Effects of a Synbiotic Formula on Functional Bowel Disorders and Gut Microbiota Profile during Long-Term Home Enteral Nutrition (LTHEN): A Pilot Study

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Randomization
2.2. Ethical Aspects
2.3. Recruitment of Participants
2.4. Intervention
2.5. Clinical and Nutritional Assessment and Sample Collection
2.6. Gut Microbiota Analysis
2.7. MG Data Open Access Repository
2.8. Volatile Organic Compounds (VOCs)
3. Results
3.1. Nutritional Assessment and Biochemical Analyses
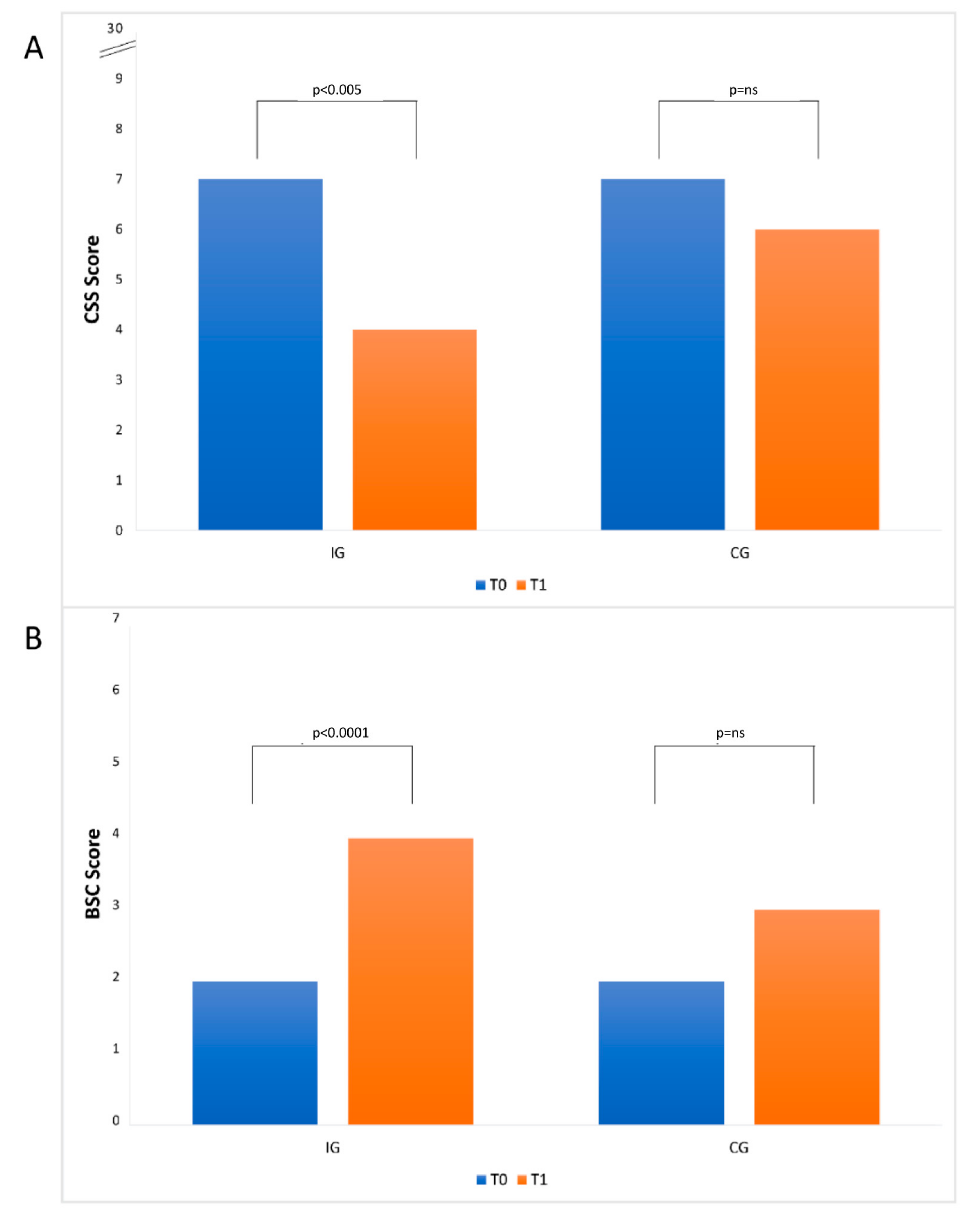
3.2. Constipation and Stool Consistency
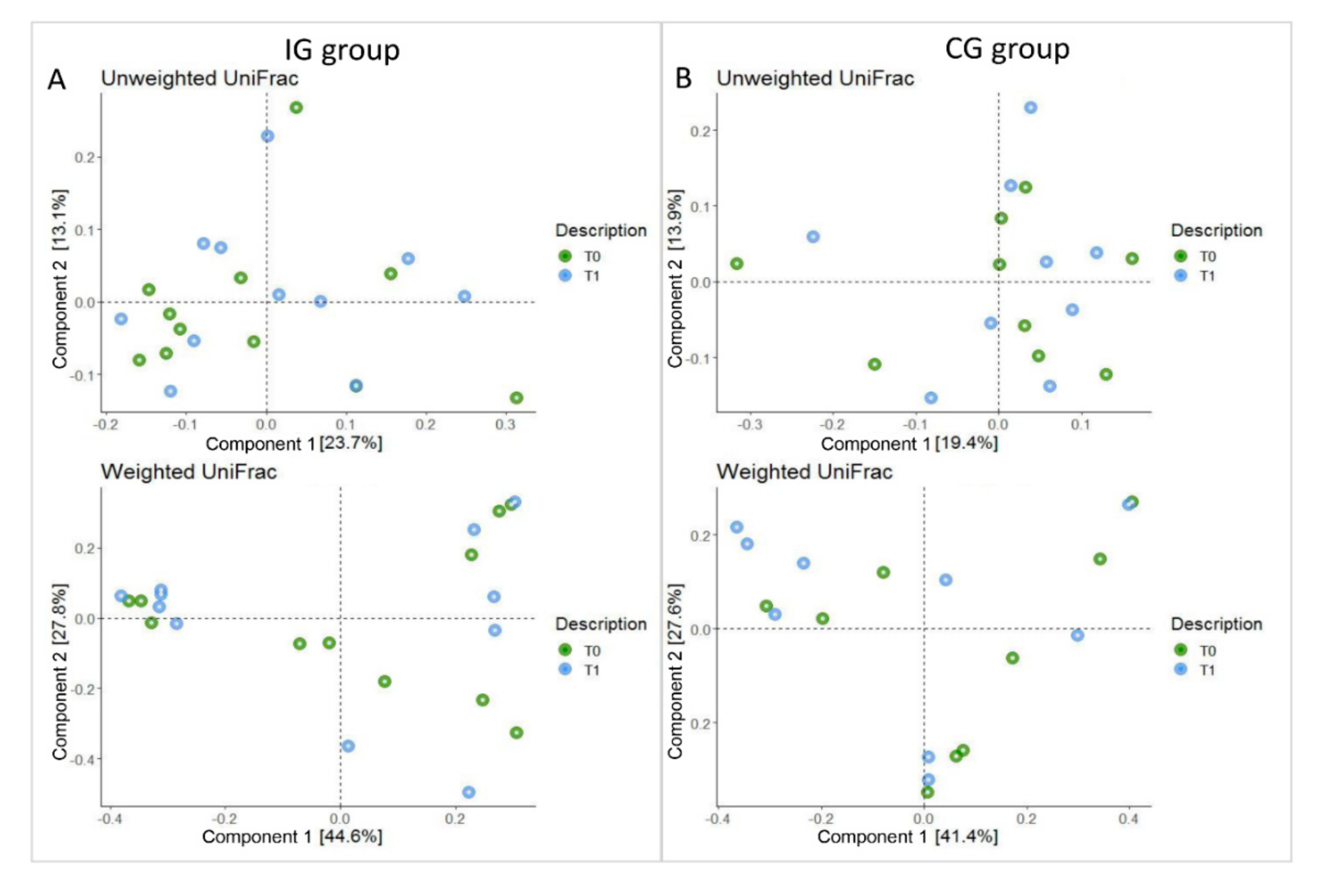
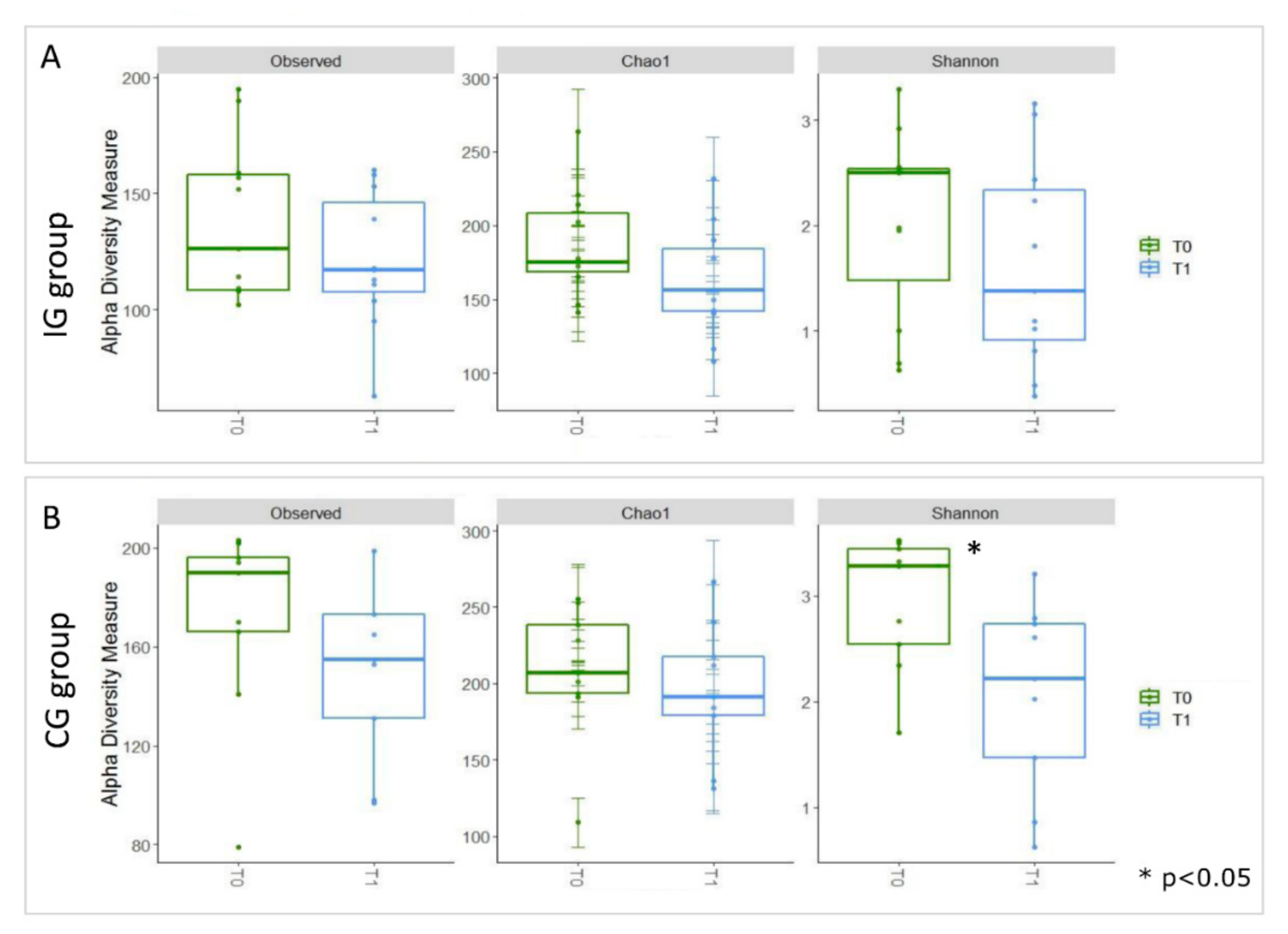
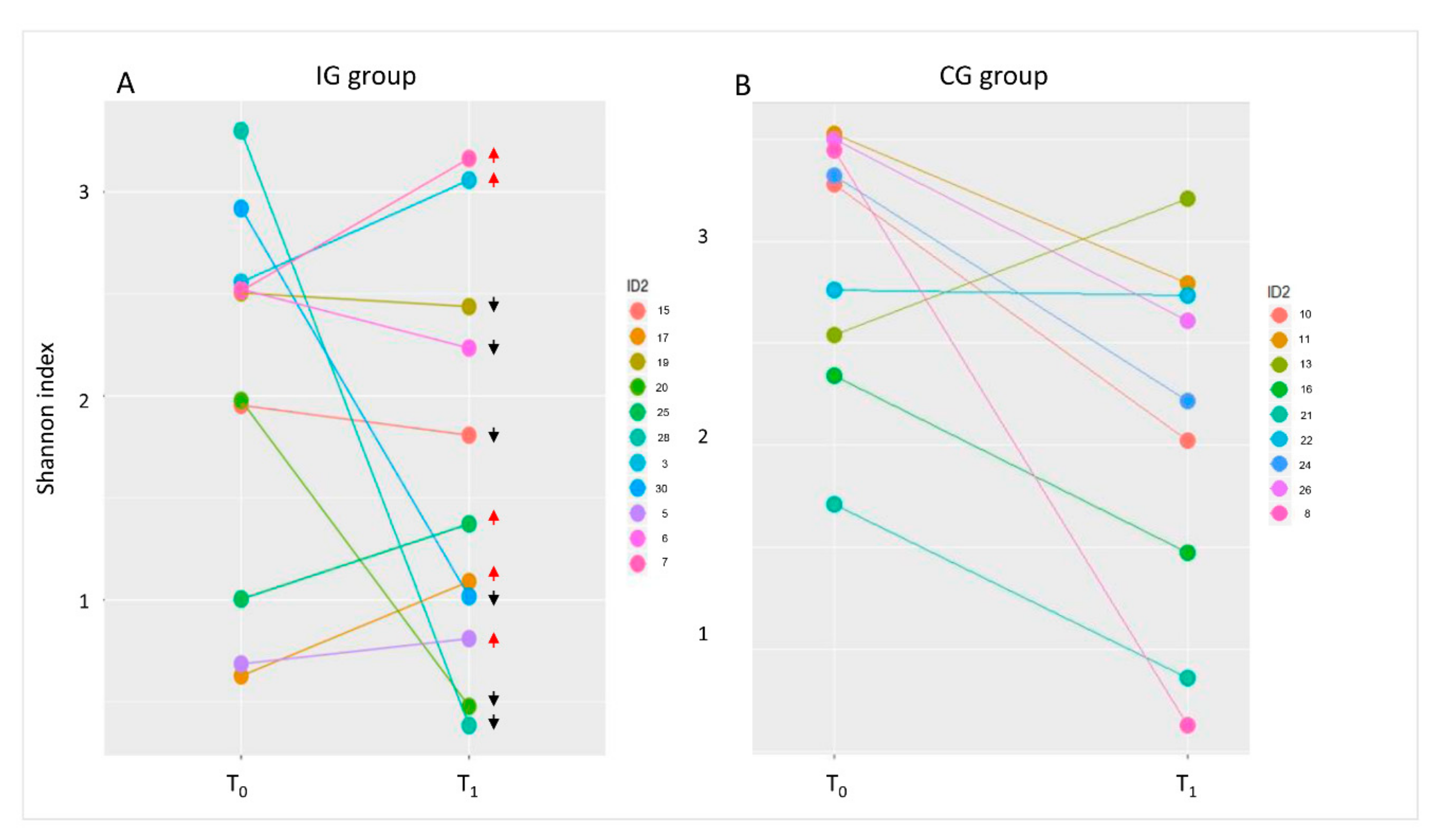
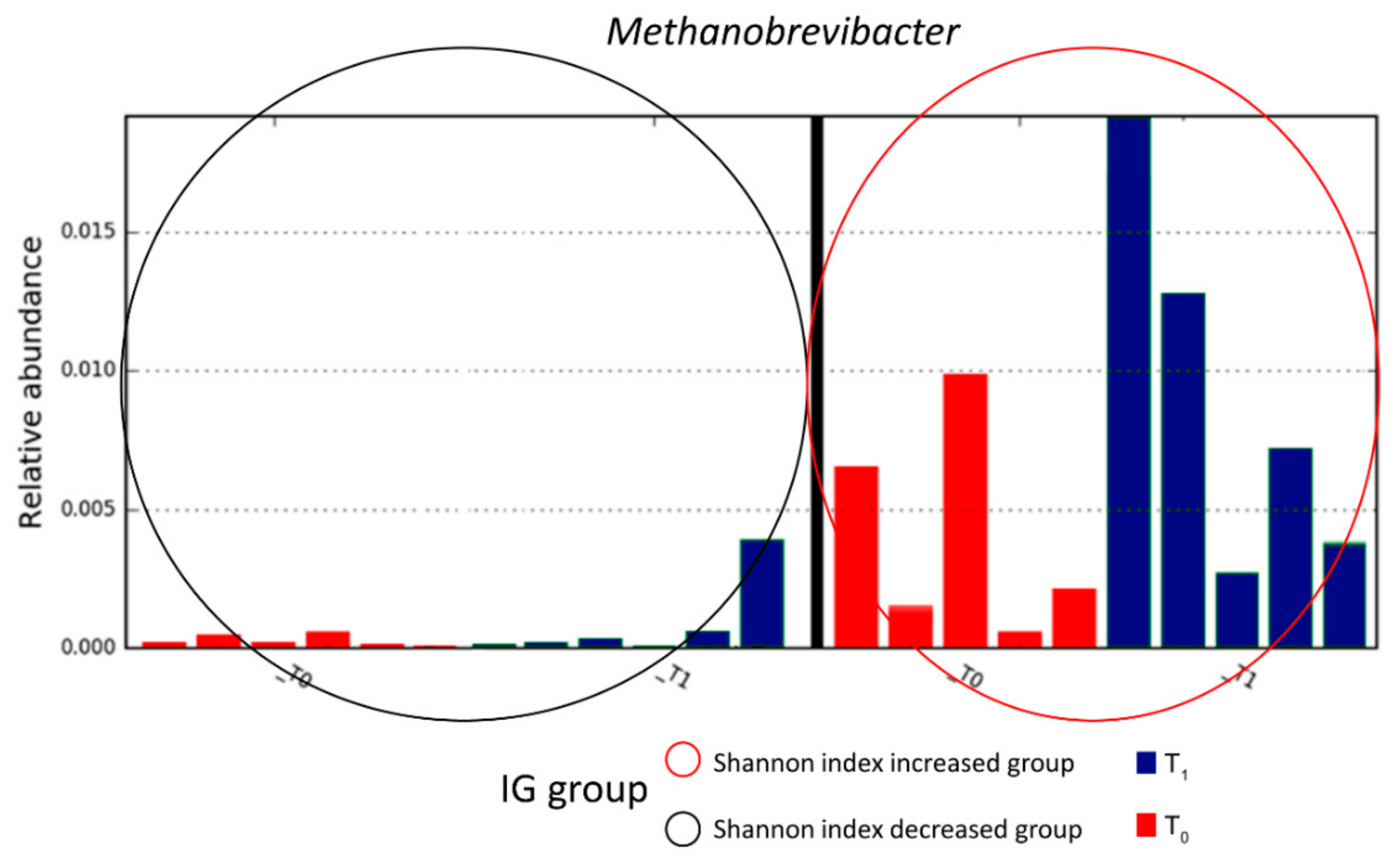
3.3. Composition of Intestinal Microbiota at Baseline (T0) and after Four Months of Intervention (T1)
3.4. Volatile Metabolome Profile at Baseline (T0) and after Four Months of Intervention (T1)
3.5. Pearson’s Correlation Test of Targeted Metagenomic and VOCs Data
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef] [PubMed]
- Zoetendal, E.G.; Rajilic-Stojanovic, M.; De Vos, W.M. High-throughput diversity and functionality analysis of the gastrointestinal tract microbiota. Gut 2008, 57, 1605–1615. [Google Scholar] [CrossRef] [PubMed]
- Del Chierico, F.; Abbatini, F.; Russo, A.; Quagliariello, A.; Reddel, S.; Capoccia, D.; Caccamo, R.; Corradini, S.G.; Nobili, V.; De Peppo, F.; et al. Gut Microbiota Markers in Obese Adolescent and Adult Patients: Age-Dependent Differential Patterns. Front. Microbiol. 2018, 9, 1210. [Google Scholar] [CrossRef] [PubMed]
- Presti, A.L.; Zorzi, F.; Del Chierico, F.; Altomare, A.; Cocca, S.; Avola, A.; De Biasio, F.; Russo, A.; Cella, E.; Reddel, S.; et al. Fecal and Mucosal Microbiota Profiling in Irritable Bowel Syndrome and Inflammatory Bowel Disease. Front. Microbiol. 2019, 10, 1655. [Google Scholar] [CrossRef] [Green Version]
- Quagliariello, A.; Del Chierico, F.; Russo, A.; Reddel, S.; Conte, G.; Lopetuso, L.R.; Ianiro, G.; Dallapiccola, B.; Cardona, F.; Gasbarrini, A.; et al. Gut Microbiota Profiling and Gut–Brain Crosstalk in Children Affected by Pediatric Acute-Onset Neuropsychiatric Syndrome and Pediatric Autoimmune Neuropsychiatric Disorders Associated With Streptococcal Infections. Front. Microbiol. 2018, 9, 675. [Google Scholar] [CrossRef]
