
High Hydrostatic Pressure Processing of Human Milk Increases Apelin and GLP-1 Contents to Modulate Gut Contraction and Glucose Metabolism in Mice Compared to Holder Pasteurization

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Milk Collection and HoP and HHP Processing
2.2. Quantification of Apelin and GLP-1 in Milk Samples
2.3. Mice
2.4. Oral Glucose Tolerance Test (OGTT)
2.5. Measurement of Isotonic Intestinal Contractions
2.6. Gene Expression
2.7. Statistics
3. Results
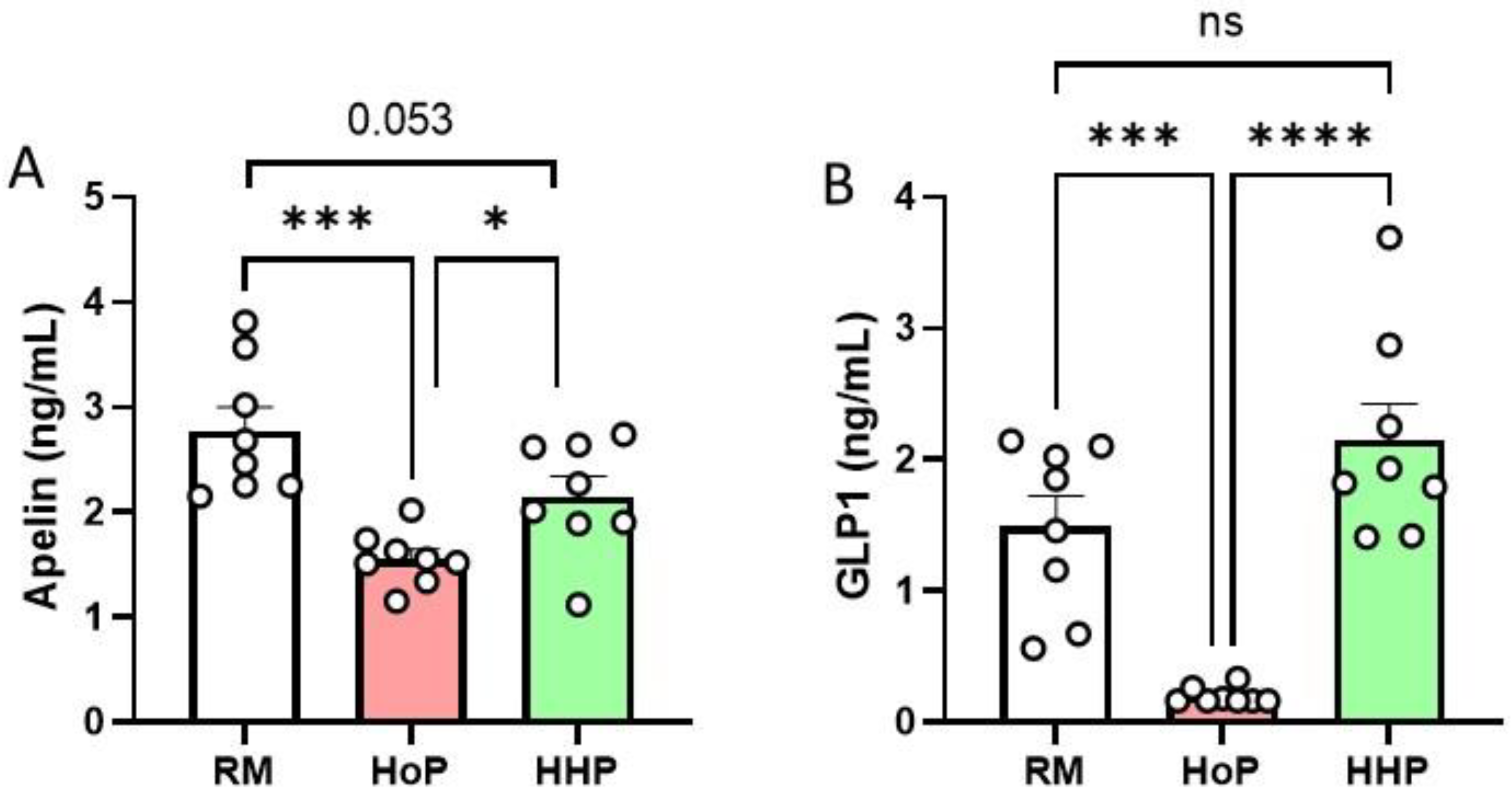
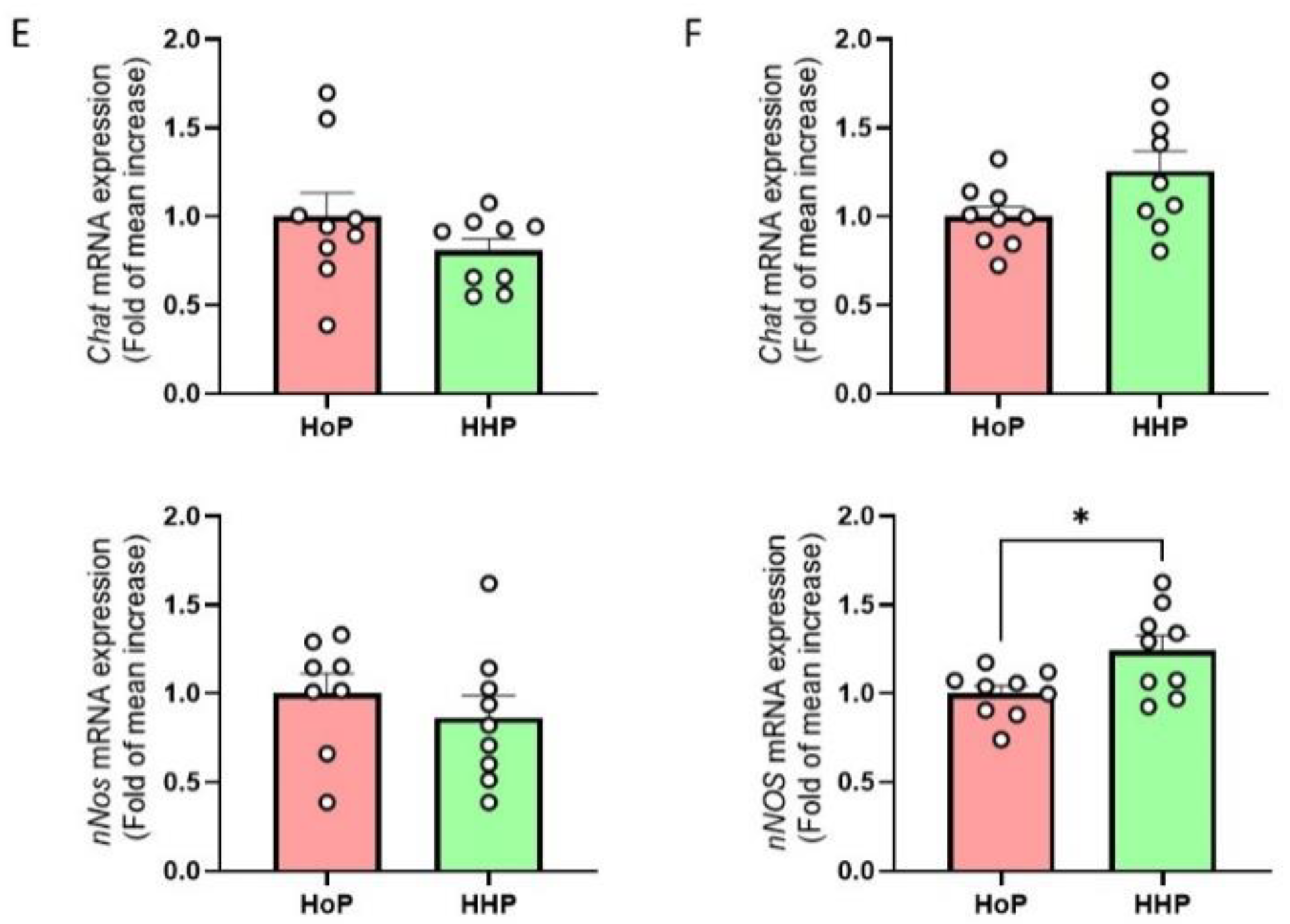
3.1. Apelin and GLP-1 Levels in RM-, HoP- and HHP-BM
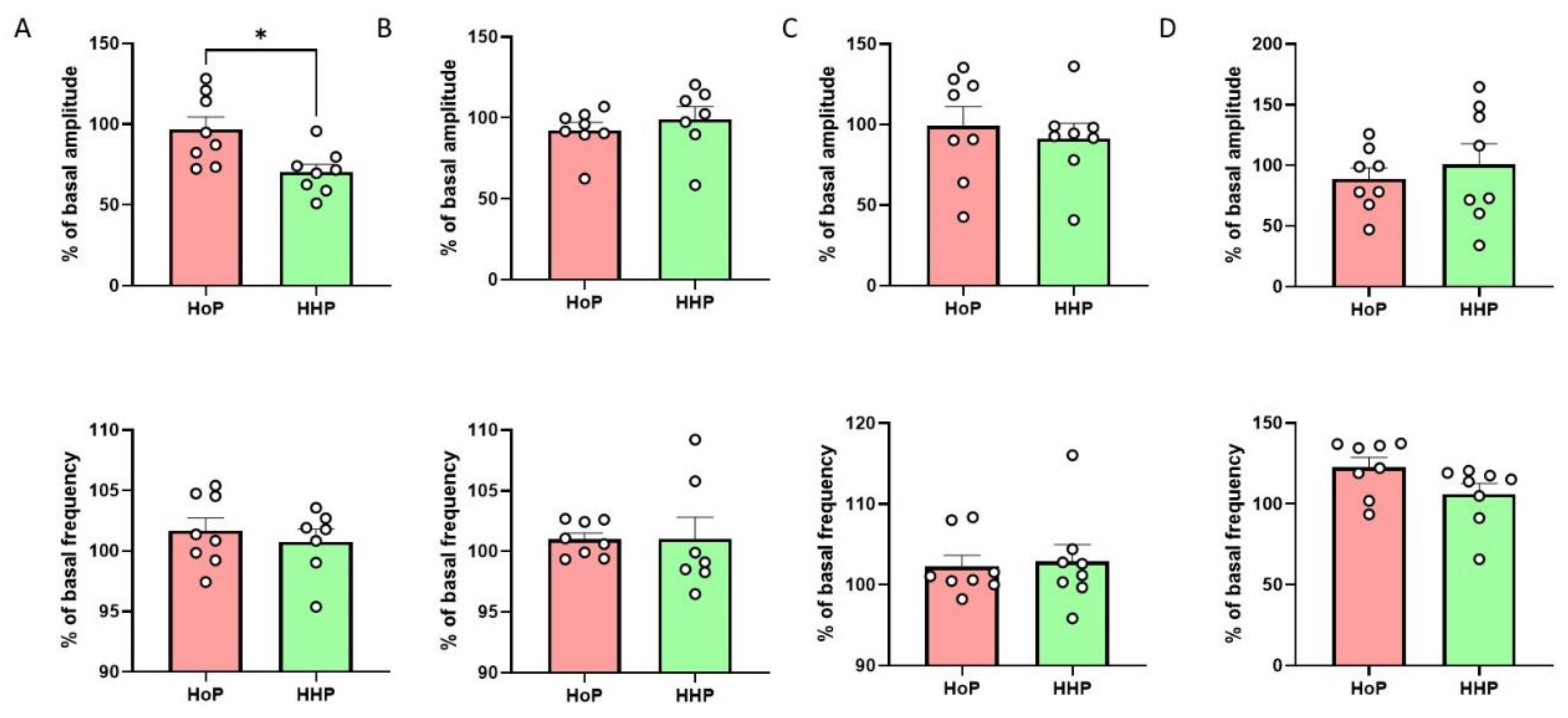
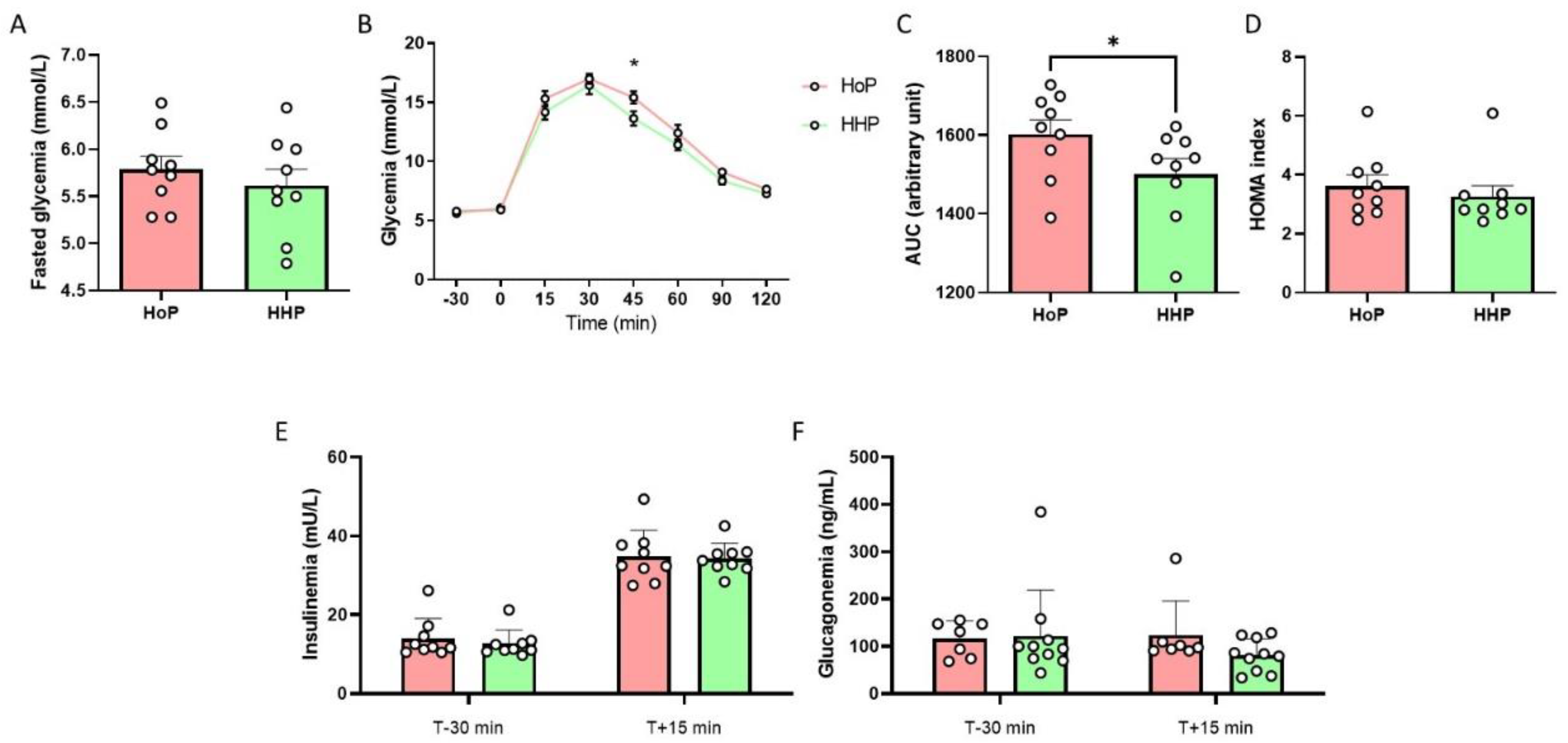
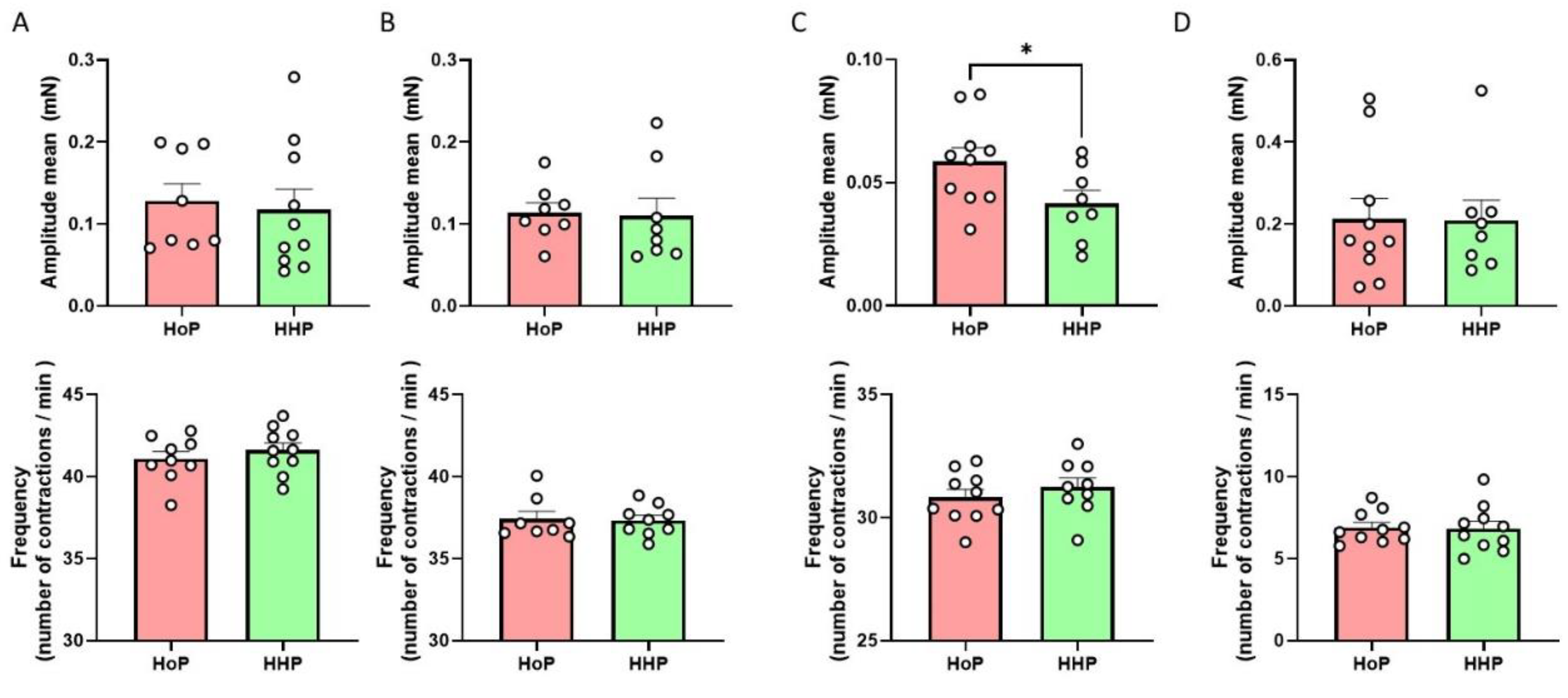
3.2. Impact of HoP- and HHP-BM on Intestinal Contractions and Glucose Metabolism in Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Duchon, J.; Barbian, M.E.; Denning, P.W. Necrotizing Enterocolitis. Clin. Perinatol. 2021, 48, 229–250. [Google Scholar] [CrossRef] [PubMed]
- Picaud, J.C.; Buffin, R. Human Milk-Treatment and Quality of Banked Human Milk. Clin. Perinatol. 2017, 44, 95–119. [Google Scholar] [CrossRef]
