
Daily Rhythm of Fractal Cardiac Dynamics Links to Weight Loss Resistance: Interaction with CLOCK 3111T/C Genetic Variant

 , , ,
, , ,
Abstract
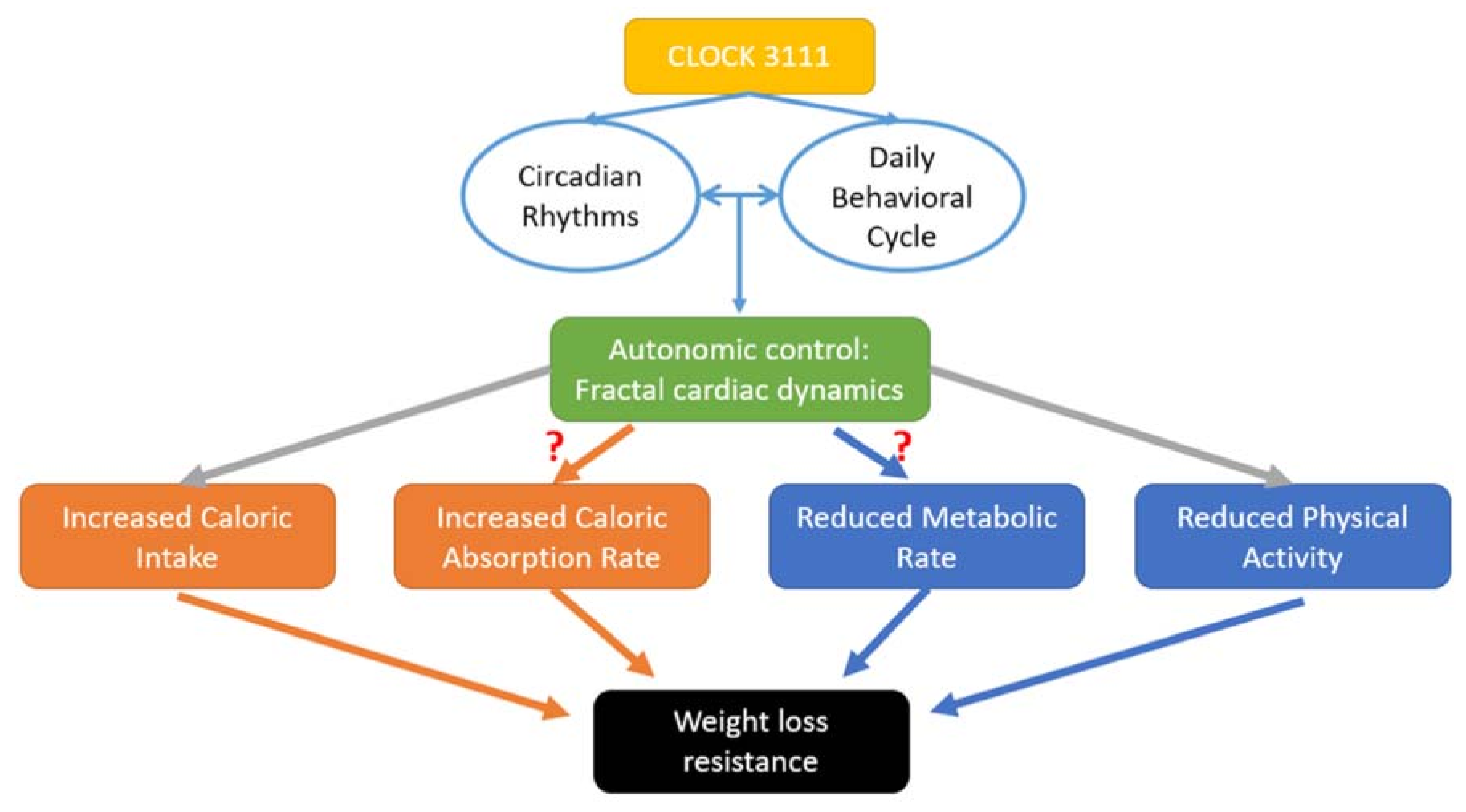
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Weight Loss Treatment
2.3. Weight Loss Evolution
2.4. Physical Activity and Energy Intake Assessment
2.5. DNA Isolation and CLOCK Genotyping
2.6. Ambulatory Electrogradiography (ECG) and Heartbeat Recordings
2.7. Fractal Analysis of Heartbeat Fluctuations
2.8. Rhythmicity Assessment
2.9. Statistical Analysis
3. Results
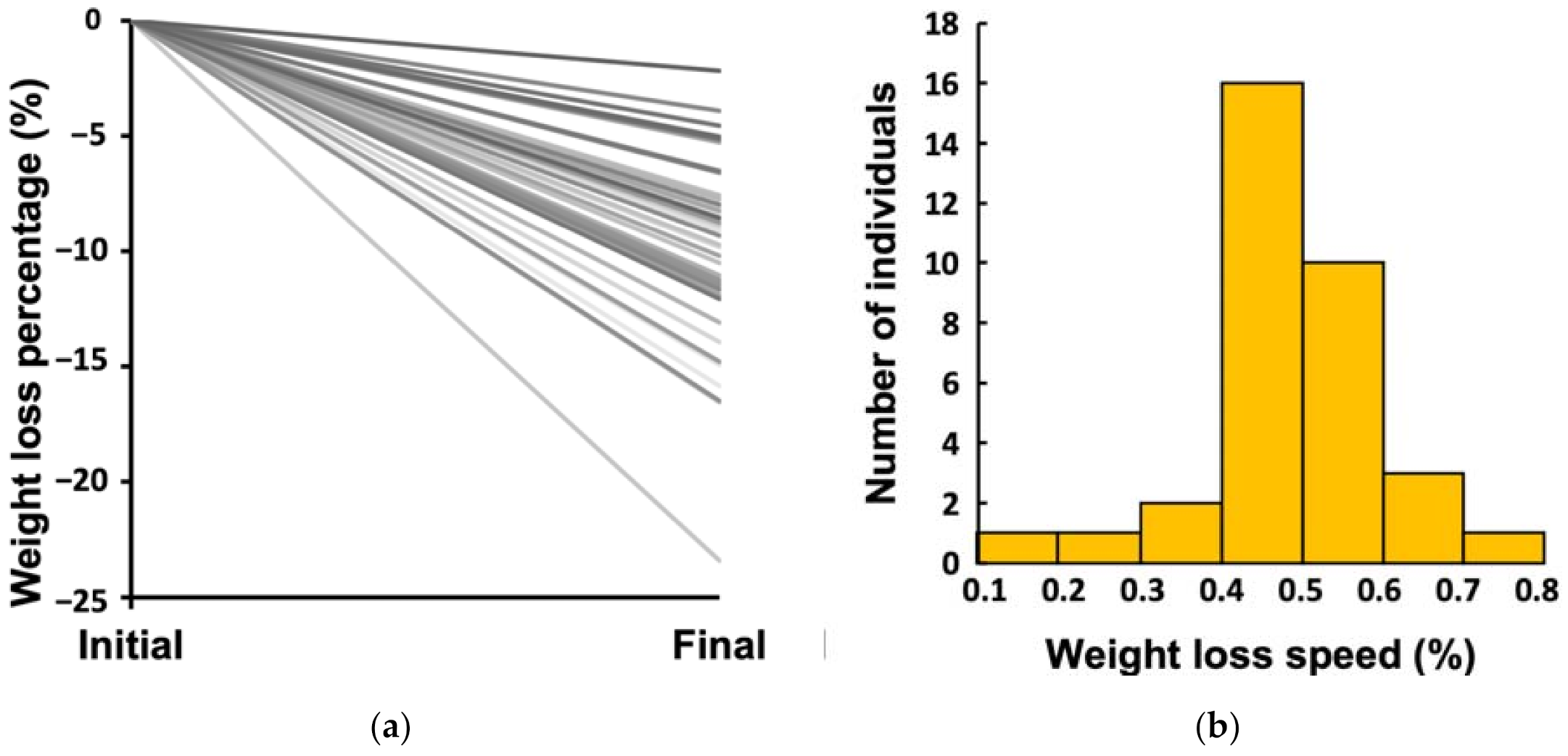
3.1. Large Inter-Individual Variability in Weight Loss Effectiveness
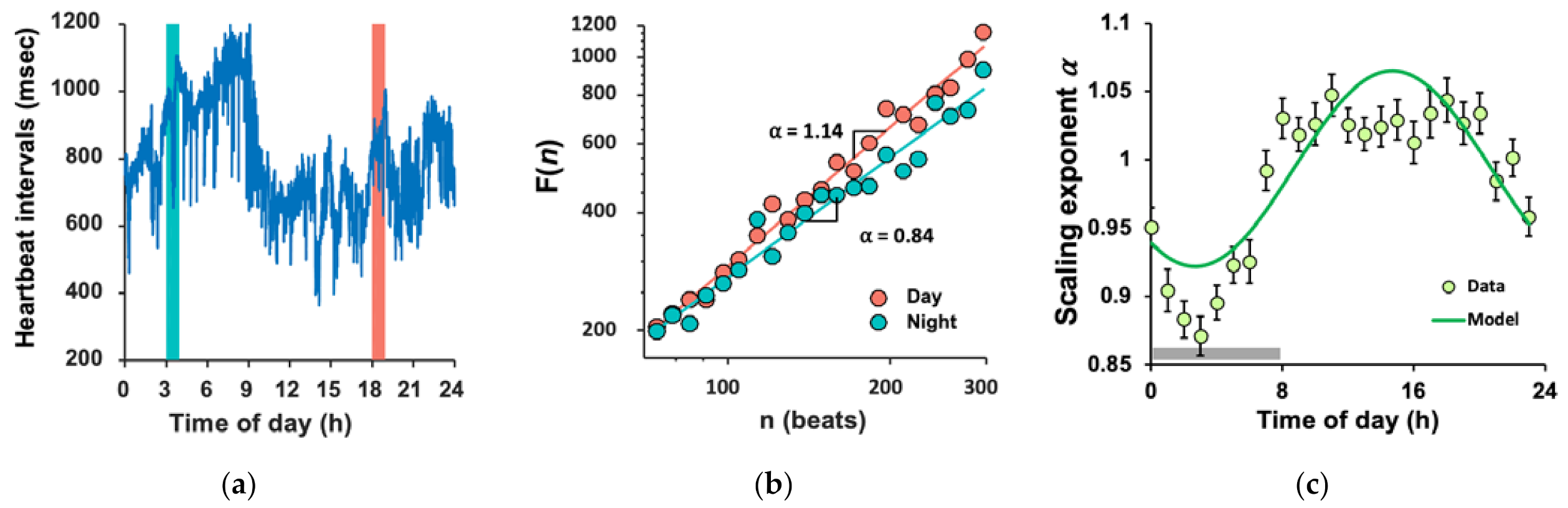
3.2. Daily Rhythm of FCD and Large Inter-Individual Variations
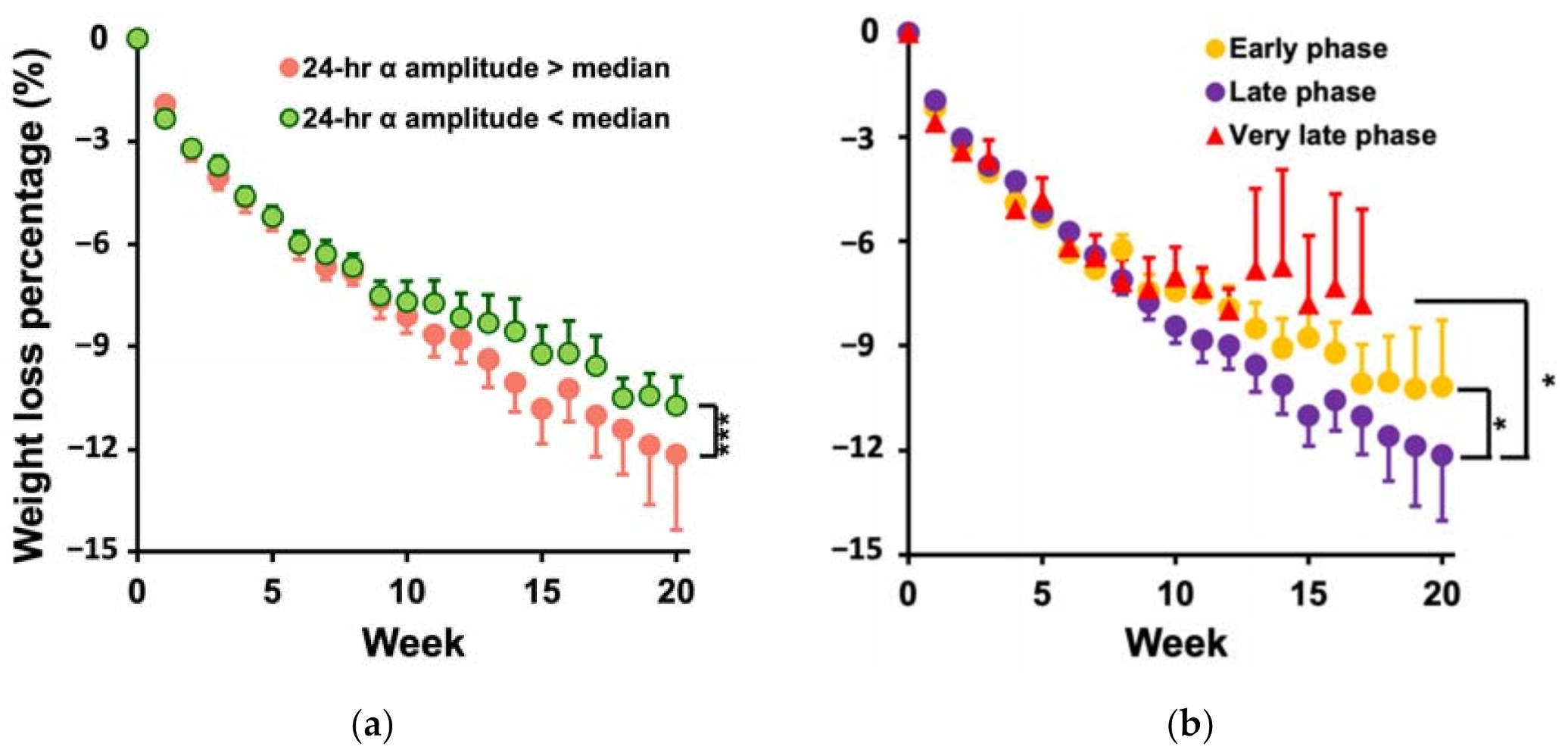
3.3. Participants with Larger Amplitude and the Later Phase of the Daily α Rhythm Lost More Weight
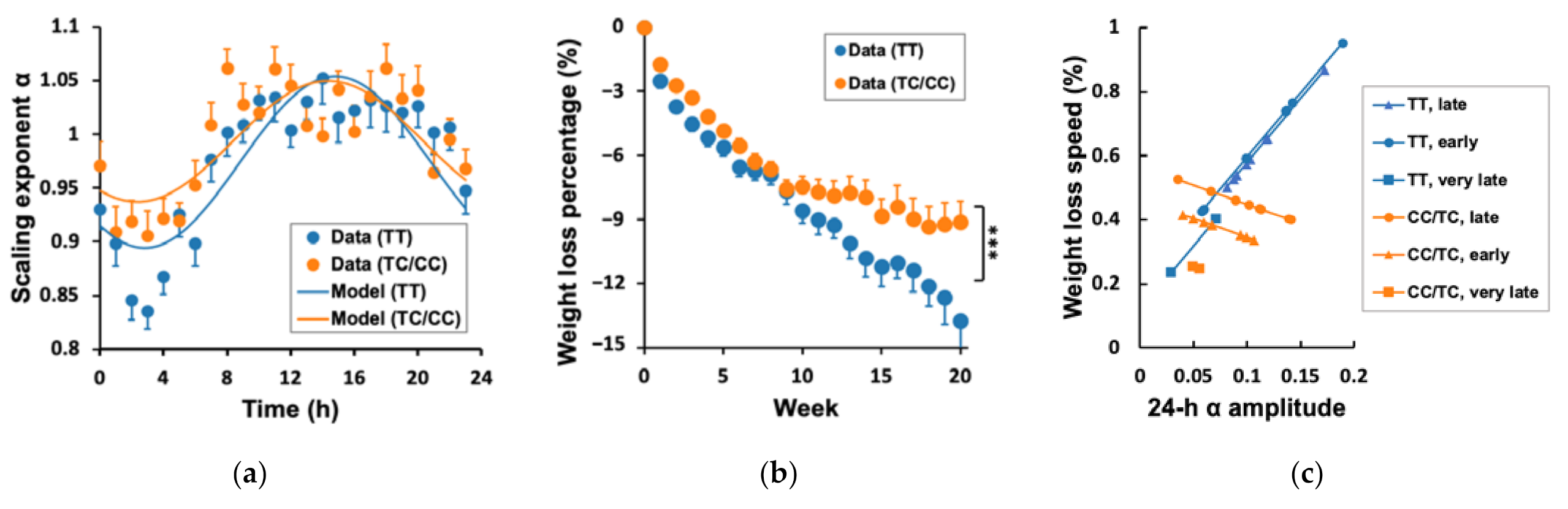
3.4. Genetic Effects on WLS, 24 h α Rhythm, and Their Association
4. Discussion
4.1. Health Implications
4.2. Direction of the Relationship between Daily Rhythm of Fractal Cardiac Dynamics and Weight Loss
4.3. Optimal Circadian Alignment
4.4. Genetics of Weight Loss Resistance
4.5. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The Obesity Epidemic (7:13) | CDC-TV | CDC. Available online: http://www.cdc.gov/cdctv/diseaseandconditions/lifestyle/obesity-epidemic.html (accessed on 7 July 2016).
- Lee, S.J.; Shin, S.W. Mechanisms, Pathophysiology, and Management of Obesity. N. Engl. J. Med. 2017, 376, 1491–1492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alford, S.; Patel, D.; Perakakis, N.; Mantzoros, C.S. Obesity as a Risk Factor for Alzheimer’s Disease: Weighing the Evidence. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2018, 19, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Dansinger, M.L.; Gleason, J.A.; Griffith, J.L.; Selker, H.P.; Schaefer, E.J. Comparison of the Atkins, Ornish, Weight Watchers, and Zone Diets for Weight Loss and Heart Disease Risk Reduction: A Randomized Trial. JAMA 2005, 293, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Corbalán, M.D.; Morales, E.M.; Canteras, M.; Espallardo, A.; Hernández, T.; Garaulet, M. Effectiveness of Cognitive-Behavioral Therapy Based on the Mediterranean Diet for the Treatment of Obesity. Nutrition 2009, 25, 861–869. [Google Scholar] [CrossRef] [PubMed]
