
The Role of Cell Proliferation and Extracellular Matrix Accumulation Induced by Food Additive Butylated Hydroxytoluene in Uterine Leiomyoma


 , , , ,
, , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Treatments
2.2. Cell Viability Assay
2.3. Colony Formation
2.4. Immunofluorescence
2.5. Protein Preparation and Western Blot
2.6. Statistical Analysis
3. Results
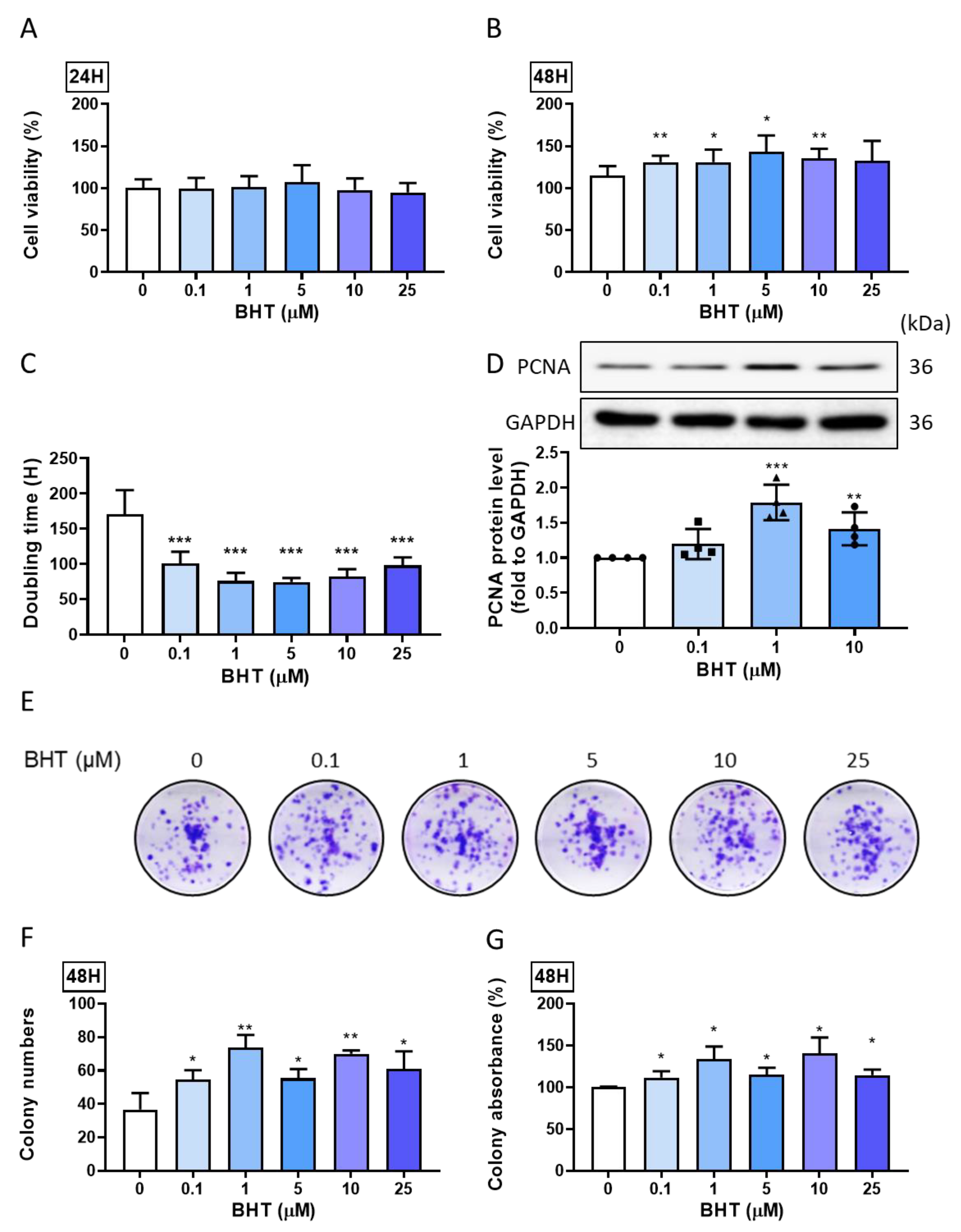
3.1. Effects of BHT on Leiomyoma Proliferation
3.1.1. Proliferative Effect
3.1.2. Colony Formation
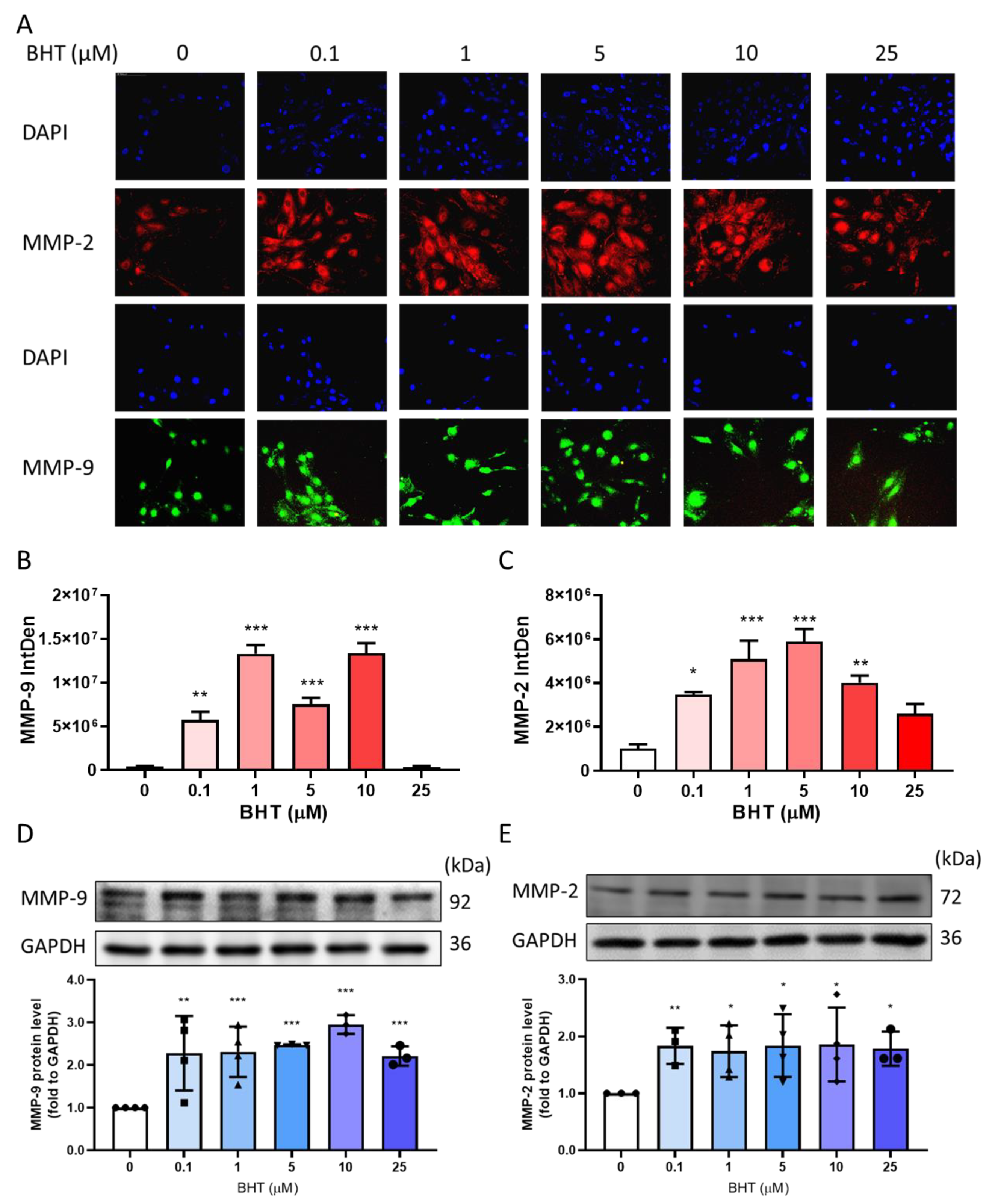
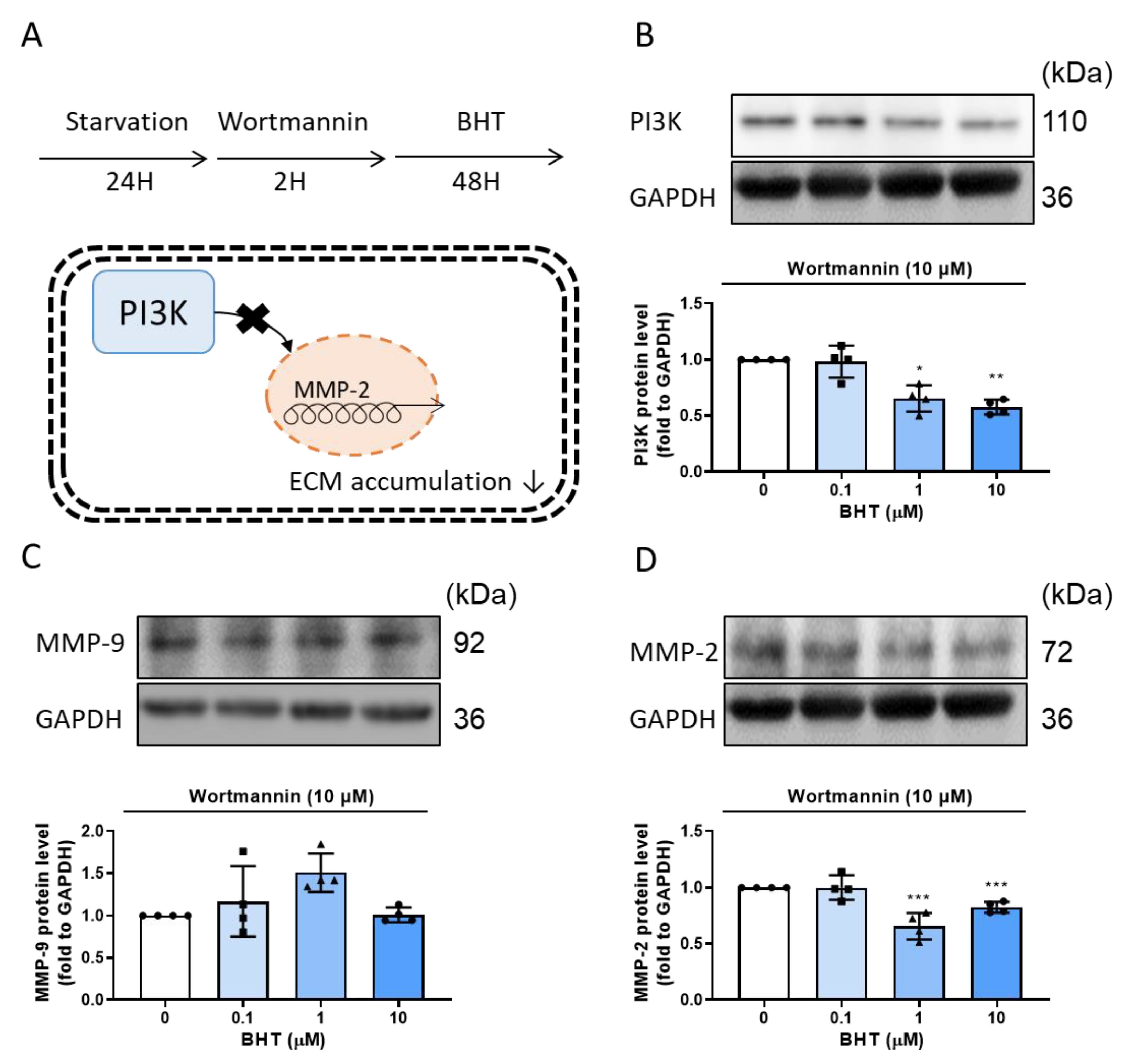
3.2. Effects of BHT on MMP Modulation
3.2.1. Effects of BHT on MMP-9 and MMP-2 Protein Expression Using Immunofluorescence
3.2.2. Effects of BHT on MMP-9 and MMP-2 Protein Expression Using Western Blot
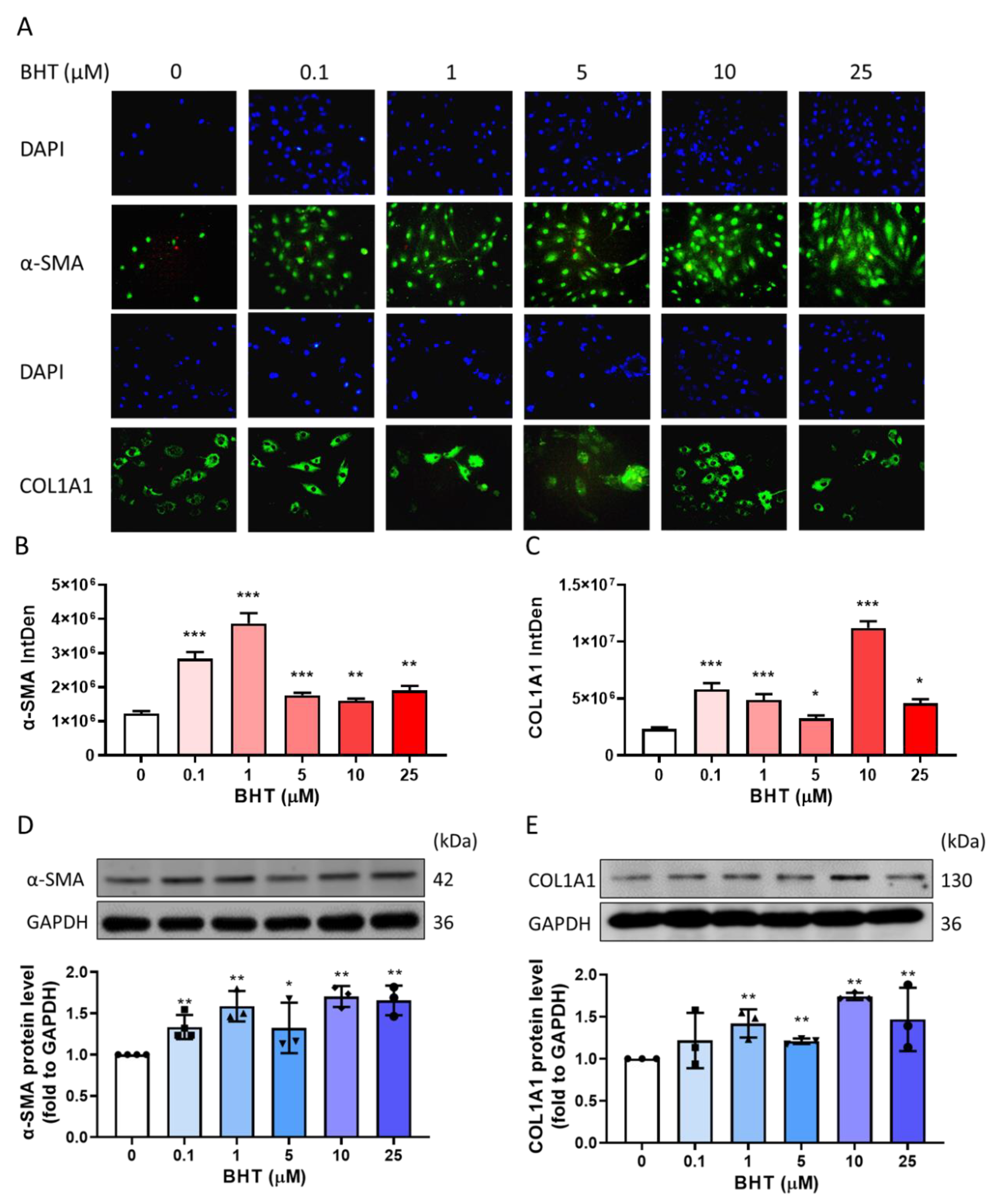
3.3. Effect of BHT on Extracellular Matrix Related Proteins
3.3.1. Effects of BHT on ECM Related Protein Expression Using Immunofluorescence
3.3.2. Effects of BHT on ECM Related Protein Expression Using Western Blot
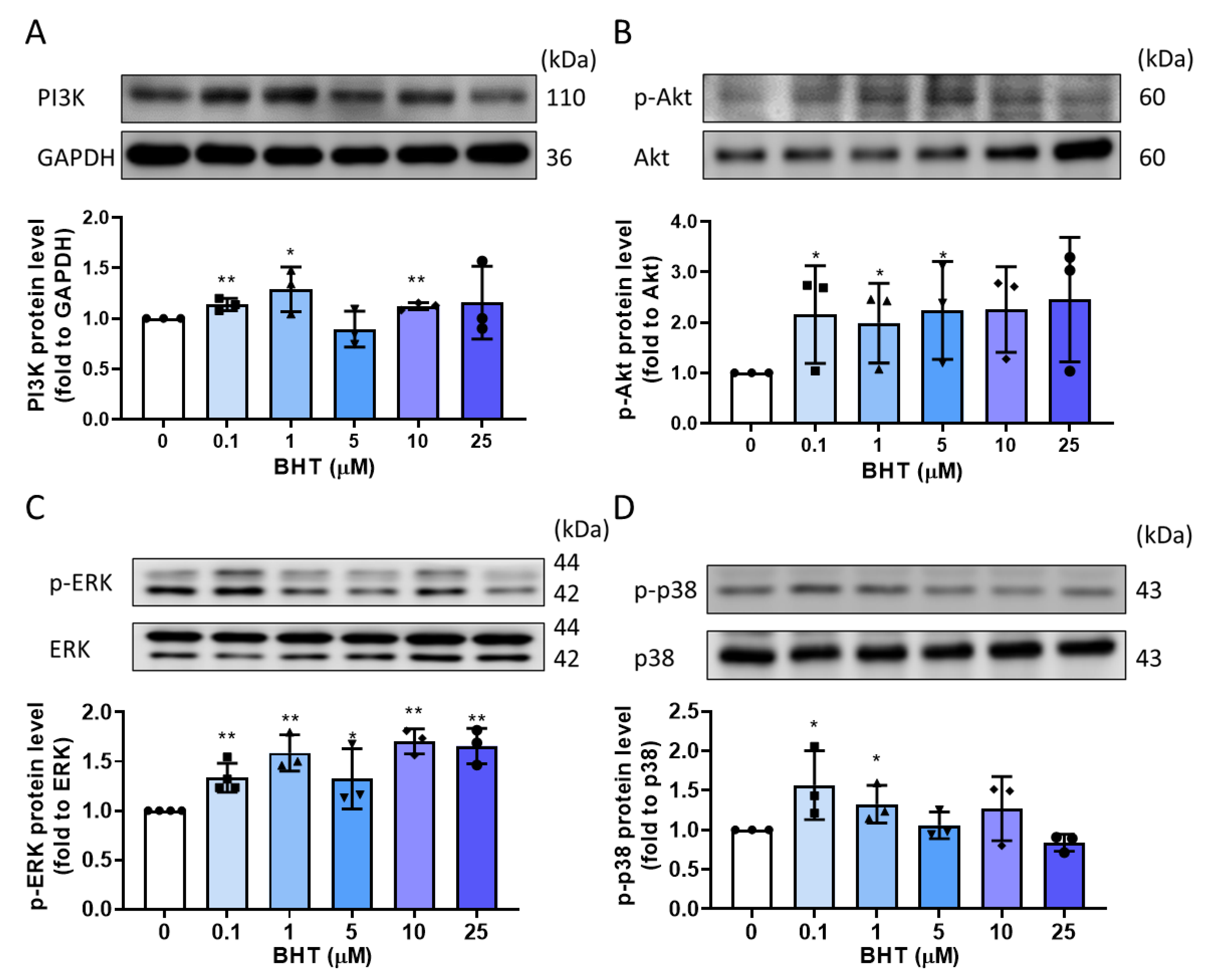
3.4. PI3K/Akt and MAPK Signaling Related Protein Expression Change in BHT Induced ECM Accumulation
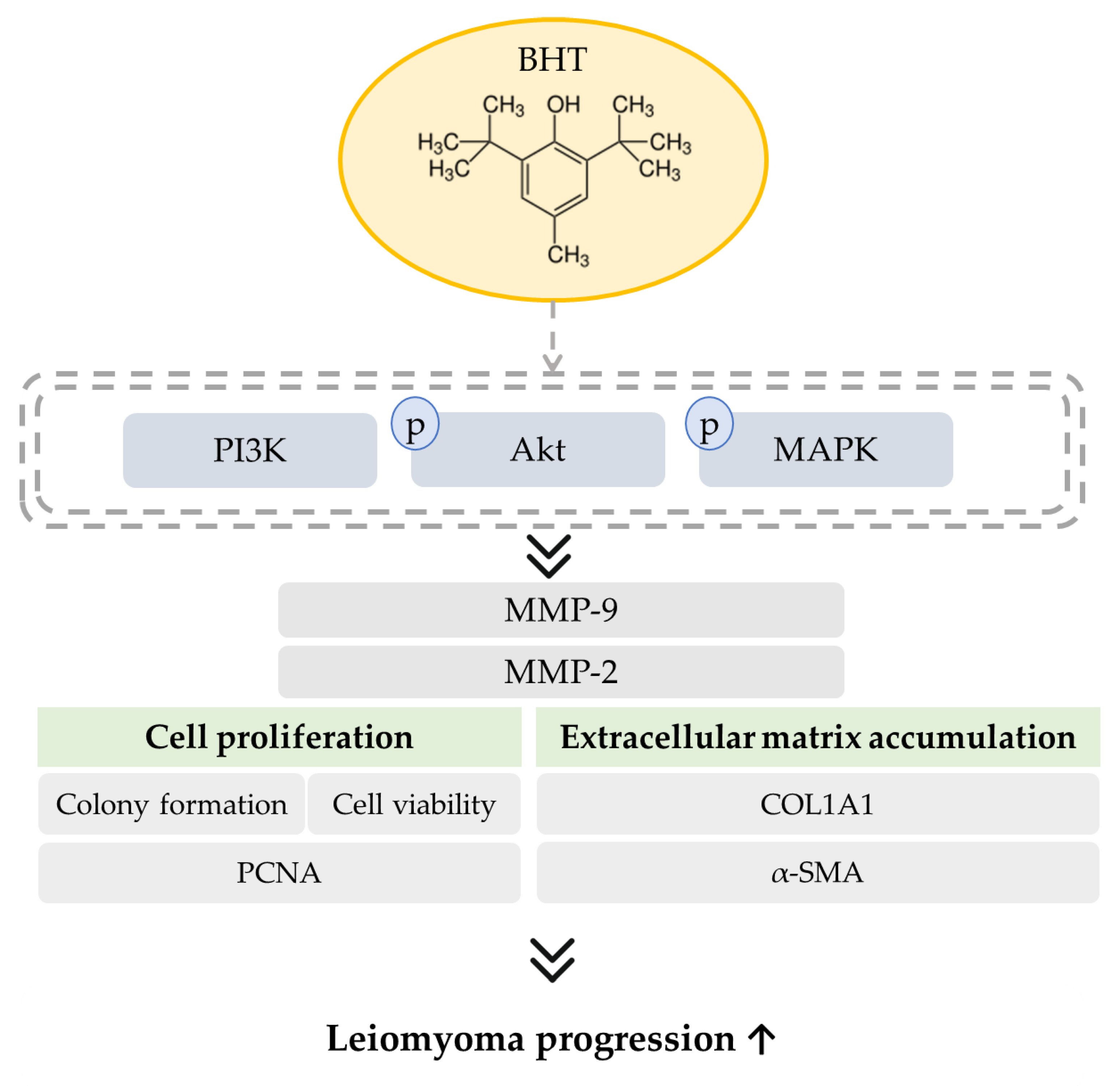
3.5. The Potential Modulative Signaling Pathway in BHT Induced ECM Accumulation
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| α-SMA | Alpha smooth muscle actin |
| BHA | Butylated hydroxyanisole |
| BHT | Butylated hydroxytoluene |
| BSA | Bovine serum albumin |
| COL1A1 | Collagen, type I |
| ECM | Extracellular matrix |
| EDCs | Environmental endocrine-disrupting chemicals |
| h | Hours |
| MAPK | Mitogen-activated protein kinase |
| MMP | Matrix metalloproteinase |
| MnSOD | Manganese superoxide dismutase |
| MTT | (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) |
| PCNA | Proliferating cell nuclear antigen |
| PI3K | Phosphoinositide 3-kinase |
| PKB | Protein kinase B |
| ROS | Reactive oxygen species |
| TIMPs | Tissue inhibitor of metalloproteinase |
References
