
Effects of Probiotic Mixture Supplementation on the Immune Response to the 13-Valent Pneumococcal Conjugate Vaccine in People Living with HIV

 , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
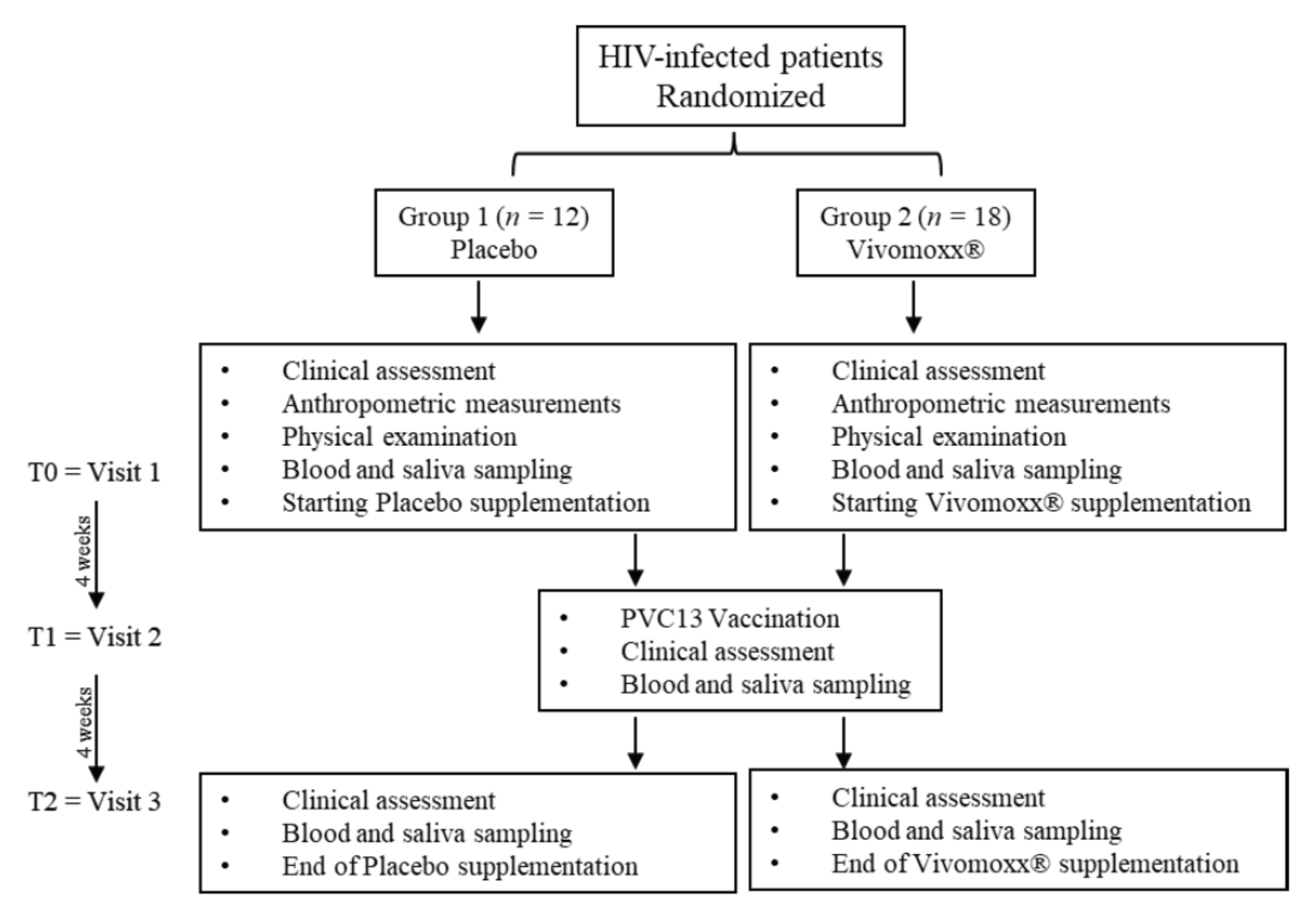
2.1. Patient Recruitment and Eligibility
2.2. Immunization Protocol and Sample Collection
2.3. Biochemical Analyses
2.4. Virologic and Immunologic Markers
2.5. Human Anti-Pneumococcal CPS13 IgG Measurements
2.6. Salivary IgA Measurements
2.7. Cytokines Measurements
2.8. Statistical Analysis
3. Results
3.1. Study Population
3.2. Metabolic and Inflammatory Markers
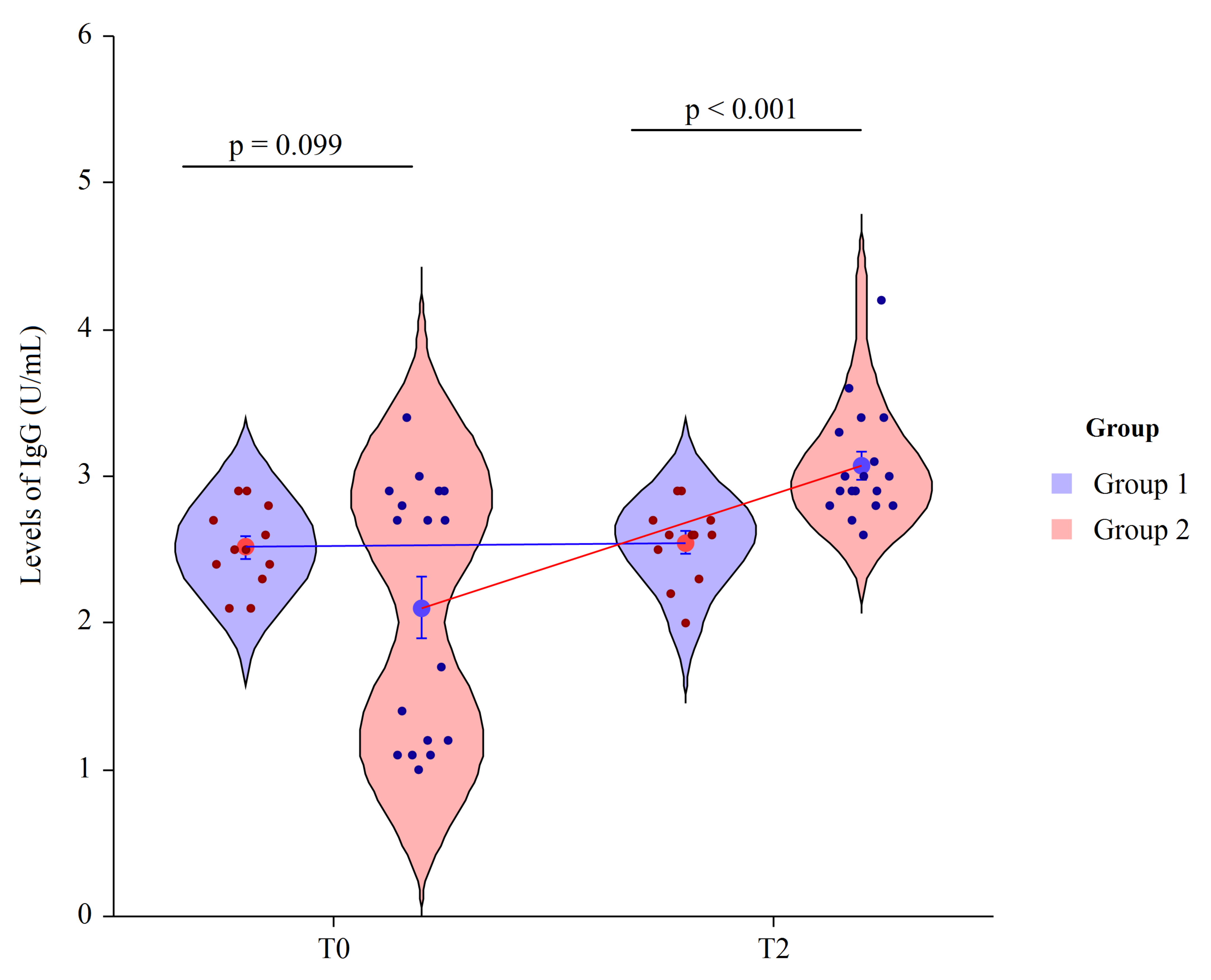
3.3. Human Anti-Pneumococcal CPS13
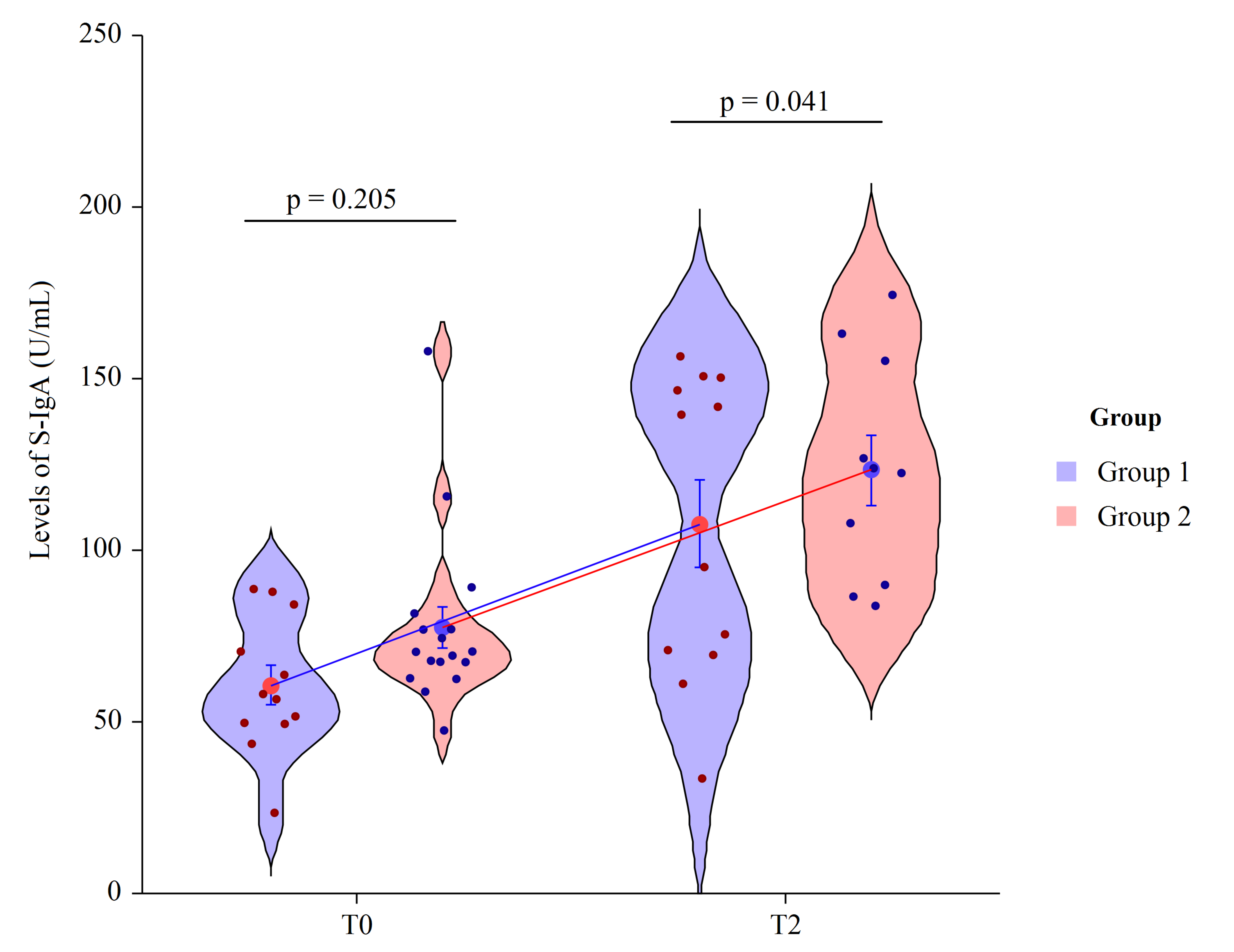
3.4. Salivary IgA
3.5. Measures of B Cell Phenotypes
3.6. Measures of Peripheral Blood T Cell Subpopulations
3.7. Serum Cytokine’s Levels
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mazzotta, E.; Riccardi, N.; Tontodonati, M.; Gabrielli, C.; Mazzocato, S.; Mazzetti, M.; Falasca, K.; Vecchiet, J.; Barchiesi, F.; Francisi, D.; et al. Prevalence and predictors of malignancies in HIV patients: Results of a retrospective multicentric Italian cohort. Infez. Med. 2019, 27, 53–57. [Google Scholar] [PubMed]
- Titanji, K.; De Milito, A.; Cagigi, A.; Thorstensson, R.; Grützmeier, S.; Atlas, A.; Hejdeman, B.; Kroon, F.P.; Lopalco, L.; Nilsson, A.; et al. Loss of memory B cells impairs maintenance of long-term serologic memory during HIV-1 infection. Blood 2006, 108, 1580–1587. [Google Scholar] [CrossRef] [Green Version]
- Yin, Z.; Rice, B.D.; Waight, P.; Miller, E.; George, R.; Brown, A.E.; Smith, R.D.; Slack, M.; Delpech, V.C. Invasive pneumococcal disease among HIV-positive individuals, 2000–2009. AIDS 2012, 26, 87–94. [Google Scholar] [CrossRef]
