
Beneficial In Vitro Effects of a Low Myo-Inositol Dose in the Regulation of Vascular Resistance and Protein Peroxidation under Inflammatory Conditions

 , , , , ,
, , , , ,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Drugs and Chemicals
2.2. Animal Studies
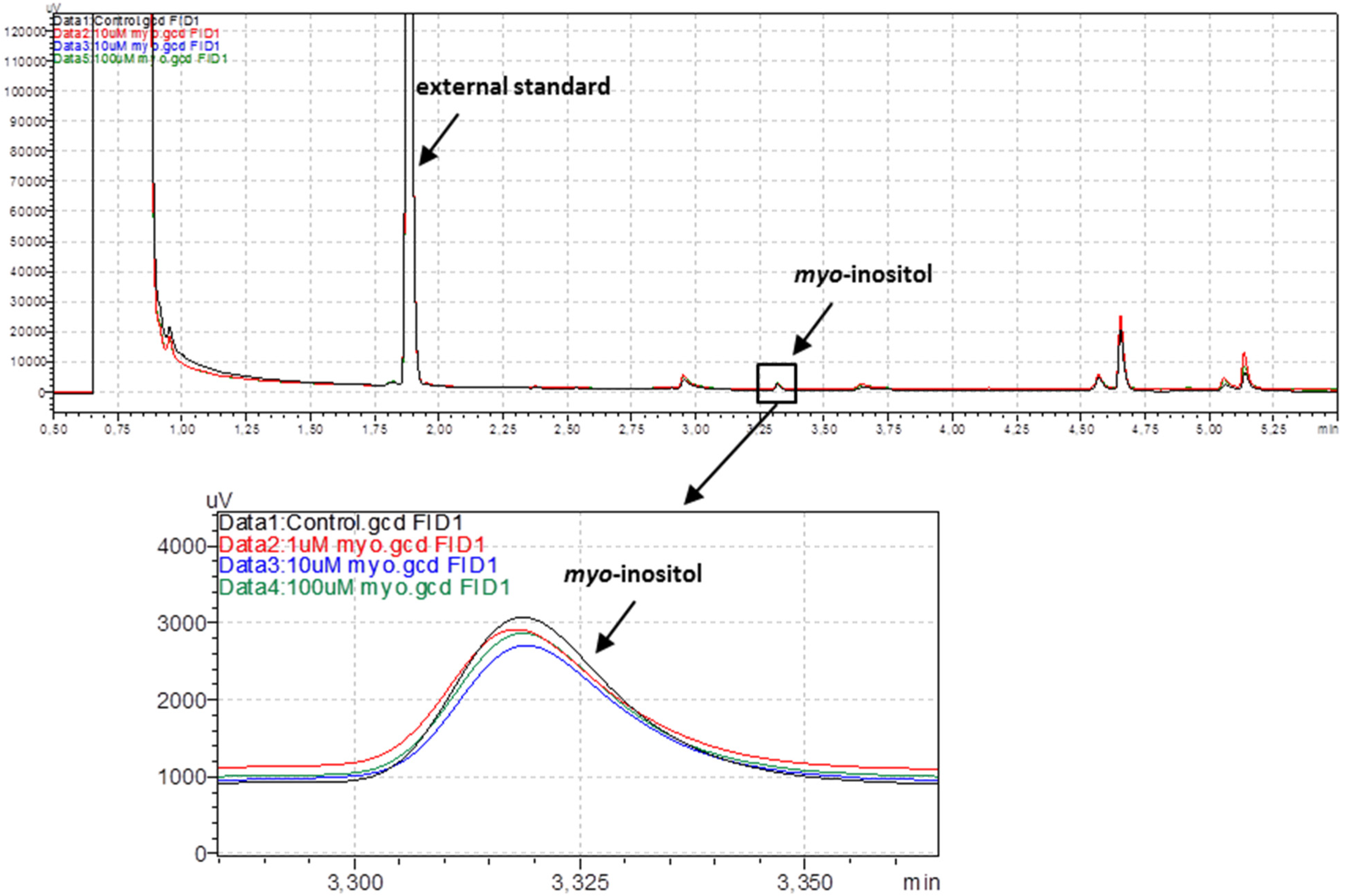
2.2.1. Gas Chromatography with Flame-Ionization Detection of Myo-Inositol
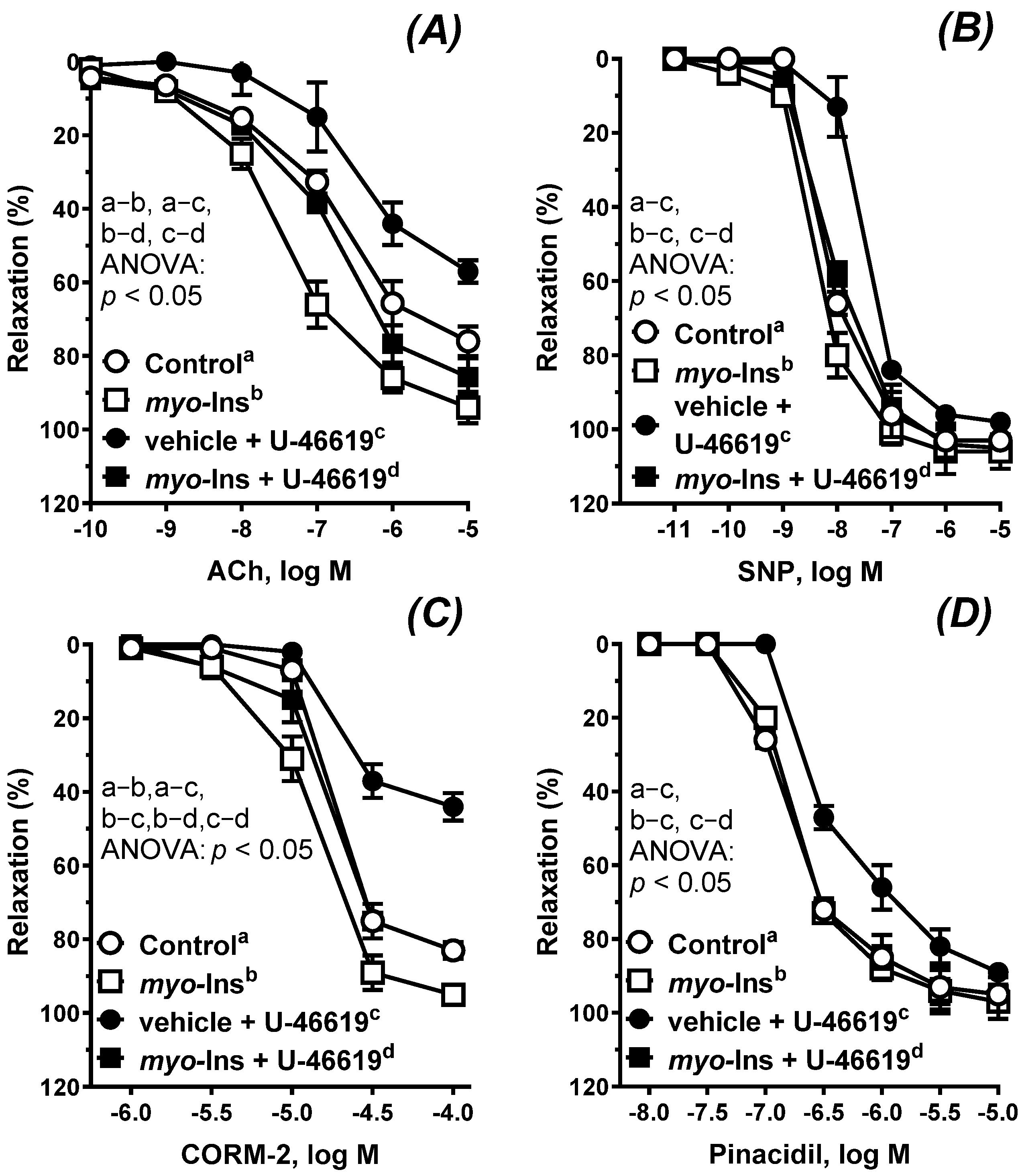
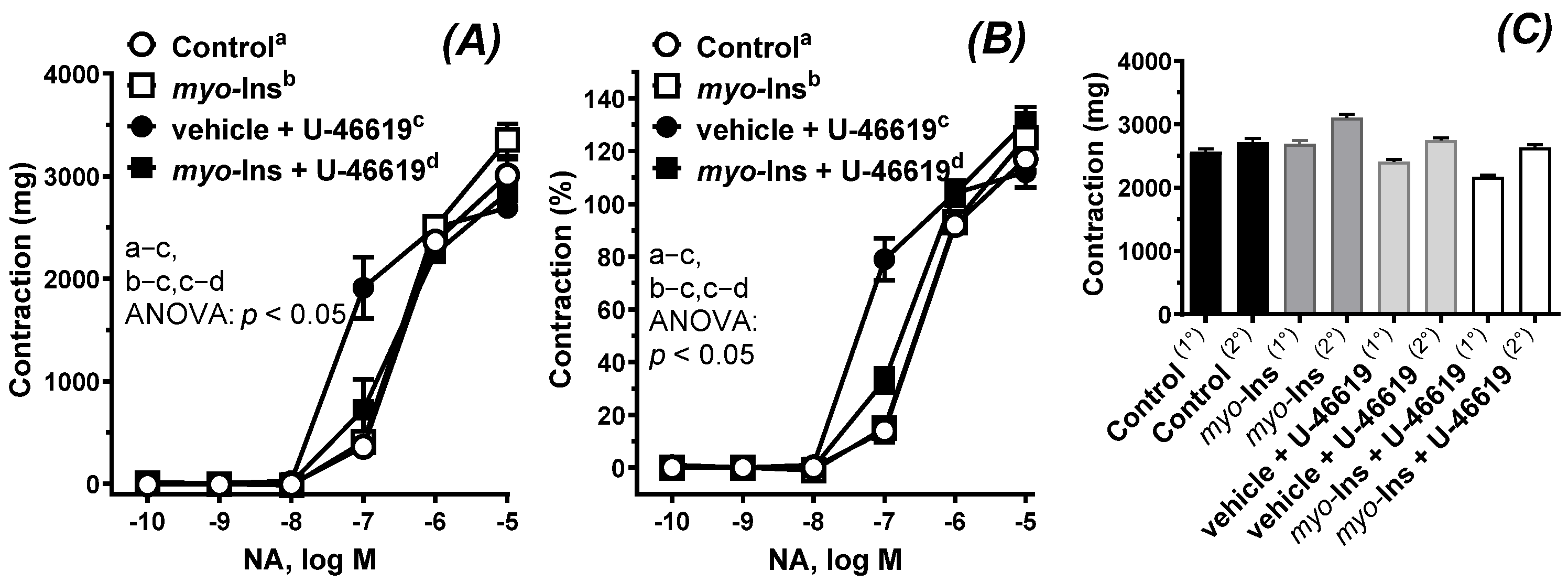
2.2.2. Vascular Reactivity Studies
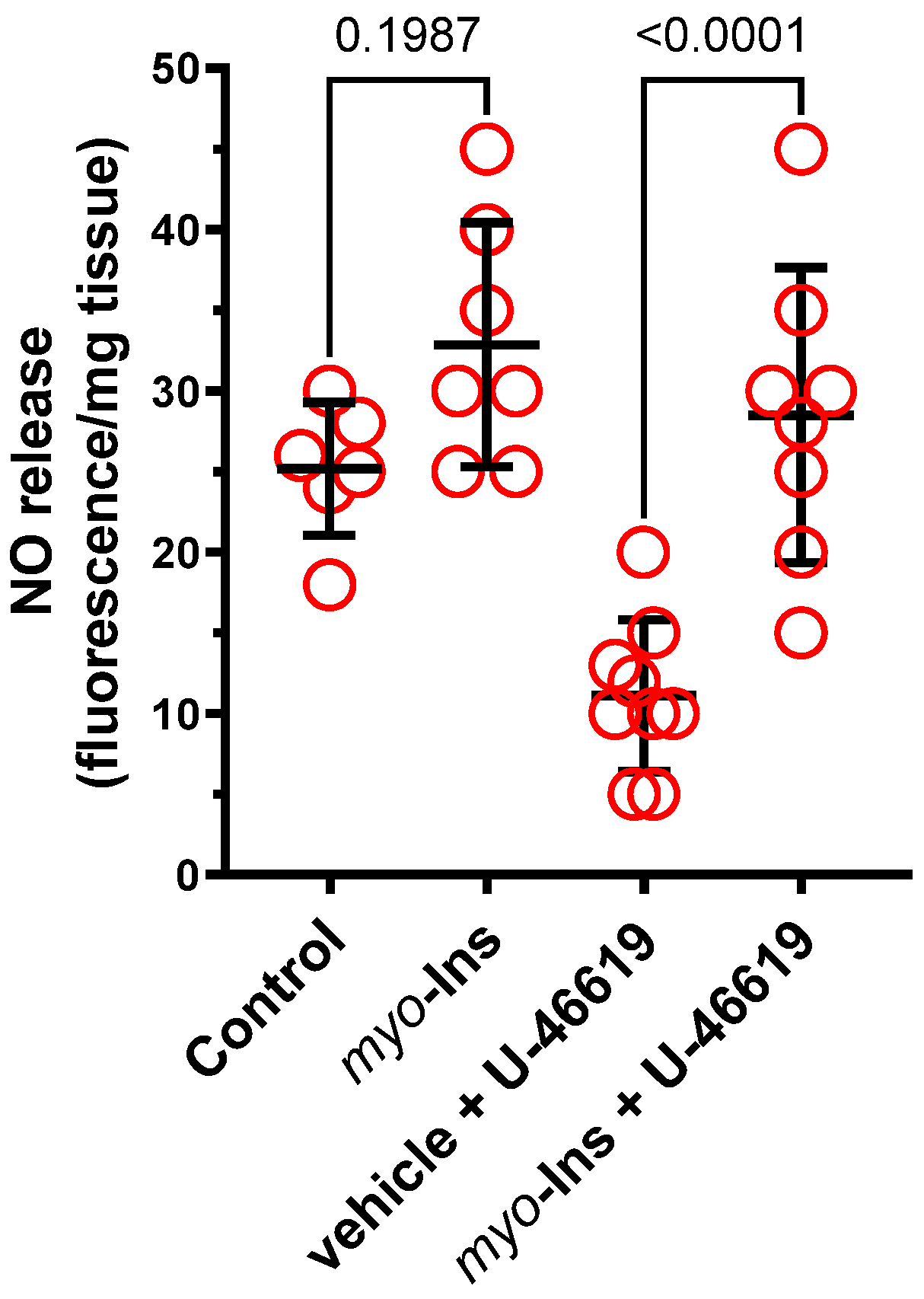
2.2.3. Nitric Oxide Release
2.2.4. Detection of Hydrogen Peroxide
2.3. In Vitro Studies on the Human Plasma
2.3.1. Markers of Oxidative Stress
2.3.2. Hemostasis Parameters
2.4. Data Analysis and Statistics
3. Results
3.1. Gas Chromatography with Flame-Ionization Detection of Myo-Inositol in Rat Arteries
3.2. Vascular Reactivity Studies
3.3. Nitric Oxide Release
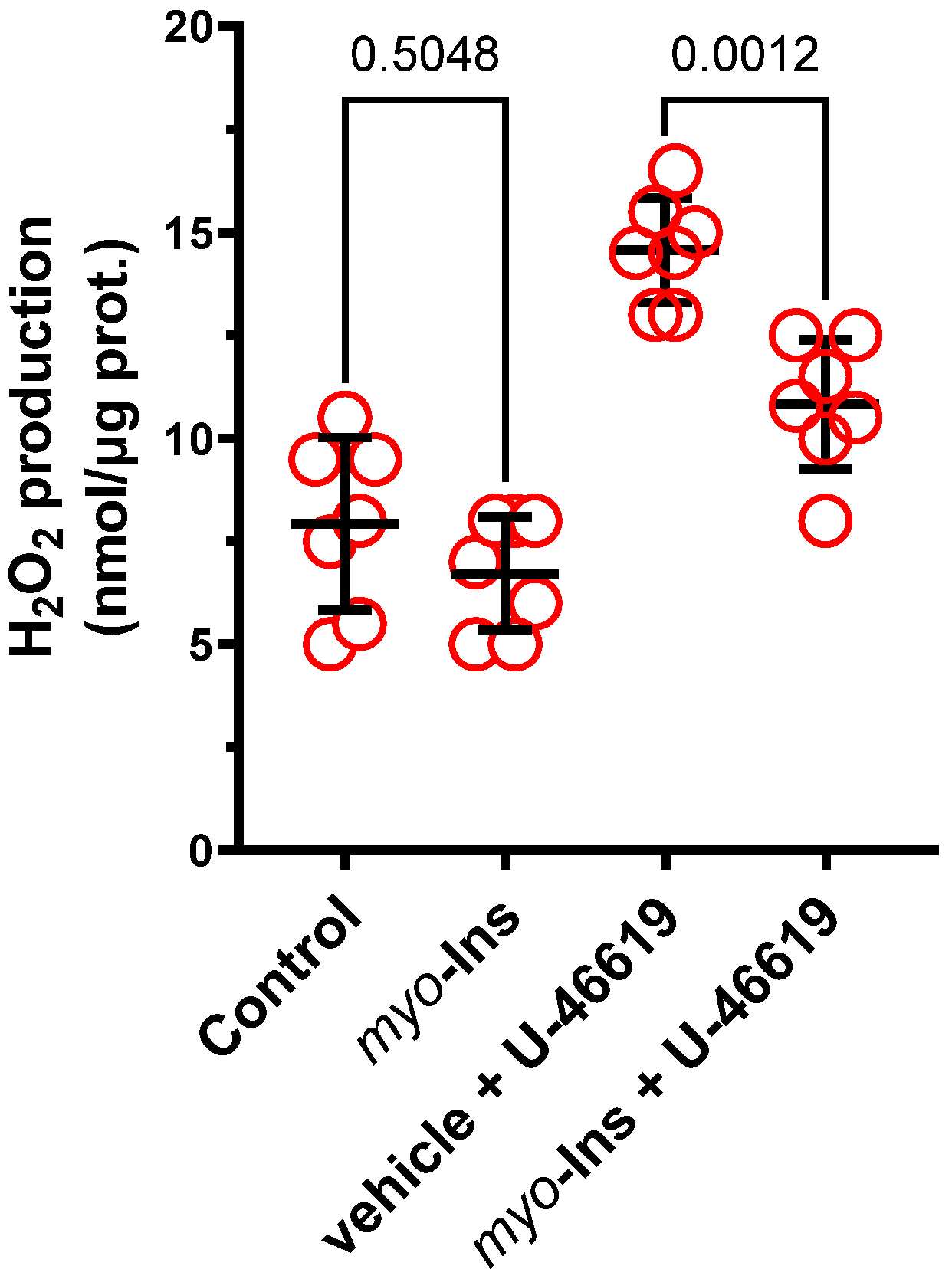
3.4. Detection of Hydrogen Peroxide
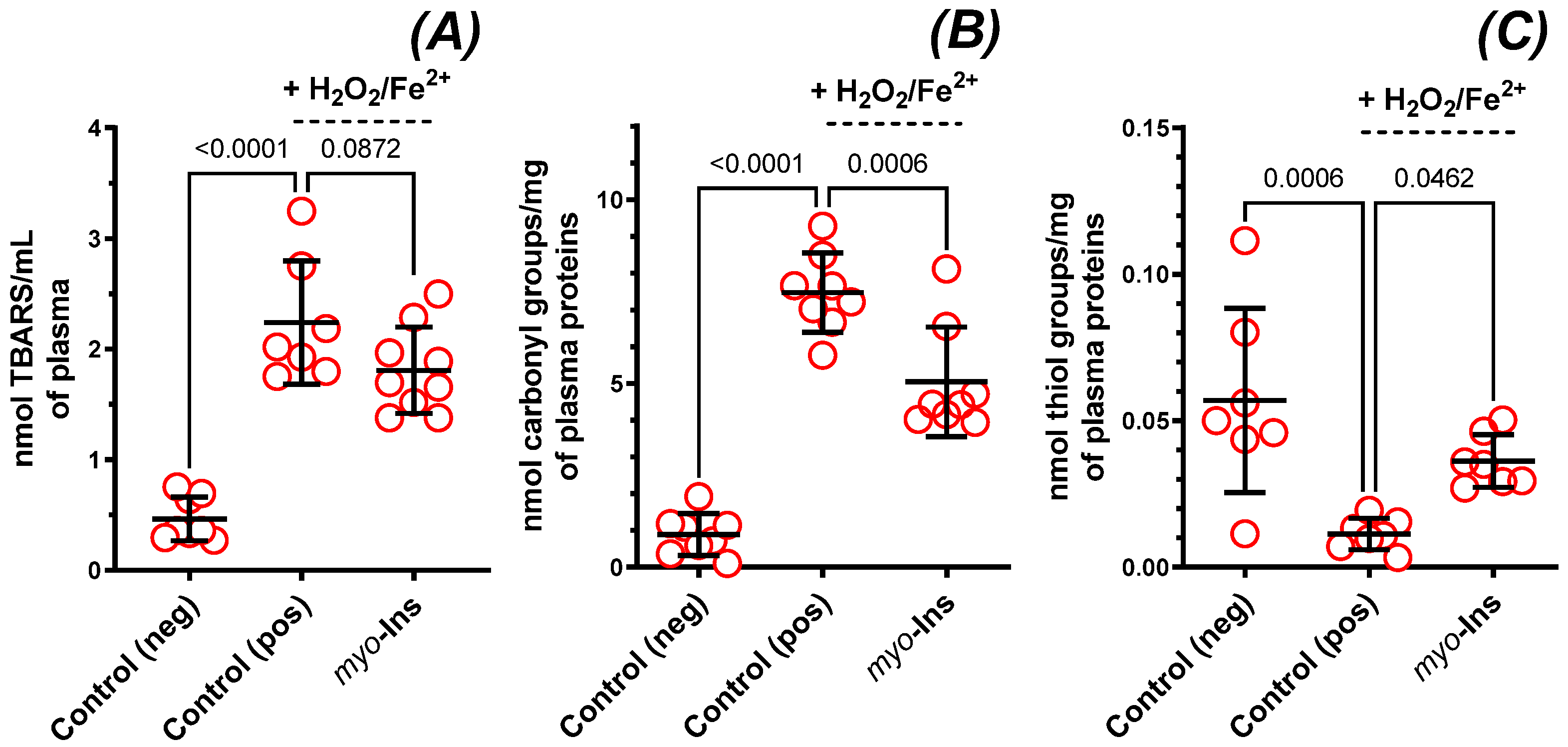
3.5. Markers of Oxidative Stress in Plasma
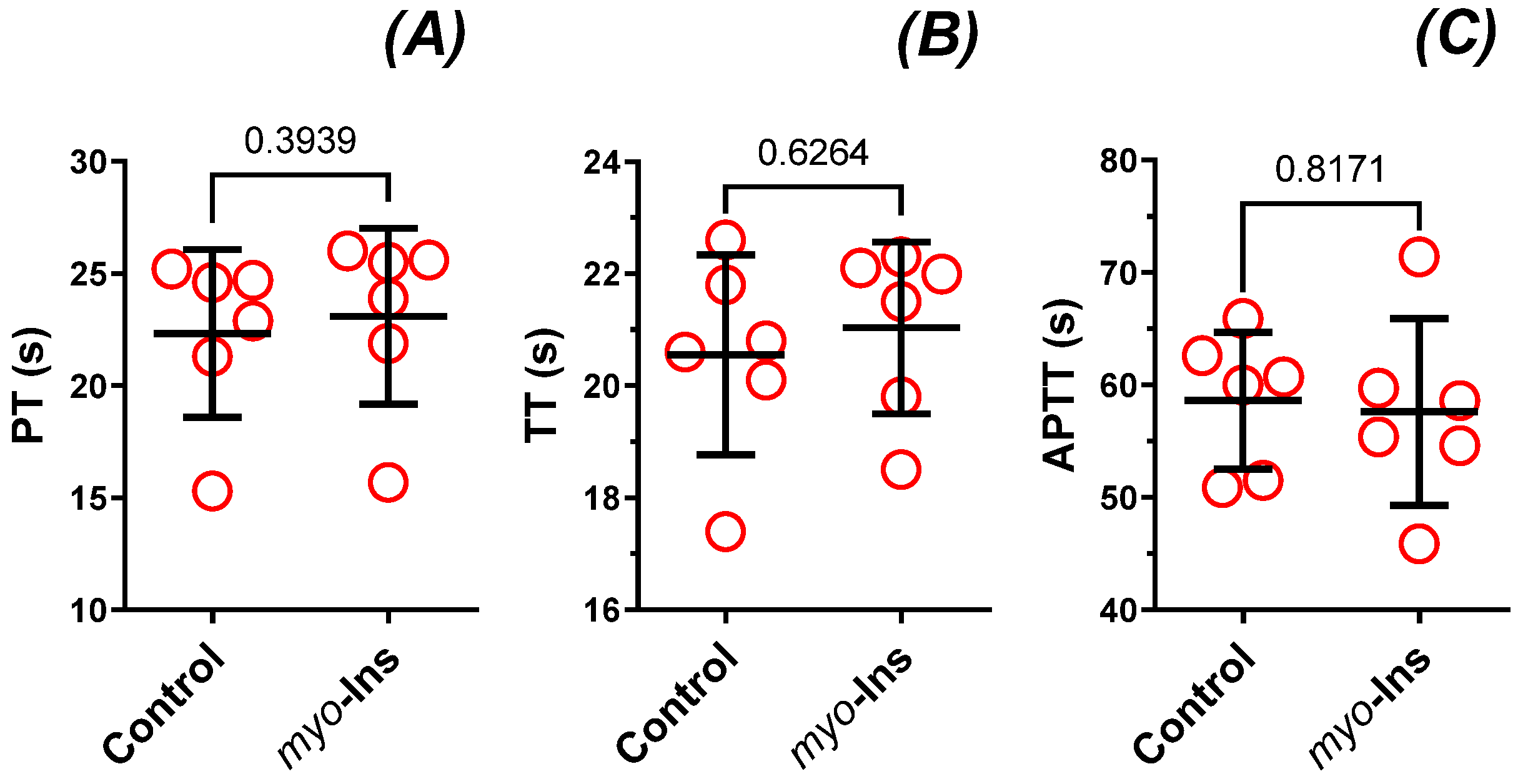
3.6. Parameters of Hemostasis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moreira, L.N.; Silva, J.F.; Silva, G.C.; Lemos, V.S.; Cortes, S.F. Activation of eNOS by D-pinitol Induces an Endothelium-Dependent Vasodilatation in Mouse Mesenteric Artery. Front. Pharmacol. 2018, 9, 528. [Google Scholar] [CrossRef] [PubMed]