- Ponziani, F.R.; Bhoori, S.; Castelli, C.; Putignani, L.; Rivoltini, L.; Del Chierico, F.; Sanguinetti, M.; Morelli, D.; Sterbini, F.P.; Petito, V.; et al. Hepatocellular Carcinoma Is Associated With Gut Microbiota Profile and Inflammation in Nonalcoholic Fatty Liver Disease. Hepatology 2019, 69, 107–120. [Google Scholar] [CrossRef]
- Del Chierico, F.; Nobili, V.; Vernocchi, P.; Russo, A.; De Stefanis, C.; Gnani, D.; Furlanello, C.; Zandonà, A.; Paci, P.; Capuani, G.; et al. Gut microbiota profiling of pediatric nonalcoholic fatty liver disease and obese patients unveiled by an integrated meta-omics-based approach. Hepatology 2017, 65, 451–464. [Google Scholar] [CrossRef]
- Vernocchi, P.; Del Chierico, F.; Russo, A.; Majo, F.; Rossitto, M.; Valerio, M.; Casadei, L.; La Storia, A.; De Filippis, F.; Rizzo, C.; et al. Gut microbiota signatures in cystic fibrosis: Loss of host CFTR function drives the microbiota enterophenotype. PLoS ONE 2018, 13, e0208171. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [Green Version]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Yang, M.; Zhou, K.; Zhang, L.; Tian, L.; Lv, S.; Jin, Y.; Qian, W.; Xiong, H.; Lin, R.; et al. Diversity of Duodenal and Rectal Microbiota in Biopsy Tissues and Luminal Contents in Healthy Volunteers. J. Microbiol. Biotechnol. 2015, 25, 1136–1145. [Google Scholar] [CrossRef] [PubMed]
- Otte, J.M.; Podolsky, D.K. Functional modulation of enterocytes by gram-positive and gram-negative microorganisms. Am. J. Physiol. Liver Physiol. 2004, 286, G613–G626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proctor, C.; Thiennimitr, P.; Chattipakorn, N.; Chattipakorn, S.C. Diet, gut microbiota and cognition. Metab. Brain Dis. 2016, 32, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Zmora, N.; Suez, J.; Elinav, E. You are what you eat: Diet, health and the gut microbiota. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 35–56. [Google Scholar] [CrossRef] [PubMed]