- López-Guimerà, G.; Dashti, H.S.; Smith, C.E.; Sánchez-Carracedo, D.; Ordovas, J.M.; Garaulet, M. CLOCK 3111 T/C SNP Interacts with Emotional Eating Behavior for Weight-Loss in a Mediterranean Population. PLoS ONE 2014, 9, e99152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solmi, M.; Gallicchio, D.; Collantoni, E.; Meneguzzo, P.; Zanetti, T.; Degortes, D.; Tenconi, E.; Bonello, E.; Veronese, A.; Ronzan, A.; et al. The Impact of Weight Suppression and Weight Loss Speed on Baseline Clinical Characteristics and Response to Treatment. Int. J. Eat. Disord. 2018, 51, 542–548. [Google Scholar] [CrossRef] [PubMed]
- Lean, M.E.J.; Astrup, A.; Roberts, S.B. Making Progress on the Global Crisis of Obesity and Weight Management. BMJ 2018, 361. [Google Scholar] [CrossRef] [Green Version]
- Reppert, S.M.; Weaver, D.R. Molecular Analysis of Mammalian Circadian Rhythms. Annu. Rev. Physiol. 2001, 63, 647–676. [Google Scholar] [CrossRef]
- Kalsbeek, A.; Kreier, F.; Fliers, E.; Sauerwein, H.P.; Romijn, J.A.; Buijs, R.M. Minireview: Circadian Control of Metabolism by the Suprachiasmatic Nuclei. Endocrinology 2007, 148, 5635–5639. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Ma, J.; Yao, K.; Su, W.; Tan, B.; Wu, X.; Huang, X.; Li, T.; Yin, Y.; Tosini, G.; et al. Circadian Rhythms and Obesity: Timekeeping Governs Lipid Metabolism. J. Pineal Res. 2020, 69. [Google Scholar] [CrossRef]
- McHill, A.W.; Wright, K.P. Role of Sleep and Circadian Disruption on Energy Expenditure and in Metabolic Predisposition to Human Obesity and Metabolic Disease: Sleep Disruption and Circadian Misalignment. Obes. Rev. 2017, 18, 15–24. [Google Scholar] [CrossRef]
- Alibhai, F.J.; LaMarre, J.; Reitz, C.J.; Tsimakouridze, E.V.; Kroetsch, J.T.; Bolz, S.-S.; Shulman, A.; Steinberg, S.; Burris, T.P.; Oudit, G.Y.; et al. Disrupting the Key Circadian Regulator CLOCK Leads to Age-Dependent Cardiovascular Disease. J. Mol. Cell. Cardiol. 2017, 105, 24–37. [Google Scholar] [CrossRef] [Green Version]
- Chellappa, S.L.; Vujovic, N.; Williams, J.S.; Scheer, F.A.J.L. Impact of Circadian Disruption on Cardiovascular Function and Disease. Trends Endocrinol. Metab. 2019, 30, 767–779. [Google Scholar] [CrossRef]
- Zerón-Rugerio, M.; Hernáez, Á.; Porras-Loaiza, A.; Cambras, T.; Izquierdo-Pulido, M. Eating Jet Lag: A Marker of the Variability in Meal Timing and Its Association with Body Mass Index. Nutrients 2019, 11, 2980. [Google Scholar] [CrossRef] [Green Version]
- Thomas, E.A.; Zaman, A.; Cornier, M.-A.; Catenacci, V.A.; Tussey, E.J.; Grau, L.; Arbet, J.; Broussard, J.L.; Rynders, C.A. Later Meal and Sleep Timing Predicts Higher Percent Body Fat. Nutrients 2020, 13, 73. [Google Scholar] [CrossRef]