- Sabry, M.; Al-Hendy, A. Medical treatment of uterine leiomyoma. Reprod. Sci. 2012, 19, 339–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sparic, R.; Mirkovic, L.; Malvasi, A.; Tinelli, A. Epidemiology of Uterine Myomas: A Review. Int. J. Fertil. Steril. 2016, 9, 424–435. [Google Scholar] [PubMed]
- Frascà, C.; Tuzzato, G.; Arena, A.; Degli Esposti, E.; Zanello, M.; Raimondo, D.; Seracchioli, R. The Role of Pelvic Ultrasound in Preoperative Evaluation for Laparoscopic Myomectomy. J. Minim. Invasive Gynecol. 2018, 25, 679–683. [Google Scholar] [CrossRef] [PubMed]
- Munro, M.G.; Critchley, H.O.; Broder, M.S.; Fraser, I.S. FIGO classification system (PALM-COEIN) for causes of abnormal uterine bleeding in nongravid women of reproductive age. Int. J. Gynaecol. Obs. 2011, 113, 3–13. [Google Scholar] [CrossRef]
- Barjon, K.; Mikhail, L.N. Uterine Leiomyomata (Fibroids). In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2019. [Google Scholar]
- Ali, M.; Al-Hendy, A. Selective progesterone receptor modulators for fertility preservation in women with symptomatic uterine fibroids. Biol. Reprod. 2017, 97, 337–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, M.S.; Ciavattini, A.; Petraglia, F.; Castellucci, M.; Ciarmela, P. Extracellular matrix in uterine leiomyoma pathogenesis: A potential target for future therapeutics. Hum. Reprod. Update 2018, 24, 59–85. [Google Scholar] [CrossRef] [Green Version]
- Greco, S.; Islam, M.S.; Zannotti, A.; Delli Carpini, G.; Giannubilo, S.R.; Ciavattini, A.; Petraglia, F.; Ciarmela, P. Quercetin and indole-3-carbinol inhibit extracellular matrix expression in human primary uterine leiomyoma cells. Reprod. Biomed. Online 2020, 40, 593–602. [Google Scholar] [CrossRef]
- Bulun, S.E. Uterine fibroids. N. Engl. J. Med. 2013, 369, 1344–1355. [Google Scholar] [CrossRef] [Green Version]
- Leppert, P.C.; Jayes, F.L.; Segars, J.H. The extracellular matrix contributes to mechanotransduction in uterine fibroids. Obstet Gynecol. Int. 2014, 2014, 783289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wight, T.N.; Potter-Perigo, S. The extracellular matrix: An active or passive player in fibrosis? Am. J. Physiol.-Gastrointest. Liver Physiol. 2011, 301, 950–955. [Google Scholar] [CrossRef] [Green Version]