- Billeaud, C. High Hydrostatic Pressure Treatment Ensures the Microbiological Safety of Human Milk Including Bacillus cereus and Preservation of Bioactive Proteins Including Lipase and Immuno-Proteins: A Narrative Review. Foods 2021, 10, 1327. [Google Scholar] [CrossRef]
- Badillo-Suarez, P.A.; Rodriguez-Cruz, M.; Nieves-Morales, X. Impact of Metabolic Hormones Secreted in Human Breast Milk on Nutritional Programming in Childhood Obesity. J. Mammary Gland Biol. Neoplasia 2017, 22, 171–191. [Google Scholar] [CrossRef]
- Schueler, J.; Alexander, B.; Hart, A.M.; Austin, K.; Larson-Meyer, D.E. Presence and dynamics of leptin, GLP-1, and PYY in human breast milk at early postpartum. Obesity 2013, 21, 1451–1458. [Google Scholar] [CrossRef]
- Fournel, A.; Drougard, A.; Duparc, T.; Marlin, A.; Brierley, S.M.; Castro, J.; Le-Gonidec, S.; Masri, B.; Colom, A.; Lucas, A.; et al. Apelin targets gut contraction to control glucose metabolism via the brain. Gut 2017, 66, 258–269. [Google Scholar] [CrossRef] [Green Version]
