
Chiliadenus iphionoides Reduces Body Weight and Improves Parameters Related to Hepatic Lipid and Glucose Metabolism in a High-Fat-Diet-Induced Mice Model of NAFLD

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chiliadenus Iphionoides Plant Extract Preparation
2.2. Experimental Animals and Diets
2.3. Oral Glucose Tolerance Test (OGTT)
2.4. Homa-IR
2.5. Animal Sacrifice and Organ Collection
2.6. Biochemical Analysis of Serum Parameters
2.7. Liver and Intestine Histology Examination
2.8. Western Blot Analysis
2.9. Quantitative Real-Time PCR
2.10. Statistical Analysis
3. Results
3.1. C. iphionoides Supplementation Reduced Body and Tissue Weight
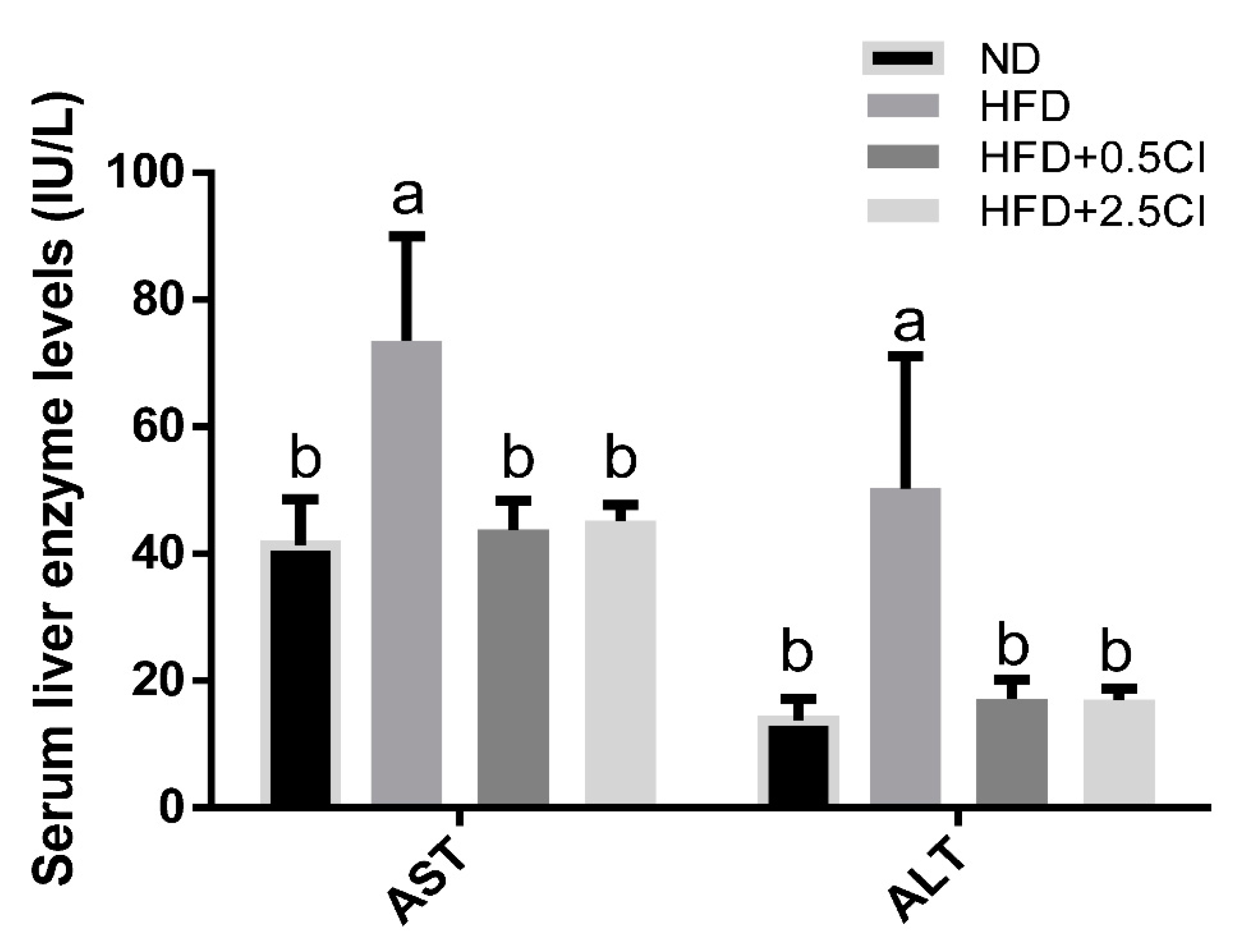
3.2. C. iphionoides Supplementation Improved Liver Enzymes and Serum Lipid Profile
3.3. Chiliadenus iphionoides Did Not Affect Glucose Tolerance
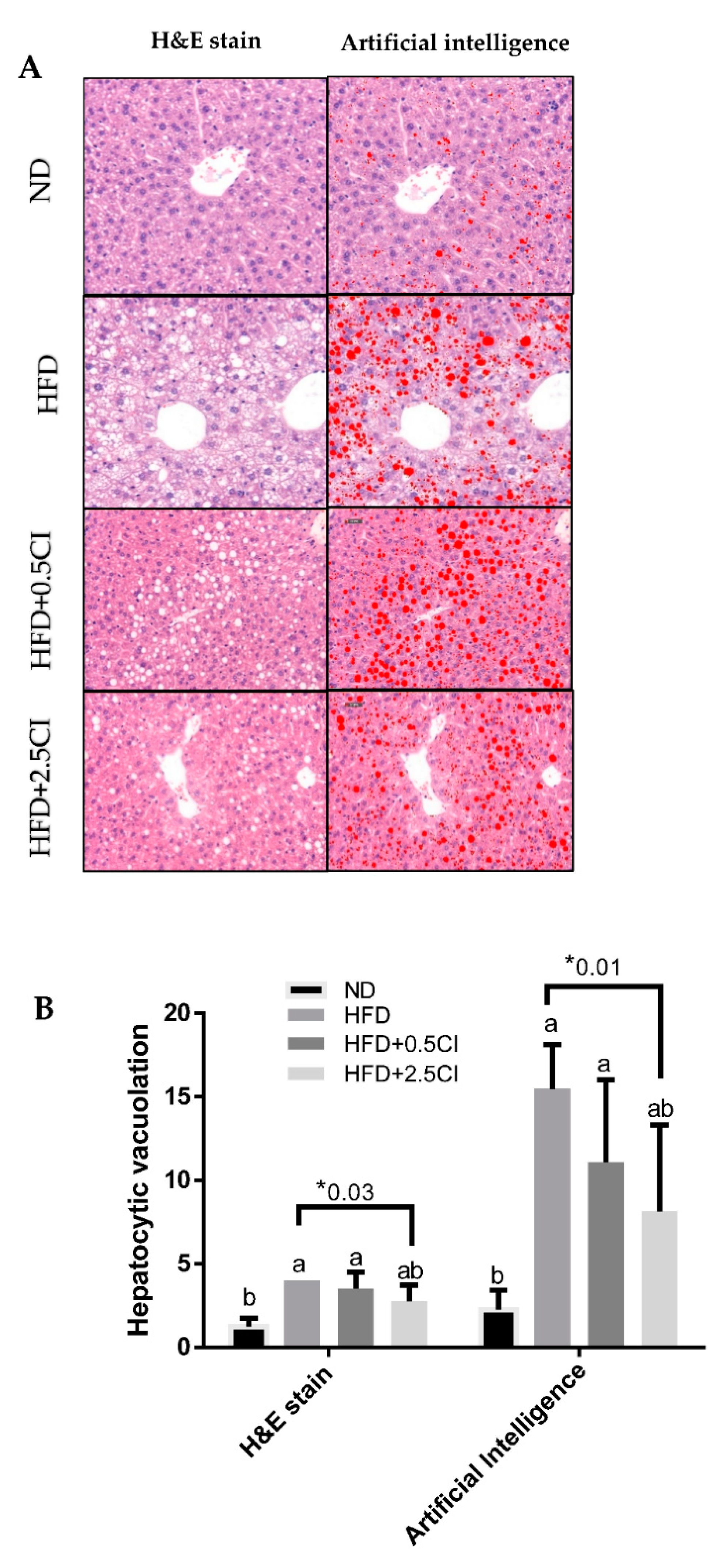
3.4. Chiliadenus iphionoides Consumption Alleviated Liver Lipid Steatosis
3.5. Chiliadenus iphionoides Impact on Genes Related to Lipid and Carbohydrate Metabolism and Inflammatory Markers
3.6. Chiliadenus iphionoides Supplementation Did Not Affect Protein Expression in Lipid and Carbohydrate Metabolism
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Moschen, A.R.; Kaser, S.; Tilg, H. Non-alcoholic steatohepatitis: A microbiota-driven disease. Trends Endocrinol. Metab. 2013, 24, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Cotter, T.G.; Rinella, M. NAFLD 2020: The State of the Disease. Gastroenterology 2020, 158, 1851–1864. [Google Scholar] [CrossRef] [PubMed]
- Lau, J.K.C.; Zhang, X.; Yu, J. Animal models of non-alcoholic fatty liver disease: Current perspectives and recent advances. J. Pathol. 2017, 241, 36–44. [Google Scholar] [CrossRef]
- Friedman, S.L.; Neuschwander-Tetri, B.A.; Rinella, M.; Sanyal, A.J. Mechanisms of NAFLD Development and Therapeutic Strategies. Nat. Med. 2018, 24, 908–922. [Google Scholar] [CrossRef] [PubMed]
- Araújo, A.R.; Rosso, N.; Bedogni, G.; Tiribelli, C.; Bellentani, S. Global epidemiology of non-alcoholic fatty liver disease/non-alcoholic steatohepatitis: What we need in the future. Liver Int. 2018, 38, 47–51. [Google Scholar] [CrossRef] [Green Version]
- Poojar, B.; Ommurugan, B.; Adiga, S.; Thomas, H.; Sori, R. Methodology Used in the Study. Asian J. Pharm. Clin. Res. 2017, 7, 1–5. [Google Scholar] [CrossRef]