- Weiser, J.N.; Ferreira, D.M.; Paton, J.C. Streptococcus pneumoniae: Transmission, colonization and invasion. Nat. Rev. Microbiol. 2018, 16, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Kohli, R.; Lo, Y.; Homel, P.; Flanigan, T.P.; Gardner, L.I.; Howard, A.A.; Rompalo, A.M.; Moskaleva, G.; Schuman, P.; Schoenbaum, E.E.; et al. Bacterial pneumonia, HIV therapy, and disease progression among HIV-infected women in the HIV epidemiologic research (HER) study. Clin. Infect. Dis. 2006, 43, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Konkle-Parker, D. Vaccination of immunocompromised individuals: IDSA clinical practice guidelines. HIV Clin. 2014, 26, 1–3. [Google Scholar] [PubMed]
- Papadatou, I.; Spoulou, V. Pneumococcal Vaccination in High-Risk Individuals: Are We Doing It Right? Clin. Vaccine Immunol. 2016, 23, 388–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- French, N.; Gordon, S.B.; Mwalukomo, T.; White, S.A.; Mwafulirwa, G.; Longwe, H.; Mwaiponya, M.; Zijlstra, E.E.; Molyneux, M.E.; Gilks, C.F. A trial of a 7-valent pneumococcal conjugate vaccine in HIV-infected adults. N. Engl. J. Med. 2010, 362, 812–822. [Google Scholar] [CrossRef] [Green Version]
- Klugman, K.P.; Madhi, S.A.; Huebner, R.E.; Kohberger, R.; Mbelle, N.; Pierce, N.; Vaccine Trialists Group. A trial of a 9-valent pneumococcal conjugate vaccine in children with and those without HIV infection. N. Engl. J. Med. 2003, 349, 1341–1348. [Google Scholar] [CrossRef]
- Bhorat, A.E.; Madhi, S.A.; Laudat, F.; Sundaraiyer, V.; Gurtman, A.; Jansen, K.U.; Scott, D.A.; Emini, E.A.; Gruber, W.C.; Schmoele-Thoma, B. Immunogenicity and safety of the 13-valent pneumococcal conjugate vaccine in HIV-infected individuals naive to pneumococcal vaccination. AIDS 2015, 29, 1345–1354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynn, M.A.; Tumes, D.J.; Choo, J.M.; Sribnaia, A.; Blake, S.J.; Leong, L.E.X.; Young, G.P.; Marshall, H.S.; Wesselingh, S.L.; Rogers, G.B.; et al. Early-Life Antibiotic-Driven Dysbiosis Leads to Dysregulated Vaccine Immune Responses in Mice. Cell Host Microbe 2018, 23, 653–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arunachalam, K.; Gill, H.S.; Chandra, R.K. Enhancement of natural immune function by dietary consumption of Bifidobacterium lactis (HN019). Eur. J. Clin. Nutr. 2000, 54, 263–267. [Google Scholar] [CrossRef] [Green Version]