- Parr, E.B.; Devlin, B.L.; Radford, B.E.; Hawley, J.A. A Delayed Morning and Earlier Evening Time-Restricted Feeding Protocol for Improving Glycemic Control and Dietary Adherence in Men with Overweight/Obesity: A Randomized Controlled Trial. Nutrients 2020, 12, 505. [Google Scholar] [CrossRef] [Green Version]
- Ozburn, A.R.; Purohit, K.; Parekh, P.K.; Kaplan, G.N.; Falcon, E.; Mukherjee, S.; Cates, H.M.; McClung, C.A. Functional Implications of the CLOCK 3111T/C Single-Nucleotide Polymorphism. Front. Psychiatry 2016, 7, 67. [Google Scholar] [CrossRef] [Green Version]
- Sookoian, S.; Gemma, C.; Gianotti, T.F.; Burgueño, A.; Castaño, G.; Pirola, C.J. Genetic Variants of Clock Transcription Factor Are Associated with Individual Susceptibility to Obesity. Am. J. Clin. Nutr. 2008, 87, 1606–1615. [Google Scholar] [CrossRef] [Green Version]
- Garaulet, M.; Lee, Y.-C.; Shen, J.; Parnell, L.D.; Arnett, D.K.; Tsai, M.Y.; Lai, C.-Q.; Ordovas, J.M. CLOCK Genetic Variation and Metabolic Syndrome Risk: Modulation by Monounsaturated Fatty Acids. Am. J. Clin. Nutr. 2009, 90, 1466–1475. [Google Scholar] [CrossRef] [Green Version]
- Garaulet, M.; Corbalán, M.D.; Madrid, J.A.; Morales, E.; Baraza, J.C.; Lee, Y.C.; Ordovas, J.M. CLOCK Gene Is Implicated in Weight Reduction in Obese Patients Participating in a Dietary Programme Based on the Mediterranean Diet. Int. J. Obes. 2010, 34, 516–523. [Google Scholar] [CrossRef] [Green Version]
- Hu, K.; Ivanov, P.C.; Hilton, M.F.; Chen, Z.; Ayers, R.T.; Stanley, H.E.; Shea, S.A. Endogenous Circadian Rhythm in an Index of Cardiac Vulnerability Independent of Changes in Behavior. Proc. Natl. Acad. Sci. USA 2004, 101, 18223–18227. [Google Scholar] [CrossRef] [Green Version]
- Scheer, F.A.J.L.; Hu, K.; Evoniuk, H.; Kelly, E.E.; Malhotra, A.; Hilton, M.F.; Shea, S.A. Impact of the Human Circadian System, Exercise, and Their Interaction on Cardiovascular Function. Proc. Natl. Acad. Sci. USA 2010, 107, 20541–20546. [Google Scholar] [CrossRef] [Green Version]
- Hu, K.; Scheer, F.A.J.L.; Laker, M.; Smales, C.; Shea, S.A. Endogenous Circadian Rhythm in Vasovagal Response to Head-Up Tilt. Circulation 2011, 123, 961–970. [Google Scholar] [CrossRef]
- Messina, G.; De Luca, V.; Viggiano, A.; Ascione, A.; Iannaccone, T.; Chieffi, S.; Monda, M. Autonomic Nervous System in the Control of Energy Balance and Body Weight: Personal Contributions. Neurol. Res. Int. 2013, 2013, 639280. [Google Scholar] [CrossRef]