- Bagherniya, M.; Nobili, V.; Blesso, C.N.; Sahebkar, A. Medicinal plants and bioactive natural compounds in the treatment of non-alcoholic fatty liver disease: A clinical review. Pharmacol. Res. 2018, 130, 213–240. [Google Scholar] [CrossRef]
- Bengtson, A.; Anderberg, A.A. Species diversification in the Mediterranean genus Chiliadenus (Inuleae-Asteraceae). Plant Syst. Evol. 2018, 304, 853–860. [Google Scholar] [CrossRef] [Green Version]
- Al-Bakheit, A.; Abu-Romman, S.; Sharab, A.; Shhab, M.A. Anti-inflammatory effect of Varthemia iphionoides extracts against prostate cancer in vitro. Eur. J. Inflamm. 2017, 15, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Abdelhalim, A.R. Pharmacological Properties and Chemical Constituents of Chiliadenus iphionoides (Syn. Varthemia iphionoides): A Review. Eur. J. Med. Plants. 2020, 31, 84–97. [Google Scholar] [CrossRef]
- Gorelick, J.; Kitron, A.; Pen, S.; Rosenzweig, T.; Madar, Z. Anti-diabetic activity of Chiliadenus iphionoides. J. Ethnopharmacol. 2011, 137, 1245–1249. [Google Scholar] [CrossRef] [PubMed]
- Abu-zaiton, A.; Alsohaili, S.; Abu-zaitoon, Y. Anti-hyperglycemic Effect and Liver Enzymes Activity of Varthemia iphionoides essential Oil in Diabetic Rats. Int. J. Adv. Biotechnol. Res. 2018, 9, 549–554. [Google Scholar]
- Heikkinen, S.; Argmann, C.A.; Champy, M.F.; Auwerx, J. Evaluation of Glucose Homeostasis. Curr. Protoc. Mol. Biol. 2007, 77, 1–22. [Google Scholar] [CrossRef]
- Ramot, Y.; Zandani, G.; Madar, Z.; Deshmukh, S.; Nyska, A. Utilization of a Deep Learning Algorithm for Microscope-Based Fatty Vacuole Quantification in a Fatty Liver Model in Mice. Toxicol. Pathol. 2020, 48, 702–707. [Google Scholar] [CrossRef]
- Schafer, K.A.; Eighmy, J.; Fikes, J.D.; Halpern, W.G.; Hukkanen, R.R.; Long, G.G.; Meseck, E.K.; Patrick, D.J.; Thibodeau, M.S.; Wood, C.E. Use of Severity Grades to Characterize Histopathologic Changes. Toxicol. Pathol. 2018, 46, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Zandani, G.; Anavi-cohen, S.; Tsybina-shimshilashvili, N.; Sela, N.; Nyska, A.; Madar, Z. Broccoli Florets Supplementation Improves Insulin Sensitivity and Alters Gut Microbiome Population—A Steatosis Mice Model Induced by High-Fat Diet. Front. Nutr. 2021, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Harlev, E.; Nevo, E.; Mirsky, N.; Ofir, R. Antidiabetic attributes of desert and steppic plants: A review. Planta Med. 2013, 79, 425–436. [Google Scholar] [CrossRef] [Green Version]