- Gill, H.S.; Rutherfurd, K.J.; Prasad, J.; Gopal, P.K. Enhancement of natural and acquired immunity by Lactobacillus rhamnosus (HN001), Lactobacillus acidophilus (HN017) and Bifidobacterium lactis (HN019). Br. J. Nutr. 2000, 83, 167–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, H.S.; Rutherfurd, K.J.; Cross, M.L.; Gopal, P.K. Enhancement of immunity in the elderly by dietary supplementation with the probiotic Bifidobacterium lactis HN019. Am. J. Clin. Nutr. 2001, 74, 833–839. [Google Scholar] [CrossRef]
- Mohr, E.; Siegrist, C.A. Vaccination in early life: Standing up to the challenges. Curr. Opin. Immunol. 2016, 41, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Artz, A.S.; Ershler, W.B.; Longo, D.L. Pneumococcal vaccination and revaccination of older adults. Clin. Microbiol. Rev. 2003, 16, 308–318. [Google Scholar] [CrossRef] [Green Version]
- Crakes, K.R.; Jiang, G. Gut Microbiome Alterations during HIV/SIV Infection: Implications for HIV Cure. Front Microbiol. 2019, 10, 1104. [Google Scholar] [CrossRef] [PubMed]
- Cristofori, F.; Dargenio, V.N.; Dargenio, C.; Miniello, V.L.; Barone, M.; Francavilla, R. Anti-Inflammatory and Immunomodulatory Effects of Probiotics in Gut Inflammation: A Door to the Body. Front Immunol. 2021, 12, 578386. [Google Scholar] [CrossRef]
- D’Angelo, C.; Reale, M.; Costantini, E. Microbiota and Probiotics in Health and HIV Infection. Nutrients 2017, 9, 615. [Google Scholar] [CrossRef] [Green Version]
- Ho, Y.L.; Brandão, A.P.; de Cunto Brandileone, M.C.; Lopes, M.H. Immunogenicity and safety of pneumococcal conjugate polysaccharide and free polysaccharide vaccines alone or combined in HIV-infected adults in Brazil. Vaccine 2013, 31, 4047–4053. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.Y.; Tsai, M.S.; Kuo, K.C.; Tsai, J.C.; Sun, H.Y.; Cheng, A.C.; Chang, S.Y.; Lee, C.H.; Hung, C.C. Pneumococcal vaccination among HIV-infected adult patients in the era of combination antiretroviral therapy. Hum. Vaccines Immunother. 2014, 10, 3700–3710. [Google Scholar] [CrossRef] [Green Version]