- Antonowski, T.; Osowski, A.; Lahuta, L.; Górecki, R.; Rynkiewicz, A.; Wojtkiewicz, J. Health-Promoting Properties of Selected Cyclitols for Metabolic Syndrome and Diabetes. Nutrients 2019, 11, 2314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bizzarri, M.; Fuso, A.; Dinicola, S.; Cucina, A.; Bevilacqua, A. Pharmacodynamics and pharmacokinetics of inositol(s) in health and disease. Expert Opin. Drug Metab. Toxicol. 2016, 12, 1181–1196. [Google Scholar] [CrossRef] [PubMed]
- Baldassarre, M.P.A.; Di Tomo, P.; Centorame, G.; Pandolfi, A.; Di Pietro, N.; Consoli, A.; Formoso, G. Myoinositol reduces inflammation and oxidative stress in human endothelial cells exposed in vivo to chronic hyperglycemia. Nutrients 2021, 13, 2210. [Google Scholar] [CrossRef]
- Owczarczyk-Saczonek, A.; Lahuta, L.B.; Ligor, M.; Placek, W.; Górecki, R.J.; Buszewski, B. The Healing-Promoting Properties of Selected Cyclitols-A Review. Nutrients 2018, 10, 1891. [Google Scholar] [CrossRef] [Green Version]
- D’Anna, R.; Di Benedetto, A.; Scilipoti, A.; Santamaria, A.; Interdonato, M.L.; Petrella, E.; Neri, I.; Pintaudi, B.; Corrado, F.; Facchinetti, F. Myo-inositol Supplementation for Prevention of Gestational Diabetes in Obese Pregnant Women: A Randomized Controlled Trial. Obstet. Gynecol. 2015, 126, 310–315. [Google Scholar] [CrossRef] [Green Version]