- Knauf, C.; Abot, A.; Wemelle, E.; Cani, P.D. Targeting the Enteric Nervous System to Treat Metabolic Disorders? “Enterosynes” as Therapeutic Gut Factors. Neuroendocrinology 2020, 110, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Burcelin, R.; Cani, P.D.; Knauf, C. Glucagon-like peptide-1 and energy homeostasis. J. Nutr. 2007, 137, 2534S–2538S. [Google Scholar] [CrossRef] [Green Version]
- Burcelin, R.; Da Costa, A.; Drucker, D.; Thorens, B. Glucose competence of the hepatoportal vein sensor requires the presence of an activated glucagon-like peptide-1 receptor. Diabetes 2001, 50, 1720–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knauf, C.; Cani, P.D.; Perrin, C.; Iglesias, M.A.; Maury, J.F.; Bernard, E.; Benhamed, F.; Gremeaux, T.; Drucker, D.J.; Kahn, C.R.; et al. Brain glucagon-like peptide-1 increases insulin secretion and muscle insulin resistance to favor hepatic glycogen storage. J. Clin. Investig. 2005, 115, 3554–3563. [Google Scholar] [CrossRef] [PubMed]
- Knauf, C.; Cani, P.D.; Kim, D.H.; Iglesias, M.A.; Chabo, C.; Waget, A.; Colom, A.; Rastrelli, S.; Delzenne, N.M.; Drucker, D.J.; et al. Role of central nervous system glucagon-like Peptide-1 receptors in enteric glucose sensing. Diabetes 2008, 57, 2603–2612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demazeau, G.; Plumecocq, A.; Lehours, P.; Martin, P.; Couedelo, L.; Billeaud, C. A