- Avci, F.Y.; Li, X.; Tsuji, M.; Kasper, D.L. A mechanism for glycoconjugate vaccine activation of the adaptive immune system and its implications for vaccine design. Nat. Med. 2011, 17, 1602–1609. [Google Scholar] [CrossRef] [Green Version]
- Shiramoto, M.; Hanada, R.; Juergens, C.; Shoji, Y.; Yoshida, M.; Ballan, B.; Cooper, D.; Gruber, W.C.; Scott, D.A.; Schmoele-Thoma, B. Immunogenicity and safety of the 13-valent pneumococcal conjugate vaccine compared to the 23-valent pneumococcal polysaccharide vaccine in elderly Japanese adults. Hum. Vaccines Immunother. 2015, 11, 2198–2206. [Google Scholar] [CrossRef]
- Keely, S.; Talley, N.J.; Hansbro, P.M. Pulmonary-intestinal crosstalk in mucosal inflammatory disease. Mucosal Immunol. 2012, 5, 7–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pabst, O.; Hornef, M. Gut microbiota: A natural adjuvant for vaccination. Immunity 2014, 41, 349–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falasca, K.; Vecchiet, J.; Ucciferri, C.; Di Nicola, M.; D’Angelo, C.; Reale, M. Effect of Probiotic Supplement on Cytokine Levels in HIV-Infected Individuals: A Preliminary Study. Nutrients 2015, 7, 8335–8347. [Google Scholar] [CrossRef]
- Mitra, S.; Stein, G.E.; Bhupalam, S.; Havlichek, D.H. Immunogenicity of 13-Valent Conjugate Pneumococcal Vaccine in Patients 50 Years and Older with End-Stage Renal Disease and on Dialysis. Clin. Vaccine Immunol. 2016, 23, 884–887. [Google Scholar] [CrossRef] [Green Version]
- Vandecasteele, S.J.; De Bacquer, D.; Caluwe, R.; Ombelet, S.; Van Vlem, B. Immunogenicity and safety of the 13-valent Pneumococcal Conjugate vaccine in 23-valent pneumococcal polysaccharide vaccine-naive and pre-immunized patients under treatment with chronic haemodialysis: A longitudinal quasi-experimental phase IV study. Clin. Microbiol. Infect. 2018, 24, 65–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Pol, W.; Vidarsson, G.; Vilé, H.A.; van de Winkel, J.G.; Rodriguez, M.E. Pneumococcal capsular polysaccharide-specific IgA triggers efficient neutrophil effector functions via FcalphaRI (CD89). J. Infect. Dis. 2000, 182, 1139–1145. [Google Scholar] [CrossRef]