- Pintaudi, B.; Di Vieste, G.; Bonomo, M. The Effectiveness of Myo-Inositol and D-Chiro Inositol Treatment in Type 2 Diabetes. Int. J. Endocrinol. 2016, 2016, 9132052. [Google Scholar] [CrossRef] [Green Version]
- Dell’Edera, D.; Sarlo, F.; Allegretti, A.; Simone, F.; Lupo, M.G.; Epifania, A.A. The influence of D-chiro-inositol and D-myo-inositol in pregnant women with glucose intolerance. Biomed. Rep. 2017, 7, 169–172. [Google Scholar] [CrossRef]
- Unfer, V.; Facchinetti, F.; Orrù, B.; Giordani, B.; Nestler, J. Myo-inositol effects in women with PCOS: A meta-analysis of randomized controlled trials. Endocr. Connect. 2017, 6, 647–658. [Google Scholar] [CrossRef] [Green Version]
- Santamaria, A.; Alibrandi, A.; Di Benedetto, A.; Pintaudi, B.; Corrado, F.; Facchinetti, F.; D’Anna, R. Clinical and metabolic outcomes in pregnant women at risk for gestational diabetes mellitus supplemented with myo-inositol: A secondary analysis from 3 RCTs. Am. J. Obstet. Gynecol. 2018, 219, 300.e1–300.e6. [Google Scholar] [CrossRef]