- Gruben, N.; Shiri-Sverdlov, R.; Koonen, D.P.Y.; Hofker, M.H. Nonalcoholic fatty liver disease: A main driver of insulin resistance or a dangerous liaison? Biochim. Biophys. Acta-Mol. Basis Dis. 2014, 1842, 2329–2343. [Google Scholar] [CrossRef] [Green Version]
- Hariri, N.; Thibault, L. High-fat diet-induced obesity in animal models. Nutr. Res. Rev. 2010, 23, 270–299. [Google Scholar] [CrossRef] [Green Version]
- Reaven, G.M. What do we learn from measurements of HOMA-IR? Diabetologia 2013, 56, 1867–1868. [Google Scholar] [CrossRef] [Green Version]
- O’Toole, A.; Moule, S.K.; Lockyer, P.J.; Halestrap, A.P. Tumour necrosis factor-α activation of protein kinase B in WEHI-164 cells is accompanied by increased phosphorylation of Ser473, but not Thr308. Biochem. J. 2001, 359, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Vadlakonda, L.; Dash, A.; Pasupuleti, M.; Kumar, K.A.; Reddanna, P. The paradox of Akt-mTOR interactions. Front. Oncol. 2013, 3, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen Shay, K.; Hagen, T.M. Age-associated impairment of Akt phosphorylation in primary rat hepatocytes is remediated by alpha-lipoic acid through PI3 kinase, PTEN, and PP2A. Biogerontology 2009, 10, 443–456. [Google Scholar] [CrossRef] [Green Version]
- Vergadi, E.; Ieronymaki, E.; Lyroni, K.; Vaporidi, K.; Tsatsanis, C. Akt Signaling Pathway in Macrophage Activation and M1/M2 Polarization. J. Immunol. 2017, 198, 1006–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frias, M.A.; Thoreen, C.C.; Jaffe, J.D.; Schroder, W.; Sculley, T.; Carr, S.A.; Sabatini, D.M. mSin1 Is Necessary for Akt/PKB Phosphorylation, and Its Isoforms Define Three Distinct mTORC2s. Curr. Biol. 2006, 16, 1865–1870. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wu, H.; Liu, Y.; Yang, L. High fat diet induced obesity model using four strains of mice: Kunming, c57bl/6, balb/c and icr. Exp. Anim. 2020, 69, 326–335. [Google Scholar] [CrossRef] [Green Version]