- Arone, L.J.; Mackintosh, R.; Rosenbaum, M.; Leibel, R.L.; Hirsch, J. Autonomic Nervous System Activity in Weight Gain and Weight Loss. Am. J. Physiol. 1995, 269, R222–R225. [Google Scholar] [CrossRef]
- Guarino, D.; Nannipieri, M.; Iervasi, G.; Taddei, S.; Bruno, R.M. The Role of the Autonomic Nervous System in the Pathophysiology of Obesity. Front. Physiol. 2017, 8, 665. [Google Scholar] [CrossRef] [Green Version]
- Goldberger, A.L.; Amaral, L.A.N.; Hausdorff, J.M.; Ivanov, P.C.; Peng, C.-K.; Stanley, H.E. Fractal Dynamics in Physiology: Alterations with Disease and Aging. Proc. Natl. Acad. Sci. USA 2002, 99 (Suppl. 1), 2466–2472. [Google Scholar] [CrossRef] [Green Version]
- Tulppo, M.P.; Kiviniemi, A.M.; Hautala, A.J.; Kallio, M.; Seppänen, T.; Mäkikallio, T.H.; Huikuri, H.V. Physiological Background of the Loss of Fractal Heart Rate Dynamics. Circulation 2005, 112, 314–319. [Google Scholar] [CrossRef] [Green Version]
- Castiglioni, P.; Merati, G. Fractal Analysis of Heart Rate Variability Reveals Alterations of the Integrative Autonomic Control of Circulation in Paraplegic Individuals. Physiol. Meas. 2017, 38, 774–786. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, P.C.; Bunde, A.; Amaral, L.A.; Havlin, S.; Fritsch-Yelle, J.; Baevsky, R.M.; Stanley, H.E.; Goldberger, A.L. Sleep-Wake Differences in Scaling Behavior of the Human Heartbeat: Analysis of Terrestrial and Long-Term Space Flight Data. Europhys. Lett. 1999, 48, 594–600. [Google Scholar] [CrossRef] [Green Version]
- Mäkikallio, T.H.; Huikuri, H.V.; Mäkikallio, A.; Sourander, L.B.; Mitrani, R.D.; Castellanos, A.; Myerburg, R.J. Prediction of Sudden Cardiac Death by Fractal Analysis of Heart Rate Variability in Elderly Subjects. J. Am. Coll. Cardiol. 2001, 37, 1395–1402. [Google Scholar] [CrossRef] [Green Version]
- Mäkikallio, A.M.; Mäkikallio, T.H.; Korpelainen, J.T.; Sotaniemi, K.A.; Huikuri, H.V.; Myllylä, V.V. Heart Rate Dynamics Predict Poststroke Mortality. Neurology 2004, 62, 1822–1826. [Google Scholar] [CrossRef] [PubMed]
- Stein, P.K.; Domitrovich, P.P.; Huikuri, H.V.; Kleiger, R.E. Cast Investigators Traditional and Nonlinear Heart Rate Variability Are Each Independently Associated with Mortality after Myocardial Infarction. J. Cardiovasc. Electrophysiol. 2005, 16, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Shea, T.J.; Marks, J.; Scheer, F.A.; Shea, S.A. Altered Scale-Invariant Cardiac Dynamics in Humans Across a Forced Desynchrony Protocol. Sleep 2011, 34, A58. [Google Scholar]