- Pallio, G.; Micali, A.; Benvenga, S.; Antonelli, A.; Marini, H.R.; Puzzolo, D.; Macaione, V.; Trichilo, V.; Santoro, G.; Irrera, N.; et al. Myo-inositol in the protection from cadmium-induced toxicity in mice kidney: An emerging nutraceutical challenge. Food Chem. Toxicol. 2019, 132, 110675. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, N.R.; Lessa, L.M.; Kerntopf, M.R.; Sousa, C.M.; Alves, R.S.; Queiroz, M.G.; Price, J.; Heimark, D.B.; Larner, J.; Du, X.; et al. Inositols prevent and reverse endothelial dysfunction in diabetic rat and rabbit vasculature metabolically and by scavenging superoxide. Proc. Natl. Acad. Sci. USA 2006, 103, 218–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, H.K. A 2020 vision for healthy people. N. Engl. J. Med. 2010, 362, 1653–1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majewski, M.; Ognik, K.; Juśkiewicz, J. The antioxidant status, lipid profile, and modulation of vascular function by fish oil supplementation in nano-copper and copper carbonate fed Wistar rats. J. Funct. Foods 2020, 64, 103595. [Google Scholar] [CrossRef]
- Majewski, M.; Jurgoński, A. The Effect of Hemp (Cannabis sativa L.) Seeds and Hemp Seed Oil on Vascular Dysfunction in Obese Male Zucker Rats. Nutrients 2021, 13, 2575. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Chino, D.; Kobayashi, T.; Obara, K.; Miyauchi, S.; Tanaka, Y. Selective and potent inhibitory effect of docosahexaenoic acid (DHA) on U46619-induced contraction in rat aorta. J. Smooth Muscle Res. 2013, 49, 63–77. [Google Scholar] [CrossRef] [Green Version]