- Cederholm, T.; Barazzoni, R.; Austin, P.D.P.; Ballmer, P.P.; Biolo, G.; Bischoff, S.C.; Compher, C.; Correia, M.I.T.; Higashiguchi, T.T.; Holst, M.M.; et al. ESPEN guidelines on definitions and terminology of clinical nutrition. Clin. Nutr. 2017, 36, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Gatti, S.; Galeazzi, T.; Franceschini, E.; Annibali, R.; Albano, V.; Verma, A.K.; De Angelis, M.; Lionetti, E.; Catassi, C. Effects of the Exclusive Enteral Nutrition on the Microbiota Profile of Patients with Crohn’s Disease: A Systematic Review. Nutrients 2017, 9, 832. [Google Scholar] [CrossRef] [Green Version]
- Schneider, S. Microbiota and enteral nutrition. Gastroentérologie Clinique Biologique 2010, 34, S57–S61. [Google Scholar] [CrossRef]
- Cataldi-Betcher, E.L.; Seltzer, M.H.; Slocum, B.A.; Jones, K.W. Complications Occurring during Enteral Nutrition Support: A Prospective Study. J. Parenter. Enter. Nutr. 1983, 7, 546–552. [Google Scholar] [CrossRef]
- Schneider, S.; Le Gall, P.; Girard-Pipau, F.; Piche, T.; Pompei, A.; Nano, J.-L.; Hébuterne, X.; Rampal, P. Total artificial nutrition is associated with major changes in the fecal flora. Eur. J. Nutr. 2000, 39, 248–255. [Google Scholar] [CrossRef]
- Borowitz, S.M. Mobility and Motility: Constipation Impairs Enteral Feeding in Disabled and Immobile Patients. Pract. Gastroenterol. 2020, 10, 12–20. [Google Scholar]
- Tarleton, S.M.; Kraft, C.A.; Dibaise, J.K. Fiber-enriched enteral formulae: Advantageous or adding fuel to the fire? Pract. Gastroenterol. 2013, 37, 11–22. [Google Scholar]
- Turza, K.C.; Krenitsky, J.; Sawyer, R.G. Enteral feeding and vasoactive agents: Suggested guidelines for clinicians. Pract. Gastroenterol. 2009, 33, 11–12+15. [Google Scholar]
- Cresci, G.; Cúe, J. The Patient With Circulatory Shock: To Feed or Not to Feed? Nutr. Clin. Pr. 2008, 23, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Elia, M.; Engfer, M.B.; Green, C.J.; Silk, D.B.A. Systematic review and meta-analysis: The clinical and physiological effects of fibre-containing enteral formulae. Aliment. Pharmacol. Ther. 2007, 27, 120–145. [Google Scholar] [CrossRef] [PubMed]