- März, W.; Kleber, M.E.; Scharnagl, H.; Speer, T.; Zewinger, S.; Ritsch, A.; Parhofer, K.G.; von Eckardstein, A.; Landmesser, U.; Laufs, U. HDL cholesterol: Reappraisal of its clinical relevance. Clin. Res. Cardiol. 2017, 106, 663–675. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Jay, A.; Brunaldi, K.; Huang, N.; Hamilton, J.A. CD36 enhances fatty acid uptake by increasing the rate of intracellular esterification but not transport across the plasma membrane. Biochemistry 2013, 52, 7254–7261. [Google Scholar] [CrossRef]
- Pawlak, M.; Lefebvre, P.; Staels, B. Molecular mechanism of PPARα action and its impact on lipid metabolism, inflammation and fibrosis in non-alcoholic fatty liver disease. J. Hepatol. 2015, 62, 720–733. [Google Scholar] [CrossRef] [Green Version]
- Förstermann, U.; Xia, N.; Li, H. Roles of Vascular Oxidative Stress and Nitric Oxide in the Pathogenesis of Atherosclerosis. Circ. Res. 2017, 120, 713–735. [Google Scholar] [CrossRef]
- Thomas, M.J.; Sorci-thomas, M.G. SAA: A link between cholesterol efflux capacity and inflammation? J. Lipid Res. 2015, 56, 1383–1385. [Google Scholar] [CrossRef] [PubMed]
- Al-Dabbas, M.M.; Kitahara, K.; Suganuma, T.; Hashimoto, F.; Tadera, K. Antioxidant and α-amylase inhibitory compounds from aerial parts of Varthemia iphionoides Boiss. Biosci. Biotechnol. Biochem. 2006, 70, 2178–2184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monraz-Méndez, C.A.; Escutia-Gutiérrez, R.; Rodriguez-Sanabria, J.S.; Galicia-Moreno, M.; Monroy-Ramírez, H.C.; Sánchez-Orozco, L.; García-Bañuelos, J.; De la Rosa-Bibiano, R.; Santos, A.; Armendáriz-Borunda, J.; et al. Moringa oleifera Improves MAFLD by Inducing Epigenetic Modifications. Nutrients 2022, 14, 4225. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normal Diet (ND) | High-Fat Diet (HFD) | |||
|---|---|---|---|---|
| Ingredients | gr | Kcal | gr | Kcal |
| Casein | 210 | 840 | 265 | 1060 |
| L- methionine | 3 | 12 | 4 | 16 |
| Cornstarch | 500 | 2000 | 0 | 0 |
| Dextrose | 100 | 400 | 160 | 640 |
| Sucrose | 39.15 | 156.6 | 90 | 360 |
| Lard | 20 | 180 | 310 | 2790 |
| Soybean oil | 20 | 180 | 30 | 270 |
| Anhydrous milkfat | 20 | 180 | 0 | 0 |
| Cellulose | 35 | 0 | 65.5 | 0 |
| Mineral mix AIN-93G-MX (94046) | 35 | 0 | 51.4 | 0 |
| Vitamin mix AIN-93-VX (94047). | 15 | 0 | 21 | 0 |
| Choline chloride | 2.75 | 0 | 3 | 0 |
| BHT | 0.014 | 0 | 0.014 | 0 |
| Total energy (Kacl) | 1000 | 3769 | 1000 | 5136 |
| Fat (%) | 9 | 60 | ||
| Protein (%) | 22 | 21 | ||