- Bogusiewicz, M.; Stryjecka-Zimmer, M.; Postawski, K.; Jakimiuk, A.J.; Rechberger, T. Activity of matrix metalloproteinase-2 and -9 and contents of their tissue inhibitors in uterine leiomyoma and corresponding myometrium. Gynecol. Endocrinol. 2007, 23, 541–546. [Google Scholar] [CrossRef]
- Lin, P.-H.; Kung, H.-L.; Chen, H.-Y.; Huang, K.-C.; Hsia, S.-M. Isoliquiritigenin Suppresses E2-Induced Uterine Leiomyoma Growth through the Modulation of Cell Death Program and the Repression of ECM Accumulation. Cancers 2019, 11, 1131. [Google Scholar] [CrossRef] [Green Version]
- Korompelis, P.; Piperi, C.; Adamopoulos, C.; Dalagiorgou, G.; Korkolopoulou, P.; Sepsa, A.; Antsaklis, A.; Papavassiliou, A.G. Expression of vascular endothelial factor-A, gelatinases (MMP-2, MMP-9) and TIMP-1 in uterine leiomyomas. Clin. Chem. Lab. Med. 2015, 53, 1415–1424. [Google Scholar] [CrossRef]
- Hastings, J.F.; Skhinas, J.N.; Fey, D.; Croucher, D.R.; Cox, T.R. The extracellular matrix as a key regulator of intracellular signalling networks. Br. J. Pharm. 2019, 176, 82–92. [Google Scholar] [CrossRef] [Green Version]
- Lin, P.-H.; Tung, Y.-T.; Chen, H.-Y.; Chiang, Y.-F.; Hong, H.-C.; Huang, K.-C.; Hsu, S.-P.; Huang, T.-C.; Hsia, S.-M. Melatonin activates cell death programs for the suppression of uterine leiomyoma cell proliferation. J. Pineal Res. 2019, 68, e12620. [Google Scholar] [CrossRef]
- Yehye, W.A.; Rahman, N.A.; Ariffin, A.; Abd Hamid, S.B.; Alhadi, A.A.; Kadir, F.A.; Yaeghoobi, M. Understanding the chemistry behind the antioxidant activities of butylated hydroxytoluene (BHT): A review. Eur. J. Med. Chem. 2015, 101, 295–312. [Google Scholar] [CrossRef] [PubMed]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharm. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanigan, R.S.; Yamarik, T.A. Final report on the safety assessment of BHT(1). Int. J. Toxicol. 2002, 21 (Suppl. S2), 19–94. [Google Scholar]
- Martins, V.; Teodoro, W.R.; Velosa, A.P.P.; Andrade, P.; Farhat, C.; Fabro, A.T.; Capelozzi, V.L. Butylated hydroxytoluene induces type-V collagen and overexpression of remodeling genes/proteins in experimental lung fibrosis. Histol. Histopathol. 2018, 33, 1111–1123. [Google Scholar]