- Fukuyama, Y.; King, J.D.; Kataoka, K.; Kobayashi, R.; Gilbert, R.S.; Oishi, K.; Hollingshead, S.K.; Briles, D.E.; Fujihashi, K. Secretory-IgA antibodies play an important role in the immunity to Streptococcus pneumoniae. J. Immunol. 2010, 185, 1755–1762. [Google Scholar] [CrossRef] [Green Version]
- Aghamohammadi, A.; Cheraghi, T.; Gharagozlou, M.; Movahedi, M.; Rezaei, N.; Yeganeh, M.; Parvaneh, N.; Abolhassani, H.; Pourpak, Z.; Moin, M. IgA deficiency: Correlation between clinical and immunological phenotypes. J. Clin. Immunol. 2009, 29, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Lue, C.; Tarkowski, A.; Mestecky, J. Systemic immunization with pneumococcal polysaccharide vaccine induces a predominant IgA2 response of peripheral blood lymphocytes and increases of both serum and secretory anti-pneumococcal antibodies. J. Immunol. 1988, 140, 3793–3800. [Google Scholar] [PubMed]
- Orami, T.; Ford, R.; Kirkham, L.A.; Thornton, R.; Corscadden, K.; Richmond, P.C.; Pomat, W.S.; van den Biggelaar, A.H.J.; Lehmann, D. Neonatal Pneumococcal Conjugate Vaccine Trial team. Pneumococcal conjugate vaccine primes mucosal immune responses to pneumococcal polysaccharide vaccine booster in Papua New Guinean children. Vaccine 2020, 38, 7977–7988. [Google Scholar] [CrossRef] [PubMed]
- Corthésy, B.; Gaskins, H.R.; Mercenier, A. Cross-talk between probiotic bacteria and the host immune system. J. Nutr. 2007, 137, 781S–790S. [Google Scholar] [CrossRef] [Green Version]
- Maidens, C.; Childs, C.; Przemska, A.; Dayel, I.B.; Yaqoob, P. Modulation of vaccine response by concomitant probiotic administration. Br. J. Clin. Pharmacol. 2013, 75, 663–670. [Google Scholar] [CrossRef] [Green Version]
- Thitilertdecha, P.; Khowawisetsut, L.; Ammaranond, P.; Poungpairoj, P.; Tantithavorn, V.; Onlamoon, N. Impact of Vaccination on Distribution of T Cell Subsets in Antiretroviral-Treated HIV-Infected Children. Dis. Markers 2017, 2017, 5729639. [Google Scholar] [CrossRef] [Green Version]