- Vila, E.; Macrae, I.M.; Reid, J.L. Differences in inositol phosphate production in blood vessels of normotensive and spontaneously hypertensive rats. Br. J. Pharmacol. 1991, 104, 296–300. [Google Scholar] [CrossRef] [Green Version]
- Nestler, J.E.; Jakubowicz, D.J.; Reamer, P.; Gunn, R.D.; Allan, G. Ovulatory and metabolic effects of D-chiro-inositol in the polycystic ovary syndrome. N. Engl. J. Med. 1999, 340, 1314–1320. [Google Scholar] [CrossRef] [Green Version]
- Fruzzetti, F.; Perini, D.; Russo, M.; Bucci, F.; Gadducci, A. Comparison of two insulin sensitizers, metformin and myo-inositol, in women with polycystic ovary syndrome (PCOS). Gynecol. Endocrinol. 2017, 33, 39–42. [Google Scholar] [CrossRef]
- Levine, R.L.; Garland, D.; Oliver, C.N.; Amici, A.; Climent, I.; Lenz, A.G.; Ahn, B.W.; Shaltiel, S.; Stadtman, E.R. Determination of carbonyl content in oxidatively modified proteins. Methods Enzymol. 1990, 186, 464–478. [Google Scholar] [CrossRef]
- Ando, Y.; Steiner, M. Sulfhydryl and disulfide groups of platelet membranes. I. Determination of sulfhydryl groups. Biochim. Biophys. Acta 1973, 311, 26–37. [Google Scholar] [CrossRef]