- Fujita, M.; Shannon, J.M.; Morikawa, O.; Gauldie, J.; Hara, N.; Mason, R.J. Overexpression of tumor necrosis factor-alpha diminishes pulmonary fibrosis induced by bleomycin or transforming growth factor-beta. Am. J. Respir. Cell Mol. Biol. 2003, 29, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A.K.; Dwyer-Nield, L.D. Two-stage 3-methylcholanthrene and butylated hydroxytoluene-induced lung carcinogenesis in mice. Methods Cell Biol. 2021, 163, 153–173. [Google Scholar] [PubMed]
- Chen, H.Y.; Huang, T.C.; Lin, L.C.; Shieh, T.M.; Wu, C.H.; Wang, K.L.; Hong, Y.H.; Hsia, S.M. Fucoidan Inhibits the Proliferation of Leiomyoma Cells and Decreases Extracellular Matrix-Associated Protein Expression. Cell Physiol. Biochem. 2018, 49, 1970–1986. [Google Scholar] [CrossRef]
- Chiang, Y.-F.; Chen, H.-Y.; Huang, K.-C.; Lin, P.-H.; Hsia, S.-M. Dietary Antioxidant Trans-Cinnamaldehyde Reduced Visfatin-Induced Breast Cancer Progression: In Vivo and In Vitro Study. Antioxidants 2019, 8, 625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, Y.-F.; Chen, H.-Y.; Chang, Y.-J.; Shih, Y.-H.; Shieh, T.-M.; Wang, K.-L.; Hsia, S.-M. Protective Effects of Fucoxanthin on High Glucose- and 4-Hydroxynonenal (4-HNE)-Induced Injury in Human Retinal Pigment Epithelial Cells. Antioxidants 2020, 9, 1176. [Google Scholar] [CrossRef]
- Donnini, S.; Morbidelli, L.; Taraboletti, G.; Ziche, M. ERK1-2 and p38 MAPK regulate MMP/TIMP balance and function in response to thrombospondin-1 fragments in the microvascular endothelium. Life Sci. 2004, 74, 2975–2985. [Google Scholar] [CrossRef]
- Wang, G.; Yin, L.; Peng, Y.; Gao, Y.; Gao, H.; Zhang, J.; Lv, N.; Miao, Y.; Lu, Z. Insulin promotes invasion and migration of KRAS(G12D) mutant HPNE cells by upregulating MMP-2 gelatinolytic activity via ERK- and PI3K-dependent signalling. Cell Prolif. 2019, 52, e12575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, S.J.; Kim, J.; Lee, S.; Baek, J.; Lee, J.E.; Cho, C.; Ha, E. Ulipristal acetate induces cell cycle delay and remodeling of extracellular matrix. Int. J. Mol. Med. 2018, 42, 1857–1864. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.; Shahin, S.M.; Sabri, N.A.; Al-Hendy, A.; Yang, Q. 1,25 Dihydroxyvitamin D3 Enhances the Antifibroid Effects of Ulipristal Acetate in Human Uterine Fibroids. Reprod. Sci. 2019, 26, 812–828. [Google Scholar] [CrossRef]