- Kondo, J. Modulatory effects of Bifidobacterium longumBB536 on defecation in elderly patients receiving enteral feeding. World J. Gastroenterol. 2013, 19, 2162–2170. [Google Scholar] [CrossRef] [PubMed]
- Fosnes, G.S.; Lydersen, S.; Farup, P.G. Drugs and Constipation in Elderly in Nursing Homes: What Is the Relation? Gastroenterol. Res. Pr. 2012, 2012, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Pandey, K.R.; Naik, S.R.; Vakil, B.V. Probiotics, prebiotics and synbiotics- A review. J. Food Sci. Technol. 2015, 52, 7577–7587. [Google Scholar] [CrossRef]
- Agachan, F.; Chen, T.; Pfeifer, J.; Reissman, P.; Wexner, S.D. A constipation scoring system to simplify evaluation and management of constipated patients. Dis. Colon Rectum 1996, 39, 681–685. [Google Scholar] [CrossRef]
- Lewis, S.J.; Heaton, K.W. Stool Form Scale as a Useful Guide to Intestinal Transit Time. Scand. J. Gastroenterol. 1997, 32, 920–924. [Google Scholar] [CrossRef]
- Romani, L.; Del Chierico, F.; Chiriaco, M.; Foligno, S.; Reddel, S.; Salvatori, G.; Cifaldi, C.; Faraci, S.; Finocchi, A.; Rossi, P.; et al. Gut Mucosal and Fecal Microbiota Profiling Combined to Intestinal Immune System in Neonates Affected by Intestinal Ischemic Injuries. Front. Cell. Infect. Microbiol. 2020, 10, 59. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Bittinger, K.; Bushman, F.D.; DeSantis, T.Z.; Andersen, G.L.; Knight, R. PyNAST: A fexible tool for aligning sequences to a template alignment. Bioinformatics 2010, 26, 266–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeSantis, T.Z.; Hugenholtz, P.; Keller, K.; Brodie, E.L.; Larsen, N.; Piceno, Y.M.; Phan, R.; Andersen, G.L. NAST: A multiple sequence alignment server for comparative analysis of 16S rRNA genes. Nucleic Acids Res. 2006, 34, W394–W399. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botticelli, A.; Vernocchi, P.; Marini, F.; Quagliariello, A.; Cerbelli, B.; Reddel, S.; Del Chierico, F.; Di Pietro, F.; Giusti, R.; Tomassini, A.; et al. Gut metabolomics profiling of non-small cell lung cancer (NSCLC) patients under immunotherapy treatment. J. Transl. Med. 2020, 18, 49. [Google Scholar] [CrossRef] [PubMed]