- Malinowska, J.; Kołodziejczyk-Czepas, J.; Moniuszko-Szajwaj, B.; Kowalska, I.; Oleszek, W.; Stochmal, A.; Olas, B. Phenolic fractions from Trifolium pallidum and Trifolium scabrum aerial parts in human plasma protect against changes induced by hyperhomocysteinemia in vitro. Food Chem. Toxicol. 2012, 50, 4023–4027. [Google Scholar] [CrossRef] [PubMed]
- Rucker, D.; Dhamoon, A.S. Physiology, Thromboxane A2. 14 September 2021. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Álvarez-Maestro, M.; Eguibar, A.; Chanca, P.; Klett-Mingo, M.; Rivas, J.G.; Buño-Soto, A.; de Bethencourt, F.R.; Ferrer, M. Androgen Deprivation Therapy in Patients With Prostate Cancer Increases Serum Levels of Thromboxane A2: Cardiovascular Implications. Front. Cardiovasc. Med. 2021, 8, 653126. [Google Scholar] [CrossRef] [PubMed]
- Majewski, M.; Klett-Mingo, M.; Verdasco-Martín, C.M.; Otero, C.; Ferrer, M. Spirulina extract improves age-induced vascular dysfunction. Pharm. Biol. 2022, 2047209. [Google Scholar] [CrossRef]
- Arshad, M.; Vijay, V.; Floyd, B.C.; Marks, B.; Sarabu, M.R.; Wolin, M.S.; Gupte, S.A. Thromboxane receptor stimulation suppresses guanylate cyclase-mediated relaxation of radial arteries. Ann. Thorac. Surg. 2006, 81, 2147–2154. [Google Scholar] [CrossRef]
- Lacolley, P.; Regnault, V.; Segers, P.; Laurent, S. Vascular Smooth Muscle Cells and Arterial Stiffening: Relevance in Development, Aging, and Disease. Physiol. Rev. 2017, 97, 1555–1617. [Google Scholar] [CrossRef]