- Borahay, M.A.; Al-Hendy, A.; Kilic, G.S.; Boehning, D. Signaling Pathways in Leiomyoma: Understanding Pathobiology and Implications for Therapy. Mol. Med. 2015, 21, 242–256. [Google Scholar] [CrossRef]
- Islam, M.S.; Greco, S.; Janjusevic, M.; Ciavattini, A.; Giannubilo, S.R.; D’Adderio, A.; Biagini, A.; Fiorini, R.; Castellucci, M.; Ciarmela, P. Growth factors and pathogenesis. Best Pract. Res. Clin. Obs. Gynaecol. 2016, 34, 25–36. [Google Scholar] [CrossRef]
- Bariani, M.V.; Rangaswamy, R.; Siblini, H.; Yang, Q.; Al-Hendy, A.; Zota, A.R. The role of endocrine-disrupting chemicals in uterine fibroid pathogenesis. Curr. Opin. Endocrinol. Diabetes Obes. 2020, 27, 380–387. [Google Scholar] [CrossRef]
- Ishikawa, H.; Ishi, K.; Serna, V.A.; Kakazu, R.; Bulun, S.E.; Kurita, T. Progesterone is essential for maintenance and growth of uterine leiomyoma. Endocrinology 2010, 151, 2433–2442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.; Xu, Q.; Xu, J.; Ren, M.L.; Cai, Y.L. Environmental exposure and risk of uterine leiomyoma: An epidemiologic survey. Eur. Rev. Med. Pharm. Sci. 2013, 17, 3249–3256. [Google Scholar]
- Katz, T.A.; Yang, Q.; Treviño, L.S.; Walker, C.L.; Al-Hendy, A. Endocrine-disrupting chemicals and uterine fibroids. Fertil. Steril. 2016, 106, 967–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagata, C.; Nakamura, K.; Oba, S.; Hayashi, M.; Takeda, N.; Yasuda, K. Association of intakes of fat, dietary fibre, soya isoflavones and alcohol with uterine fibroids in Japanese women. Br. J. Nutr. 2009, 101, 1427–1431. [Google Scholar] [CrossRef] [Green Version]
- Krawczyk, K.; Marynowicz, W.; Gogola-Mruk, J.; Jakubowska, K.; Tworzydło, W.; Opydo-Chanek, M.; Ptak, A. A mixture of persistent organic pollutants detected in human follicular fluid increases progesterone secretion and mitochondrial activity in human granulosa HGrC1 cells. Reprod. Toxicol. 2021, 104, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Lourenço, S.C.; Moldão-Martins, M.; Alves, V.D. Antioxidants of Natural Plant Origins: From Sources to Food Industry Applications. Molecules 2019, 24, 4132. [Google Scholar] [CrossRef] [Green Version]