- Garaulet, M.; Vera, B.; Bonnet-Rubio, G.; Gómez-Abellán, P.; Lee, Y.-C.; Ordovás, J.M. Lunch Eating Predicts Weight-Loss Effectiveness in Carriers of the Common Allele at PERILIPIN1: The ONTIME (Obesity, Nutrigenetics, Timing, Mediterranean) Study. Am. J. Clin. Nutr. 2016, 104, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garaulet, M.; Sánchez-Moreno, C.; Smith, C.E.; Lee, Y.-C.; Nicolás, F.; Ordovás, J.M. Ghrelin, Sleep Reduction and Evening Preference: Relationships to CLOCK 3111 T/C SNP and Weight Loss. PLoS ONE 2011, 6, e17435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hercberg, S.; Preziosi, P.; Galan, P. Iron Deficiency in Europe. Pub. Health Nutr. 2001, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mataix Verdu, J.; Manas Almendros, M.; Llopis Gonzalez, J.; Martinez de Victoria Munoz, E. Tabla de Composicion de Alimentos Espanoles. 1995. Available online: https://agris.fao.org/agris-search/search.do?recordID=XF2015033769 (accessed on 16 July 2016).
- Roman-Viñas, B.; Serra-Majem, L.; Hagströmer, M.; Ribas-Barba, L.; Sjöström, M.; Segura-Cardona, R. International Physical Activity Questionnaire: Reliability and Validity in a Spanish Population. Eur. J. Sport Sci. 2010, 10, 297–304. [Google Scholar] [CrossRef]
- Craig, C.L.; Marshall, A.L.; Sjöström, M.; Bauman, A.E.; Booth, M.L.; Ainsworth, B.E.; Pratt, M.; Ekelund, U.; Yngve, A.; Sallis, J.F.; et al. International Physical Activity Questionnaire: 12-Country Reliability and Validity. Med. Sci. Sports Exerc. 2003, 35, 1381–1395. [Google Scholar] [CrossRef] [Green Version]
- Task Force of the European Society of Cardiology; North American Society of Pacing and Electrophysiology. HEART RATE VARIABILITY. Standards of Measurement, Physiological Interpretation, and Clinical Use. Circulation 1996, 93, 1043–1065. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Jeong, K.; Yoon, J.; Lee, M. A Simple Real-Time QRS Detection Algorithm. In Proceedings of the 18th Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Amsterdam, The Netherlands, 31 October–3 November 1996; Volume 4, pp. 1396–1398. [Google Scholar]
- Peng, C.K.; Havlin, S.; Hausdorff, J.M.; Mietus, J.E.; Stanley, H.E.; Goldberger, A.L. Fractal Mechanisms and Heart Rate Dynamics. Long-Range Correlations and Their Breakdown with Disease. J. Electrocardiol. 1995, 28 (Suppl. 1), 59–65. [Google Scholar] [CrossRef]
