.; Winfrey, V.P.; Hill, K.E.; Burk, R.F. Megalin mediates selenoprotein p uptake by kidney proximal tubule epithelial cells. J. Biol. Chem. 2008, 283, 6854–6860. [Google Scholar] [CrossRef] [Green Version]
- Méplan, C.; Crosley, L.K.; Nicol, F.; Beckett, G.J.; Howie, A.F.; Hill, K.E.; Horgan, G.; Mathers, J.C.; Arthur, J.R.; Hesketh, J.E. Genetic polymorphisms in the human selenoprotein p gene determine the response of selenoprotein markers to selenium supplementation in a gender-specific manner (the selgen study). FASEB J. 2007, 21, 3063–3074. [Google Scholar] [CrossRef] [Green Version]
- Lundholm, L.; Putnik, M.; Otsuki, M.; Andersson, S.; Ohlsson, C.; Gustafsson, J.A.; Dahlman-Wright, K. Effects of estrogen on gene expression profiles in mouse hypothalamus and white adipose tissue: Target genes include glutathione peroxidase 3 and cell death-inducing DNA fragmentation factor, alpha-subunit-like effector a. J. Endocrinol. 2008, 196, 547–557. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.S.; Kim, A.Y.; Choi, J.W.; Kim, M.; Yasue, S.; Son, H.J.; Masuzaki, H.; Park, K.S.; Kim, J.B. Dysregulation of adipose glutathione peroxidase 3 in obesity contributes to local and systemic oxidative stress. Mol. Endocrinol. 2008, 22, 2176–2189. [Google Scholar] [CrossRef] [Green Version]
- Castro, A.M.; Macedo-de la Concha, L.E.; Pantoja-Meléndez, C.A. Low-grade inflammation and its relation to obesity and chronic degenerative diseases. Revista Médica del Hospital General de México 2017, 80, 101–105. [Google Scholar] [CrossRef]
- Larvie, D.Y.; Doherty, J.L.; Donati, G.L.; Armah, S.M. Relationship between selenium and hematological markers in young adults with normal weight or overweight/obesity. Antioxidants (Basel) 2019, 8, 463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burk, R.F.; Hill, K.E.; Motley, A.K.; Winfrey, V.P.; Kurokawa, S.; Mitchell, S.L.; Zhang, W. Selenoprotein p and apolipoprotein e receptor-2 interact at the blood-brain barrier and also within the brain to maintain an essential selenium pool that protects against neurodegeneration. FASEB J. 2014, 28, 3579–3588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forceville, X.; Mostert, V.; Pierantoni, A.; Vitoux, D.; Le Toumelin, P.; Plouvier, E.; Dehoux, M.; Thuillier, F.; Combes, A. Selenoprotein p, rather than glutathione peroxidase, as a potential marker of septic shock and related syndromes. Eur. Surg. Res. 2009, 43, 338–347. [Google Scholar] [CrossRef] [PubMed]
- Burk, R.F.; Hill, K.E. Selenoprotein p. A selenium-rich extracellular glycoprotein. J. Nutr. 1994, 124, 1891–1897. [Google Scholar] [CrossRef] [PubMed]
- Pico, C.; Serra, F.; Rodriguez, A.M.; Keijer, J.; Palou, A. Biomarkers of nutrition and health: New tools for new approaches. Nutrients 2019, 11, 1092. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age [years] | Se [µg/L] | SELENOP [mg/L] | GPx Activity [U/L] | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Weeks | 0 | 10 | 20 | 40 | 0 | 10 | 20 | 40 | 0 | 10 | 20 | 40 | ||
| - fish oil | 61 | 77.8 | 77.1 | 73.1 | 69.6 | 4.8 | 5.6 | 5.9 | 5.7 | 384.0 | 557.0 | 490.2 | 521.2 | |
| SD | 11.3 | 7.1 | 7.4 | 6.1 | 8.2 | 0.6 | 0.8 | 0.9 | 1.2 | 82.5 | 152.1 | 114.7 | 117.8 | |
| Range | 32–76 | 63.3–89.7 | 63.3–90.7 | 62.4–83.4 | 59.4–93.0 | 3.9–5.9 | 3.6–6.7 | 3.09–7.2 | 3.2–8.7 | 136.8–495.5 | 219.4–909.1 | 177.3–716.8 | 213.5–772.8 | |
| Weeks | 0 | 10 | 20 | 40 | 0 | 10 | 20 | 40 | 0 | 10 | 20 | 40 | ||
| + fish oil | 57 | 83.3 | 79.2 | 78.6 | 67.8 | 5.1 | 5.5 | 5.7 | 5.5 | 425.9 | 557.5 | 512.8 | 520.7 | |
| SD | 13.4 | 21.1 | 13.9 | 11.1 | 11.9 | 2.0 | 1.5 | 1.2 | 1.3 | 150.6 | 123.0 | 146.1 | 139.0 | |
| Range | 33–76 | 53.5–138.5 | 58.6–106.6 | 62.0–106.8 | 53.2–94.4 | 3.1–11.0 | 3.5–8.8 | 3.4–8.1 | 4.1–9.4 | 267.0–914.5 | 396.4–916.8 | 342.2–973.1 | 388.2–979.5 | |
| p-Value | 0.4750 | 0.9339 | 0.1131 | 0.3090 | 0.7575 | 0.3014 | 0.3618 | 0.3224 | 0.7206 | 0.9052 | 0.7722 | 0.4028 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Müller, S.M.; Dawczynski, C.; Wiest, J.; Lorkowski, S.; Kipp, A.P.; Schwerdtle, T. Functional Biomarkers for the Selenium Status in a Human Nutritional Intervention Study. Nutrients 2020, 12, 676. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12030676
Müller SM, Dawczynski C, Wiest J, Lorkowski S, Kipp AP, Schwerdtle T. Functional Biomarkers for the Selenium Status in a Human Nutritional Intervention Study. Nutrients. 2020; 12(3):676. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12030676
Chicago/Turabian StyleMüller, Sandra M., Christine Dawczynski, Johanna Wiest, Stefan Lorkowski, Anna P. Kipp, and Tanja Schwerdtle. 2020. "Functional Biomarkers for the Selenium Status in a Human Nutritional Intervention Study" Nutrients 12, no. 3: 676. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12030676