- Union ECJOJE: Regulation (EC) No 1333/2008 of the European Parliament and of the Council of 16 December 2008 on Food Additives; 2008; Volume 354, pp. 16–33. Available online: http://data.europa.eu/eli/reg/2008/1333/oj (accessed on 30 August 2021).
- Vandghanooni, S.; Forouharmehr, A.; Eskandani, M.; Barzegari, A.; Kafil, V.; Kashanian, S.; Ezzati Nazhad Dolatabadi, J. Cytotoxicity and DNA fragmentation properties of butylated hydroxyanisole. DNA Cell Biol. 2013, 32, 98–103. [Google Scholar] [CrossRef]
- Jeong, S.H.; Kim, B.Y.; Kang, H.G.; Ku, H.O.; Cho, J.H. Effects of butylated hydroxyanisole on the development and functions of reproductive system in rats. Toxicology 2005, 208, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhao, Y.; Liu, W.; Li, Z.; Souders, C.L., 2nd; Martyniuk, C.J. Butylated hydroxytoluene induces hyperactivity and alters dopamine-related gene expression in larval zebrafish (Danio rerio). Environ. Pollut. 2020, 257, 113624. [Google Scholar] [CrossRef] [PubMed]
- Fasihnia, S.H.; Peighambardoust, S.H.; Peighambardoust, S.J.; Oromiehie, A.; Soltanzadeh, M.; Peressini, D. Migration analysis, antioxidant, and mechanical characterization of polypropylene-based active food packaging films loaded with BHA, BHT, and TBHQ. J. Food Sci. 2020, 85, 2317–2328. [Google Scholar] [CrossRef]
- Ousji, O.; Sleno, L. Identification of In Vitro Metabolites of Synthetic Phenolic Antioxidants BHT, BHA, and TBHQ by LC-HRMS/MS. Int. J. Mol. Sci. 2020, 21, 9525. [Google Scholar] [CrossRef]
- Pop, A.; Drugan, T.; Gutleb, A.C.; Lupu, D.; Cherfan, J.; Loghin, F.; Kiss, B. Estrogenic and anti-estrogenic activity of butylparaben, butylated hydroxyanisole, butylated hydroxytoluene and propyl gallate and their binary mixtures on two estrogen responsive cell lines (T47D-Kbluc, MCF-7). J. Appl. Toxicol. 2018, 38, 944–957. [Google Scholar] [CrossRef]