- Garner, C.E.; Smith, S.; de Lacy Costello, B.; White, P.; Spencer, R.; Probert, C.S.J.; Ratcliffe, N.M. Volatile organic compounds from feces and their potential for diagnosis of gastrointestinal disease. FASEB J. 2007, 21, 1675–1688. [Google Scholar] [CrossRef] [Green Version]
- McClave, S.A.; Rd, B.E.T.; Martindale, R.G.; Rd, M.M.W.; Johnson, D.R.; Rd, C.B.; Rn, M.S.M.; Davanos, E.; Rice, T.W.; Rd, G.A.C.; et al. Guidelines for the Provision and Assessment of Nutrition Support Therapy in the Adult Critically Ill Patient. J. Parenter. Enter. Nutr. 2016, 40, 159–211. [Google Scholar] [CrossRef]
- Singer, P.; Blaser, A.R.; Berger, M.M.; Alhazzani, W.; Calder, P.C.; Casaer, M.P.; Hiesmayr, M.; Mayer, K.; Montejo, J.C.; Pichard, C.; et al. ESPEN guideline on clinical nutrition in the intensive care unit. Clin. Nutr. 2019, 38, 48–79. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.; Wang, Y.; Huang, Y.; Cui, Y.; Xiaoting, W.; Rao, Z.; Zhou, Y.; Wu, X. Effects of fiber and probiotics on diarrhea associated with enteral nutrition in gastric cancer patients. Medicine 2017, 96, e8418. [Google Scholar] [CrossRef] [PubMed]
- Whelan, K.; Judd, P.A.; Preedy, V.R.; Simmering, R.; Jann, A.; Taylor, M.A. Fructooligosaccharides and Fiber Partially Prevent the Alterations in Fecal Microbiota and Short-Chain Fatty Acid Concentrations Caused by Standard Enteral Formula in Healthy Humans. J. Nutr. 2005, 135, 1896–1902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chelakkot, C.; Ghim, J.; Ryu, S.H. Mechanisms regulating intestinal barrier integrity and its pathological implications. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Million, M.; Tomas, J.; Wagner, C.; Lelouard, H.; Raoult, D.; Gorvel, J.-P. New insights in gut microbiota and mucosal immunity of the small intestine. Hum. Microbiome J. 2018, 7, 23–32. [Google Scholar] [CrossRef]
- Markowiak-Kopeć, P.; Śliżewska, K. The Effect of Probiotics on the Production of Short-Chain Fatty Acids by Human Intestinal Microbiome. Nutrients 2020, 12, 1107. [Google Scholar] [CrossRef]
- Redanz, S.; Cheng, X.; Giacaman, R.A.; Pfeifer, C.S.; Merritt, J.; Kreth, J. Live and let die: Hydrogen peroxide production by the commensal flora and its role in maintaining a symbiotic microbiome. Mol. Oral Microbiol. 2018, 33, 337–352. [Google Scholar] [CrossRef]