- Peng, C.-K.; Havlin, S.; Stanley, H.E.; Goldberger, A.L. Quantification of Scaling Exponents and Crossover Phenomena in Nonstationary Heartbeat Time Series. Chaos Interdiscip. J. Nonlinear Sci. 1995, 5, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Bunde, A.; Havlin, S.; Kantelhardt, J.W.; Penzel, T.; Peter, J.-H.; Voigt, K. Correlated and Uncorrelated Regions in Heart-Rate Fluctuations during Sleep. Phys. Rev. Lett. 2000, 85, 3736–3739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, M.-T.; Bandin, C.; Yang, H.-W.; Scheer, F.A.; Hu, K.; Garaulet, M. CLOCK 3111T/C Genetic Variant Influences the Daily Rhythm of Autonomic Nervous Function: Relevance to Body Weight Control. Int. J. Obes. 2018, 42, 190–197. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.-W.; Garaulet, M.; Li, P.; Bandin, C.; Lin, C.; Lo, M.-T.; Kun, H. Supplemental Figures and Tables. Figshare Digit. Repos. 2021. [Google Scholar] [CrossRef]
- Anderson, J.W.; Konz, E.C.; Frederich, R.C.; Wood, C.L. Long-Term Weight-Loss Maintenance: A Meta-Analysis of US Studies. Am. J. Clin. Nutr. 2001, 74, 579–584. [Google Scholar] [CrossRef]
- Froy, O.; Garaulet, M. The Circadian Clock in White and Brown Adipose Tissue: Mechanistic, Endocrine, and Clinical Aspects. Endocr. Rev. 2018, 39, 261–273. [Google Scholar] [CrossRef]
- Hiller-Sturmhöfel, S.; Bartke, A. The Endocrine System. Alcohol Health Res. World 1998, 22, 153–164. [Google Scholar]
- Buijs, R.M.; Kalsbeek, A. Hypothalamic Integration of Central and Peripheral Clocks. Nat. Rev. Neurosci. 2001, 2, 521–526. [Google Scholar] [CrossRef]
- Buijs, R.M.; Escobar, C.; Swaab, D.F. The Circadian System and the Balance of the Autonomic Nervous System. Handb. Clin. Neurol. 2013, 117, 173–191. [Google Scholar] [CrossRef]
- Sun, M.; Feng, W.; Wang, F.; Li, P.; Li, Z.; Li, M.; Tse, G.; Vlaanderen, J.; Vermeulen, R.; Tse, L.A. Meta-Analysis on Shift Work and Risks of Specific Obesity Types. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2018, 19, 28–40. [Google Scholar] [CrossRef]
- Furlan, R.; Barbic, F.; Piazza, S.; Tinelli, M.; Seghizzi, P.; Malliani, A. Modifications of Cardiac Autonomic Profile Associated with a Shift Schedule of Work. Circulation 2000, 102, 1912–1916. [Google Scholar] [CrossRef]
- Sawamoto, R.; Nozaki, T.; Furukawa, T.; Tanahashi, T.; Morita, C.; Hata, T.; Komaki, G.; Sudo, N. Higher Sleep Fragmentation Predicts a Lower Magnitude of Weight Loss in Overweight and Obese Women Participating in a Weight-Loss Intervention. Nutr. Diabetes 2014, 4, e144. [Google Scholar] [CrossRef] [Green Version]