- Dell’Acqua, R.; Galli, L.; Poli, A.; Mastrangelo, A.; Guffanti, M.; Tadini, P.; Zandona, D.; Danise, A.; Gianotti, N.; Lazzarin, A.; et al. Viro-immunological outcomes after 13-valent pneumococcal vaccination in HIV-1-infected individuals on stable virological suppression. AIDS 2019, 33, 1987–1994. [Google Scholar] [CrossRef]
- Rossheim, A.E.; Young, A.M.; Siik, J.; Cunningham, T.D.; Troy, S.B. Association of time since pneumococcal polysaccharide vaccine receipt and CD4 count with antibody response to the 13-valent pneumococcal conjugate vaccine in HIV-infected adults. Hum. Vaccines Immunother. 2016, 12, 2117–2123. [Google Scholar] [CrossRef] [Green Version]
- Rask, C.; Adlerberth, I.; Berggren, A.; Ahrén, I.L.; Wold, A.E. Differential effect on cell-mediated immunity in human volunteers after intake of different lactobacilli. Clin. Exp. Immunol. 2013, 172, 321–332. [Google Scholar] [CrossRef]
- Van der Poll, T.; Marchant, A.; Keogh, C.V.; Goldman, M.; Lowry, S.F. Interleukin-10 impairs host defense in murine pneumococcal pneumonia. J. Infect. Dis. 1996, 174, 994–1000. [Google Scholar] [CrossRef] [Green Version]
- González, L.A.; Melo-González, F.; Sebastián, V.P.; Vallejos, O.P.; Noguera, L.P.; Suazo, I.D.; Schultz, B.M.; Manosalva, A.H.; Peñaloza, H.F.; Soto, J.A.; et al. Characterization of the Anti-Inflammatory Capacity of IL-10-Producing Neutrophils in Response to Streptococcus pneumonia Infection. Front Immunol. 2021, 12, 638917. [Google Scholar] [CrossRef] [PubMed]
- Madsen, M.; Lebenthal, Y.; Cheng, Q.; Smith, B.L.; Hostetter, M.K. A pneumococcal protein that elicits interleukin-8 from pulmonary epithelial cells. J. Infect. Dis. 2000, 181, 1330–1336. [Google Scholar] [CrossRef] [PubMed]
- Fish, E.N. The X-files in immunity: Sex-based differences predispose immune responses. Nat. Rev. Immunol. 2008, 8, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. HIV Surveillance Report, 2018. (Updated); 2020. Available online: http://www.cdc.gov/hiv/library/reports/hiv-surveillance.html (accessed on 7 December 2021).