- Zheng, J.; Gänzle, M.G.; Lin, X.B.; Ruan, L.; Sun, M. Diversity and dynamics of bacteriocins from human microbiome. Environ. Microbiol. 2014, 17, 2133–2143. [Google Scholar] [CrossRef]
- Der Vaart, I.B.V.; Heath, M.; Guagnini, F.; Kramer, M.F. In vitro evidence for efficacy in food intolerance for the multispecies probiotic formulation Ecologic® Tolerance (Syngut™). Benef. Microbes 2016, 7, 111–118. [Google Scholar] [CrossRef]
- Fassio, F.; Facioni, M.S.; Guagnini, F. Lactose Maldigestion, Malabsorption, and Intolerance: A Comprehensive Review with a Focus on Current Management and Future Perspectives. Nutrients 2018, 10, 1599. [Google Scholar] [CrossRef] [Green Version]
- Rossi, R.; Rossi, L.; Fassio, F. Clinical Follow-up of 96 Patients Affected by Irritable Bowel Syndrome Treated with a Novel Multi-strain Symbiotic. J. Contemp. Immunol. 2015. [Google Scholar] [CrossRef]
- Gibson, G.R.; Roberfroid, M.B. Dietary Modulation of the Human Colonic Microbiota: Introducing the Concept of Prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef] [PubMed]
- Gagliardi, A.; Totino, V.; Cacciotti, F.; Iebba, V.; Neroni, B.; Bonfiglio, G.; Trancassini, M.; Passariello, C.; Pantanella, F.; Schippa, S. Rebuilding the Gut Microbiota Ecosystem. Int. J. Environ. Res. Public Heal. 2018, 15, 1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vyas, U.; Ranganathan, N. Probiotics, Prebiotics, and Synbiotics: Gut and Beyond. Gastroenterol. Res. Pr. 2012, 2012, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, P.T.; Kashyap, P.C.; Preidis, G.A. Microbiota on biotics: Probiotics, prebiotics, and synbiotics to optimize growth and metabolism. Am. J. Physiol. Liver Physiol. 2020, 319, G382–G390. [Google Scholar] [CrossRef]
- Venegas-Borsellino, C.; Kwon, M. Impact of Soluble Fiber in the Microbiome and Outcomes in Critically Ill Patients. Curr. Nutr. Rep. 2019, 8, 347–355. [Google Scholar] [CrossRef]
- Johnson, T.W.; Gim, S.S.R.; Epp, L.; Mundi, M.S. Addressing Frequent Issues of Home Enteral Nutrition Patients. Nutr. Clin. Pr. 2019, 34, 186–195. [Google Scholar] [CrossRef] [Green Version]