- Ham, J.; Lim, W.; Whang, K.Y.; Song, G. Butylated hydroxytoluene induces dysregulation of calcium homeostasis and endoplasmic reticulum stress resulting in mouse Leydig cell death. Environ. Pollut. 2020, 256, 113421. [Google Scholar] [CrossRef] [PubMed]
- Ough, M.; Lewis, A.; Zhang, Y.; Hinkhouse, M.M.; Ritchie, J.M.; Oberley, L.W.; Cullen, J.J. Inhibition of cell growth by overexpression of manganese superoxide dismutase (MnSOD) in human pancreatic carcinoma. Free Radic. Res. 2004, 38, 1223–1233. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Chae, S.W.; Hur, G.C.; Cho, S.J.; Kim, M.K.; Choi, J.; Nam, S.Y.; Kim, W.H.; Yang, H.K.; Lee, B.L. Manganese superoxide dismutase expression correlates with a poor prognosis in gastric cancer. Pathobiology 2002, 70, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.J.; Zhao, W.; Venkataraman, S.; Robbins, M.E.; Buettner, G.R.; Kregel, K.C.; Oberley, L.W. Activation of matrix metalloproteinase-2 by overexpression of manganese superoxide dismutase in human breast cancer MCF-7 cells involves reactive oxygen species. J. Biol. Chem. 2002, 277, 20919–20926. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.M.; Wu, T.C.; Shieh, S.H.; Wu, Y.H.; Li, M.C.; Sheu, G.T.; Cheng, Y.W.; Chen, C.Y.; Lee, H. MnSOD promotes tumor invasion via upregulation of FoxM1-MMP2 axis and related with poor survival and relapse in lung adenocarcinomas. Mol. Cancer Res. 2013, 11, 261–271. [Google Scholar] [CrossRef] [Green Version]
- Siwik, D.A.; Pagano, P.J.; Colucci, W.S. Oxidative stress regulates collagen synthesis and matrix metalloproteinase activity in cardiac fibroblasts. Am. J. Physiol. Cell Physiol. 2001, 280, C53–C60. [Google Scholar] [CrossRef]
- Cirigliano, M. Bioidentical hormone therapy: A review of the evidence. J. Women’s Health 2007, 16, 600–631. [Google Scholar] [CrossRef] [PubMed]
- Qian, Q.; Wang, Q.; Zhan, P.; Peng, L.; Wei, S.Z.; Shi, Y.; Song, Y. The role of matrix metalloproteinase 2 on the survival of patients with non-small cell lung cancer: A systematic review with meta-analysis. Cancer Investig. 2010, 28, 661–669. [Google Scholar] [CrossRef]