{kind=link}
{kind=link}
{kind=link}
| Group 1 | Group 2 | p-Value | |
|---|---|---|---|
| n = 12 | n = 18 | ||
| Age (years) mean ± SD | 48.7± 9.1 | 47.5 ± 9.3 | 0.735 |
| BMI (kg/m2) | 27.0 ± 3.8 | 24.7 ± 2.0 | 0.117 |
| Risk factor, n (%) | |||
| eterosex | 6 (50.0) | 11 (61.1) | 0.656 |
| omosex | 6 (50.0) | 7 (38.9) |
| Group 1 | Group 2 | p-Value | |
|---|---|---|---|
| hs-CRP (mg/100 mL) T0 | 2.4 ± 0.3 | 2.6 ± 2.8 | 0.827 |
| T2 | 0.9 ± 0.7 | 7.0 ± 19.7 | 0.204 |
| Gly (mmol/L) T0 | 86.5 ± 25.4 | 92.1 ± 41.9 | 0.652 |
| T2 | 97.2 ± 33.5 | 93.8 ± 30.1 | 0.784 |
| TG (mg/dL) T0 | 153.0 ± 108.0 | 131.0 ± 90.8 | 0.580 |
| T2 | 120.0 ± 59.9 | 126.0 ± 78.9 | 0.829 |
| Chol (mg/dL) T0 | 198.0 ± 53.2 | 209.0 ± 38.9 | 0.561 |
| T2 | 200.0 ± 37.5 | 200.0 ± 31.1 | 0.970 |
| HDL (mg/dL) T0 | 49.2 ± 14.4 | 54.1 ± 10.6 | 0.335 |
| T2 | 50.8 ± 15.5 | 51.1 ± 11.3 | 0.954 |
| LDL (mg/dL) T0 | 118.0 ± 37.1 | 138.0 ± 32.4 | 0.145 |
| T2 | 111.0 ± 31.9 | 129.0 ± 27.1 | 0.129 |
| AST (IU/L−1) T0 | 34.8 ± 34.5 | 20.7 ± 7.8 | 0.190 |
| T2 | 30.8 ± 25.8 | 20.4 ± 7.9 | 0.203 |
| ALT (IU/L−1) T0 | 32.0 ± 15.3 | 24.8 ± 18.3 | 0.255 |
| T2 | 30.9 ± 10.0 | 21.5 ± 14.2 | 0.042 |
| GGT (IU/L−1) T0 | 32.5 ± 20.7 | 26.7 ± 25.6 | 0.503 |
| T2 | 34.3 ± 24.0 | 23.3 ± 16.6 | 0.182 |
| CRE (mL/min) T0 | 1.3 ± 1.2 | 1.0 ± 0.2 | 0.392 |
| T2 | 1.5 ± 1.7 | 1.0 ± 0.2 | 0.326 |
| T0 | T1 | T2 | ||
|---|---|---|---|---|
| B lymphocytes (%) mean ± SD | Group 1 | 10.6 ± 4.7 | 11.1 ± 9.5 | 12.7 ± 9.0 |
| Group 2 | 10.4 ± 4.9 | 15.5 ± 8.6 | 19.2 ± 9.5 | |
| p-value | 0.911 | 0.180 | 0.071 | |
| B lymphocytes (cells/μL) mean ± SD | Group 1 | 248.0 ± 91.5 | 260.0 ± 97.5 | 329.0 ± 165.4 |
| Group 2 | 214.0 ± 59.5 | 281.0 ± 104.5 | 307.0 ± 65.5 | |
| p-value | 0.271 | 0.580 | 0.668 |
| T0 | T1 | T2 | ||
|---|---|---|---|---|
| CD4+ (%) mean ± SD | Group 1 | 23.4 ± 9.1 | 24.5 ± 9.6 | 23.9 ± 8.5 |
| Group 2 | 28.9 ± 3.3 | 36.9 ± 9.3 * | 33.6 ± 7.5 # | |
| p-value | 0.066 | 0.002 | 0.009 | |
| CD8+ (%) mean ± SD | Group 1 | 45.8 ± 11.0 | 45.4 ± 10.6 | 46.0 ± 9.4 |