- Tezcan, A.H.; Yagmurdur, H.; Leblebici, F. Enteral nutrition preference in critical care: Fibre-enriched or fi-bre-free? Asia Pac. J. Clin. Nutr. 2016, 25. [Google Scholar] [CrossRef]
- Soares, G.G.D.C.; Marinho, C.H.; Pitol, R.; Andretta, C.; Oliveira, E.; Martins, C.; Riella, M.C. Sporulated Bacillus as alternative treatment for diarrhea of hospitalized adult patients under enteral nutrition: A pilot randomized controlled study. Clin. Nutr. ESPEN 2017, 22, 13–18. [Google Scholar] [CrossRef]
- Tremaroli, V.; Bäckhed, F. Functional interactions between the gut microbiota and host metabolism. Nat. Cell Biol. 2012, 489, 242–249. [Google Scholar] [CrossRef]
- Samuel, B.S.; Hansen, E.E.; Manchester, J.K.; Coutinho, P.M.; Henrissat, B.; Fulton, R.; Latreille, P.; Kim, K.; Wilson, R.K.; Gordon, J.I. Genomic and metabolic adaptations of Methanobrevibacter smithii to the human gut. Proc. Natl. Acad. Sci. USA 2007, 104, 10643–10648. [Google Scholar] [CrossRef] [Green Version]
- Dridi, B.; Henry, M.; El Khéchine, A.; Raoult, D.; Drancourt, M. High Prevalence of Methanobrevibacter smithii and Methanosphaera stadtmanae Detected in the Human Gut Using an Improved DNA Detection Protocol. PLoS ONE 2009, 4, e7063. [Google Scholar] [CrossRef] [PubMed]
- Nowak, A.; Libudzisz, Z. Influence of phenol, p-cresol and indole on growth and survival of intestinal lactic acid bacteria. Anaerobe 2006, 12, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Rondanelli, M.; Perdoni, F.; Infantino, V.; Faliva, M.A.; Peroni, G.; Iannello, G.; Nichetti, M.; Alalwan, T.A.; Perna, S.; Cocuzza, C.E. Volatile Organic Compounds as Biomarkers of Gastrointestinal Diseases and Nutritional Status. J. Anal. Methods Chem. 2019, 2019, 7247802-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Blood Variables | IG T0 | IG T1 | p-Value | CG T0 | CG T1 | p-Value |
|---|---|---|---|---|---|---|
| Glycemia (g/dL) | 81 ± 30 | 79 ± 32 | 0.11 | 89 ± 22 | 87 ± 32 | 0.10 |
| Total Protein (g/dL) | 6.6 ± 0.50 | 6.3 ± 0.50 | 0.14 | 6.7 ± 0.50 | 6.6 ± 0.50 | 0.12 |
| Albumin (g/dL) | 3.7 ± 0.30 | 3.7 ± 0.30 | 0.17 | 3.7 ± 0.20 | 3.8 ± 0.30 | 0.11 |