- Adach, W.; Żuchowski, J.; Moniuszko-Szajwaj, B.; Szumacher-Strabel, M.; Stochmal, A.; Olas, B.; Cieslak, A. Comparative Phytochemical, Antioxidant, and Hemostatic Studies of Extract and Four Fractions from Paulownia Clone in Vitro 112 Leaves in Human Plasma. Molecules 2020, 25, 4371. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Control (Vehicle) | Myo-Inositol (1 µM) | Vehicle + U-46619 (0.1 nM) | Myo-Inositol + U-46619 | ||||
|---|---|---|---|---|---|---|---|---|
| Emax (%) | LogEC50 | Emax (%) | LogEC50 | Emax (%) | LogEC50 | Emax (%) | LogEC50 | |
| ACh | 76.36 ac | −6.785 a | 91.40 a | −7.347 a | 58.47 bc | −6.498 b | 86.69 b | −6.834 b |
| ±SEM | 2.023 | 0.070 | 2.384 | 0.056 | 0.807 | 0.032 | 1.879 | 0.058 |
| SNP | 103.7 | −8.186 c | 106.9 | −8.363 | 101.1 | −7.478 bc | 104.3 | −8.076 b |
| ±SEM | 3.835 | 0.114 | 3.4 | 0.108 | 4.861 | 0.137 | 1.646 | 0.088 |
| CORM-2 | 83.08 a | −4.733 | 95.61 a | −4.878 | 44.08 bc | −4.676 | 83.29 b | −4.779 |
| ±SEM | 0.040 | 0.138 | 2.800 | 0.030 | 0.014 | 0.078 | 4.240 | 0.057 |
| Pinacidil | 97.72 | −6.977 c | 100.7 | −6.906 | 92.49 | −6.489 bc | 97.67 | −6.988 b |
| ±SEM | 3.162 | 0.106 | 4.876 | 0.149 | 6.711 | 0.165 | 3.608 | 0.130 |
| NA | 125.1 | −6.359 c | 133.2 | −6.314 | 112.9 b | −7.264 bc | 135.1 b | −6.525 b |