| Carbohydrate (%) | 65 | 19 | ||
| Name | Reverse | Forward |
|---|---|---|
| 18s | 5′-CCTCAGTTCCGAAAACCAAC-3′ | 5’-ACCGCAGCTAGGAATAATGG-3’ |
| iNOS | 5′-TCTCTGCTCTCAGCTCCAAG-3′ | 5′-AGCTCCCTCCTTCTCCTTCT-3′ |
| SAA-1 | 5′-GGTCAGCAATGGTGTCCTCA-3′ | 5′-GATGAAGCTACTCACCAGCCT-3′ |
| PPARα | 5′-CTGCGCATGCTCCGTG-3′ | 5′-CTTCCCAAAGCTCCTTCAAAAA- 3′ |
| CD36 | 5′-AAAGGCATTGGCTGGAAGAA-3′ | 5′-TCCTCTGACATTTGCAGGTCTATC-3′ |
| Fasn | 5′-GGTCGTTTCTCCATTAAATTCTCAT-3′ | 5′-CTAGAAACTTTCCCAGAAATCTTCC-3′ |
| Srebp-1c | 5′-TAGATGGTGGCTGCTGAGTG-3′ | 5′-GATCAAAGAGGAGCCAGTGC-3′ |
| CPT-1 | 5′-CAGCGAGTAGCGCATAGTCA-3′ | 5′-TGAGTGGCGTCCTCTTTGG-3′ |
| Group | ||||
|---|---|---|---|---|
| ND (n = 10) | HFD (n = 10) | HFD + 0.5CI (n = 10) | HFD + 2.5CI (n = 10) | |
| Initial body weight (g) | 22.06 ± 0.20 a | 21.76 ± 0.34 a | 21.17 ± 0.31 a | 21.39 ± 0.23 a |
| Final body weight (g) | 27.19 ± 0.41 c | 40.63 ± 1.23 a | 35.65 ± 1.06 b | 32.99 ± 1.13 b |
| Food intake (g/day) | 3.27 ± 0.48 a | 2.87 ± 0.45 b | 2.64 ± 0.34 b | 2.75 ± 0.31 b |
| Adipose tissue weight (g) | 0.60 ± 0.06 b | 1.8 ± 0.12 a | 1.59 ± 0.12 ab | 1.17 ± 0.16 b |
| Liver tissue weight (g) | 0.96 ± 0.03 b | 1.32 ± 0.07 a | 1.10 ± 0.05 b | 1.02 ± 0.05 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zandani, G.; Anavi-Cohen, S.; Yudelevich, T.; Nyska, A.; Dudai, N.; Madar, Z.; Gorelick, J. Chiliadenus iphionoides Reduces Body Weight and Improves Parameters Related to Hepatic Lipid and Glucose Metabolism in a High-Fat-Diet-Induced Mice Model of NAFLD. Nutrients 2022, 14, 4552. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14214552
Zandani G, Anavi-Cohen S, Yudelevich T, Nyska A, Dudai N, Madar Z, Gorelick J. Chiliadenus iphionoides Reduces Body Weight and Improves Parameters Related to Hepatic Lipid and Glucose Metabolism in a High-Fat-Diet-Induced Mice Model of NAFLD. Nutrients. 2022; 14(21):4552. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14214552
Chicago/Turabian StyleZandani, Gil, Sarit Anavi-Cohen, Tamar Yudelevich, Abraham Nyska, Nativ Dudai, Zecharia Madar, and Jonathan Gorelick. 2022. "Chiliadenus iphionoides Reduces Body Weight and Improves Parameters Related to Hepatic Lipid and Glucose Metabolism in a High-Fat-Diet-Induced Mice Model of NAFLD" Nutrients 14, no. 21: 4552. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14214552