- Dean, D.A.; Goldberger, A.L.; Mueller, R.; Kim, M.; Rueschman, M.; Mobley, D.; Sahoo, S.S.; Jayapandian, C.P.; Cui, L.; Morrical, M.G.; et al. Scaling Up Scientific Discovery in Sleep Medicine: The National Sleep Research Resource. Sleep 2016, 39, 1151–1164. [Google Scholar] [CrossRef] [Green Version]
- Choquet, H.; Meyre, D. Genetics of Obesity: What Have We Learned? Curr. Genom. 2011, 12, 169–179. [Google Scholar] [CrossRef] [Green Version]
- Rohde, K.; Keller, M.; Poulsen, L. la C.; Blueher, M.; Kovacs, P.; Bottcher, Y. Genetics and Epigenetics in Obesity. Metab. Clin. Exp. 2019, 92, 37–50. [Google Scholar] [CrossRef] [Green Version]
- Garaulet, M.; Esteban Tardido, A.; Lee, Y.-C.; Smith, C.E.; Parnell, L.D.; Ordovás, J.M. SIRT1 and CLOCK 3111T> C Combined Genotype Is Associated with Evening Preference and Weight Loss Resistance in a Behavioral Therapy Treatment for Obesity. Int. J. Obes. 2012, 36, 1436–1441. [Google Scholar] [CrossRef] [Green Version]
- Giovaninni, N.P.; Fuly, J.T.; Moraes, L.I.; Coutinho, T.N.; Trarbach, E.B.; Jorge, A.A.d.L.; Costalonga, E.F. Study of the Association between 3111T/C Polymorphism of the CLOCK Gene and the Presence of Overweight in Schoolchildren. J. Pediatr. 2014, 90, 500–505. [Google Scholar] [CrossRef] [Green Version]
- Galbete, C.; Contreras, R.; Martínez, J.A.; Martínez-González, M.Á.; Guillén-Grima, F.; Marti, A. Physical Activity and Sex Modulate Obesity Risk Linked to 3111T/C Gene Variant of the CLOCK Gene in an Elderly Population: The SUN Project. Chronobiol. Int. 2012, 29, 1397–1404. [Google Scholar] [CrossRef]
- Monteleone, P.; Tortorella, A.; Docimo, L.; Maldonato, M.N.; Canestrelli, B.; De Luca, L.; Maj, M. Investigation of 3111T/C Polymorphism of the CLOCK Gene in Obese Individuals with or without Binge Eating Disorder: Association with Higher Body Mass Index. Neurosci. Lett. 2008, 435, 30–33. [Google Scholar] [CrossRef]
- Kreier, F.; Yilmaz, A.; Kalsbeek, A.; Romijn, J.A.; Sauerwein, H.P.; Fliers, E.; Buijs, R.M. Hypothesis: Shifting the Equilibrium from Activity to Food Leads to Autonomic Unbalance and the Metabolic Syndrome. Diabetes 2003, 52, 2652–2656. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CLOCK 3111T/C | |||
|---|---|---|---|
| Risk Carriers (TC + CC) | Non-Risk Carriers (TT) | Group Difference | |
| n = 20 | n = 20 | ||
| Obesity Parameters | Mean ± SD | Mean ± SD | p-Value |
| Age (y) | 42.0 ± 9.7 | 46.5 ± 11.04 | NS |
| Height (m) | 1.63 ± 0.06 | 1.63 ± 0.05 | NS |
| Weight (kg) | 74.87 ± 8.83 | 76.32 ± 12.00 | NS |
| BMI (kg/m2) | 28.14 ± 2.41 | 28.66 ± 4.41 | NS |