New High Hydrostatic Pressure Process to Assure the Microbial Safety of Human Milk While Preserving the Biological Activity of Its Main Components. Front. Public Health 2018, 6, 306. [Google Scholar] [CrossRef] [PubMed]
- Abot, A.; Wemelle, E.; Laurens, C.; Paquot, A.; Pomie, N.; Carper, D.; Bessac, A.; Mas Orea, X.; Fremez, C.; Fontanie, M.; et al. Identification of new enterosynes using prebiotics: Roles of bioactive lipids and mu-opioid receptor signalling in humans and mice. Gut 2021, 70, 1078–1087. [Google Scholar] [CrossRef]
- Wesolowska, A.; Sinkiewicz-Darol, E.; Barbarska, O.; Strom, K.; Rutkowska, M.; Karzel, K.; Rosiak, E.; Oledzka, G.; Orczyk-Pawilowicz, M.; Rzoska, S.; et al. New Achievements in High-Pressure Processing to Preserve Human Milk Bioactivity. Front. Pediatr. 2018, 6, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escuder-Vieco, D.; Espinosa-Martos, I.; Rodriguez, J.M.; Fernandez, L.; Pallas-Alonso, C.R. Effect of HTST and Holder Pasteurization on the Concentration of Immunoglobulins, Growth Factors, and Hormones in Donor Human Milk. Front. Immunol. 