| ±SEM | 6.744 | 0.113 | 5.493 | 0.084 | 7.268 | 0.171 | 2.074 | 0.035 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rolnik, A.; Olas, B.; Szablińska-Piernik, J.; Lahuta, L.B.; Rynkiewicz, A.; Cygański, P.; Socha, K.; Gromadziński, L.; Thoene, M.; Majewski, M. Beneficial In Vitro Effects of a Low Myo-Inositol Dose in the Regulation of Vascular Resistance and Protein Peroxidation under Inflammatory Conditions. Nutrients 2022, 14, 1118. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14051118
Rolnik A, Olas B, Szablińska-Piernik J, Lahuta LB, Rynkiewicz A, Cygański P, Socha K, Gromadziński L, Thoene M, Majewski M. Beneficial In Vitro Effects of a Low Myo-Inositol Dose in the Regulation of Vascular Resistance and Protein Peroxidation under Inflammatory Conditions. Nutrients. 2022; 14(5):1118. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14051118
Chicago/Turabian StyleRolnik, Agata, Beata Olas, Joanna Szablińska-Piernik, Lesław Bernard Lahuta, Andrzej Rynkiewicz, Piotr Cygański, Katarzyna Socha, Leszek Gromadziński, Michael Thoene, and Michał Majewski. 2022. "Beneficial In Vitro Effects of a Low Myo-Inositol Dose in the Regulation of Vascular Resistance and Protein Peroxidation under Inflammatory Conditions" Nutrients 14, no. 5: 1118. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14051118