| Transferrin (g/dL) | 205 ± 75 | 209 ± 39 | 0.17 | 211 ± 78 | 212 ± 42 | 0.16 |
| Cholesterol (mg/dL) | 153 ± 21 | 148 ± 34 | 0.12 | 160 ± 44 | 157 ± 50 | 0.16 |
| Triglycerides (mg/dL) | 75 ± 20 | 82 ± 40 | 0.11 | 86 ± 18 | 91 ± 22 | 0.12 |
| Haemoglobin (g/dL) | 13.2 ± 1.20 | 13.4 ± 1.30 | 0.13 | 13.40 ± 2.10 | 13 ± 2 | 0.14 |
| Vitamin B 12 (ng/L) | 567 ± 204 | 547 ± 198 | 0.10 | 588 ± 334 | 573 ± 292 | 0.11 |
| Folic Acid (ng/L) | 15.6 ± 4.20 | 15.3 ± 3.90 | 0.13 | 16.30 ± 3.20 | 17.3 ± 4.20 | 0.15 |
| Enteral Nutrition Intake | ||||||
| Protein (g) | 62 ± 16 | 61 ± 16 | 0.10 | 64 ± 17 | 63 ± 12 | 0.10 |
| Lipid (g) | 60 ± 14 | 58 ± 11 | 0.12 | 61 ± 5 | 60 ± 11 | 012 |
| Carbohydrates (g) | 180 ± 37 | 173 ± 21 | 0.11 | 185 ± 188 | 182 ± 28 | 0.11 |
| Energy (kcal) | 1508 ± 212 | 1457 ± 188 | 0.11 | 1545 ± 177 | 1520 ± 161 | 0.10 |
| Fibre (g) | 18 ± 4 | 17 ± 2 | 0.10 | 15 ± 4 | 16 ± 3 | 0.10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Onofrio, V.; Del Chierico, F.; Belci, P.; Vernocchi, P.; Quagliariello, A.; Reddel, S.; Conta, G.; Mancino, M.V.; Fadda, M.; Scigliano, M.C.; et al. Effects of a Synbiotic Formula on Functional Bowel Disorders and Gut Microbiota Profile during Long-Term Home Enteral Nutrition (LTHEN): A Pilot Study. Nutrients 2021, 13, 87. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13010087
D’Onofrio V, Del Chierico F, Belci P, Vernocchi P, Quagliariello A, Reddel S, Conta G, Mancino MV, Fadda M, Scigliano MC, et al. Effects of a Synbiotic Formula on Functional Bowel Disorders and Gut Microbiota Profile during Long-Term Home Enteral Nutrition (LTHEN): A Pilot Study. Nutrients. 2021; 13(1):87. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13010087
Chicago/Turabian StyleD’Onofrio, Valentina, Federica Del Chierico, Paola Belci, Pamela Vernocchi, Andrea Quagliariello, Sofia Reddel, Giorgia Conta, Maria Vittoria Mancino, Maurizio Fadda, Maria Carmine Scigliano, and et al. 2021. "Effects of a Synbiotic Formula on Functional Bowel Disorders and Gut Microbiota Profile during Long-Term Home Enteral Nutrition (LTHEN): A Pilot Study" Nutrients 13, no. 1: 87. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13010087