2018, 9, 2222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abot, A.; Lucas, A.; Bautzova, T.; Bessac, A.; Fournel, A.; Le-Gonidec, S.; Valet, P.; Moro, C.; Cani, P.D.; Knauf, C. Galanin enhances systemic glucose metabolism through enteric Nitric Oxide Synthase-expressed neurons. Mol. Metab. 2018, 10, 100–108. [Google Scholar] [CrossRef]
- Wen, J.; Phillips, S.F.; Sarr, M.G.; Kost, L.J.; Holst, J.J. PYY and GLP-1 contribute to feedback inhibition from the canine ileum and colon. Am. J. Physiol. 1995, 269, G945–G952. [Google Scholar] [CrossRef]
- Holst, J.J. The physiology of glucagon-like peptide 1. Physiol. Rev. 2007, 87, 1409–1439. [Google Scholar] [CrossRef] [PubMed]
- Borgmann, D.; Ciglieri, E.; Biglari, N.; Brandt, C.; Cremer, A.L.; Backes, H.; Tittgemeyer, M.; Wunderlich, F.T.; Bruning, J.C.; Fenselau, H. Gut-brain communication by distinct sensory neurons differently controls feeding and glucose metabolism. Cell Metab. 2021, 33, 1466–1482.e7. [Google Scholar] [CrossRef]
- Grasset, E.; Puel, A.; Charpentier, J.; Collet, X.; Christensen, J.E.; Terce, F.; Burcelin, R. A Specific Gut Microbiota Dysbiosis of Type 2 Diabetic Mice Induces GLP-1 Resistance through an Enteric NO-Dependent and Gut-Brain Axis Mechanism. Cell Metab. 2017, 25, 1075–1090.e5. [Google Scholar] [CrossRef] [Green Version]