| Waist (cm) | 89.10 ± 9.55 | 91.60 ± 10.80 | NS |
| Hip (cm) | 105.65 ± 8.72 | 110.25 ± 10.50 | NS |
| Body Fat (%) | 35.03 ± 3.72 | 34.81 ± 6.26 | NS |
| Glucose (mg/dL) | 84.80 ± 20.47 | 87.11 ± 10.16 | NS |
| Insulin (µUI/mL) | 10.27 ± 13.99 | 6.03 ± 3.16 | NS |
| Total Triglycerides (mg/dL) | 104.35 ± 43.25 | 87.00 ± 28.90 | NS |
| Total Cholesterol (mg/dL) | 198.10 ± 24.67 | 194.83 ± 26.08 | NS |
| Cholesterol HDL (mg/dL) | 57.80 ± 12.99 | 54.33 ± 14.36 | NS |
| Cholesterol LDL (mg/dL) | 119.45 ± 18.65 | 123.03 ± 26.90 | NS |
| Leptin (ng/mL) | 21.14 ± 9.65 | 20.34 ± 9.32 | NS |
| Adiponectin (ng/mL) | 74.60 ± 26.62 | 78.70 ± 25.24 | NS |
| IL6 (µg/mL) | 24.79 ± 5.90 | 24.81 ± 6.18 | NS |
| Total Energy Intake (kcal/day) | 1980 ± 767 | 1893 ± 601 | NS |
| Physical Activity Test (METs) | 3614 ± 2802 | 2842 ± 2886 | NS |
| Sleep Duration (hours) | 7.8 ± 0.6 | 7.4 ± 0.8 | NS |
| Bedtime | 0:36 ± 55 (min) | 0:21 ± 47 (min) | NS |
| Wake-Up Time | 8:18 ± 46 (min) | 7:47 ± 38 (min) | 0.028 |
| Breakfast Time | 9:30 ± 64 (min) | 8:35 ± 48 (min) | 0.004 |
| Lunch Time | 14:51 ± 29 (min) | 14:20 ± 23 (min) | NS |
| Dinner Time | 21:32 ± 32 (min) | 21:25 ± 24 (min) | NS |
| Intervention Duration (Weeks) | 16.9 ± 6.6 | 16.8 ± 9.4 * | NS |
| Total Weight Loss (Kg) | 7.18 ± 2.20 | 9.02 ± 4.56 * | NS |
| Medication Use (Total) | 9 | 9 | NS |
| Anti-hypertensives | 2 | 2 | NS |
| Lipid Lowering Drug | 1 | 1 | NS |
| Anti-diabetics | 0 | 1 | NS |
| Antidepressants | 3 | 4 | NS |
| Anxiolytics | 2 | 3 | NS |
| Thyroid Hormone | 3 | 0 | 0.036 |
| Contraceptives | 1 | 3 | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.-W.; Garaulet, M.; Li, P.; Bandin, C.; Lin, C.; Lo, M.-T.; Hu, K. Daily Rhythm of Fractal Cardiac Dynamics Links to Weight Loss Resistance: Interaction with CLOCK 3111T/C Genetic Variant. Nutrients 2021, 13, 2463. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13072463
Yang H-W, Garaulet M, Li P, Bandin C, Lin C, Lo M-T, Hu K. Daily Rhythm of Fractal Cardiac Dynamics Links to Weight Loss Resistance: Interaction with CLOCK 3111T/C Genetic Variant. Nutrients. 2021; 13(7):2463. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13072463
Chicago/Turabian StyleYang, Hui-Wen, Marta Garaulet, Peng Li, Cristina Bandin, Chen Lin, Men-Tzung Lo, and Kun Hu. 2021. "Daily Rhythm of Fractal Cardiac Dynamics Links to Weight Loss Resistance: Interaction with CLOCK 3111T/C Genetic Variant" Nutrients 13, no. 7: 2463. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13072463