- Yamaki, K.; Taneda, S.; Yanagisawa, R.; Inoue, K.; Takano, H.; Yoshino, S. Enhancement of allergic responses in vivo and in vitro by butylated hydroxytoluene. Toxicol. Appl. Pharm. 2007, 223, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Parra, E.R.; Boufelli, G.; Bertanha, F.; Samorano Lde, P.; Aguiar, A.C., Jr.; Costa, F.M.; Capelozzi, V.L.; Barbas-Filho, J.V. Temporal evolution of epithelial, vascular and interstitial lung injury in an experimental model of idiopathic pulmonary fibrosis induced by butyl-hydroxytoluene. Int. J. Exp. Pathol. 2008, 89, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Umemura, T.; Kodama, Y.; Hioki, K.; Inoue, T.; Nomura, T.; Kurokawa, Y. Butylhydroxytoluene (BHT) increases susceptibility of transgenic rasH2 mice to lung carcinogenesis. J. Cancer Res. Clin. Oncol. 2001, 127, 583–590. [Google Scholar] [CrossRef]
- Parra, E.R.; Pincelli, M.S.; Teodoro, W.R.; Velosa, A.P.P.; Martins, V.; Rangel, M.P.; Barbas-Filho, J.V.; Capelozzi, V.L. Modeling pulmonary fibrosis by abnormal expression of telomerase/apoptosis/collagen V in experimental usual interstitial pneumonia. Braz. J. Med Biol. Res. 2014, 47, 567–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiang, Y.-F.; Chen, H.-Y.; Ali, M.; Shieh, T.-M.; Huang, Y.-J.; Wang, K.-L.; Chang, H.-Y.; Huang, T.-C.; Hong, Y.-H.; Hsia, S.-M. The Role of Cell Proliferation and Extracellular Matrix Accumulation Induced by Food Additive Butylated Hydroxytoluene in Uterine Leiomyoma. Nutrients 2021, 13, 3074. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13093074
Chiang Y-F, Chen H-Y, Ali M, Shieh T-M, Huang Y-J, Wang K-L, Chang H-Y, Huang T-C, Hong Y-H, Hsia S-M. The Role of Cell Proliferation and Extracellular Matrix Accumulation Induced by Food Additive Butylated Hydroxytoluene in Uterine Leiomyoma. Nutrients. 2021; 13(9):3074. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13093074
Chicago/Turabian StyleChiang, Yi-Fen, Hsin-Yuan Chen, Mohamed Ali, Tzong-Ming Shieh, Yun-Ju Huang, Kai-Lee Wang, Hsin-Yi Chang, Tsui-Chin Huang, Yong-Han Hong, and Shih-Min Hsia. 2021. "The Role of Cell Proliferation and Extracellular Matrix Accumulation Induced by Food Additive Butylated Hydroxytoluene in Uterine Leiomyoma" Nutrients 13, no. 9: 3074. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13093074