- Charpentier, J.; Briand, F.; Lelouvier, B.; Servant, F.; Azalbert, V.; Puel, A.; Christensen, J.E.; Waget, A.; Branchereau, M.; Garret, C.; et al. Liraglutide targets the gut microbiota and the intestinal immune system to regulate insulin secretion. Acta Diabetol. 2021, 58, 881–897. [Google Scholar] [CrossRef] [PubMed]
- Amato, A.; Cinci, L.; Rotondo, A.; Serio, R.; Faussone-Pellegrini, M.S.; Vannucchi, M.G.; Mule, F. Peripheral motor action of glucagon-like peptide-1 through enteric neuronal receptors. Neurogastroenterol. Motil. Off. J. Eur. Gastrointest. Motil. Soc. 2010, 22, 664-e203. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Tang, Q.; Quan, X.; Ren, H.; Chen, W.; Xia, H.; Luo, H. Effects of exendin-4 on colonic motility in rats and its underlying mechanism. Neurogastroenterol. Motil. Off. J. Eur. Gastrointest. Motil. Soc. 2019, 31, e13482. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene of Interest | Forward Sequence | Reverse Sequence |
|---|---|---|
| Hprt | GTTCTTTGCTGACCTGCTGGAT | CCCCGTTGACTGATCATTACAG |
| nNos | ACGTCAAGTACGCCACCAACA | GCGAGTTCCACACTCGGAAGT |
| Chat | TGATCTTTGCTCGGCAGCACT | TTGGCCCAGTCAGTGGGAATG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wemelle, E.; Marousez, L.; de Lamballerie, M.; Knauf, C.; Lesage, J. High Hydrostatic Pressure Processing of Human Milk Increases Apelin and GLP-1 Contents to Modulate Gut Contraction and Glucose Metabolism in Mice Compared to Holder Pasteurization. Nutrients 2022, 14, 219. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14010219
Wemelle E, Marousez L, de Lamballerie M, Knauf C, Lesage J. High Hydrostatic Pressure Processing of Human Milk Increases Apelin and GLP-1 Contents to Modulate Gut Contraction and Glucose Metabolism in Mice Compared to Holder Pasteurization. Nutrients. 2022; 14(1):219. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14010219
Chicago/Turabian StyleWemelle, Eve, Lucie Marousez, Marie de Lamballerie, Claude Knauf, and Jean Lesage. 2022. "High Hydrostatic Pressure Processing of Human Milk Increases Apelin and GLP-1 Contents to Modulate Gut Contraction and Glucose Metabolism in Mice Compared to Holder Pasteurization" Nutrients 14, no. 1: 219. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14010219