| Group 2 | 39.3 ± 10.8 | 44.9 ± 5.38 | 43.9 ± 5.0 | |
| p-value | 0.123 | 0.885 | 0.499 | |
| CD4+ (cells/μL) mean ± SD | Group 1 | 600.0 ± 149.5 | 612.0 ± 297.8 | 591.0 ± 178.7 |
| Group 2 | 691.0 ± 134.5 | 820.0 ± 263.9 | 710.0 ± 154.9 | |
| p-value | 0.103 | 0.063 | 0.063 | |
| CD8+ (cells/μL) mean ± SD | Group 1 | 1073.0 ± 423.0 | 1093.0 ± 445.0 | 1097.0 ± 407.0 |
| Group 2 | 794.0 ± 266.0 | 916.0 ± 317.0 | 872.0 ± 308.0 | |
| p-value | 0.081 | 0.079 | 0.119 |
| IL-8 | T0 | T1 | T2 |
|---|---|---|---|
| Group 1 | 47.3 ± 18.3 | 46.7 ± 15.3 | 42.4 ± 14.1 |
| Group 2 | 54.4 ± 25.2 | 117.0 ± 20.0 * | 152.0 ± 15.6 *# |
| p-value | 0.380 | <0.001 | <0.001 |
| IL-10 | T0 | T1 | T2 |
|---|---|---|---|
| Group 1 | 14.0 ± 25.7 | 15.3 ± 5.1 | 18.6 ± 14.0 |
| Group 2 | 24.1 ± 28.8 | 96.2 ± 41.1 * | 143.0 ± 59.3 *# |
| p-value | 0.194 | <0.001 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reale, M.; Ucciferri, C.; Costantini, E.; Di Nicola, M.; Porreca, A.; Di Giovanni, P.; Pontolillo, M.; Auricchio, A.; Vecchiet, J.; Falasca, K. Effects of Probiotic Mixture Supplementation on the Immune Response to the 13-Valent Pneumococcal Conjugate Vaccine in People Living with HIV. Nutrients 2021, 13, 4412. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13124412
Reale M, Ucciferri C, Costantini E, Di Nicola M, Porreca A, Di Giovanni P, Pontolillo M, Auricchio A, Vecchiet J, Falasca K. Effects of Probiotic Mixture Supplementation on the Immune Response to the 13-Valent Pneumococcal Conjugate Vaccine in People Living with HIV. Nutrients. 2021; 13(12):4412. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13124412
Chicago/Turabian StyleReale, Marcella, Claudio Ucciferri, Erica Costantini, Marta Di Nicola, Annamaria Porreca, Pamela Di Giovanni, Michela Pontolillo, Antonio Auricchio, Jacopo Vecchiet, and Katia Falasca. 2021. "Effects of Probiotic Mixture Supplementation on the Immune Response to the 13-Valent Pneumococcal Conjugate Vaccine in People Living with HIV" Nutrients 13, no. 12: 4412. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13124412