
Short-Term Intake of Theobroma grandiflorum Juice Fermented with Lacticaseibacillus rhamnosus ATCC 9595 Amended the Outcome of Endotoxemia Induced by Lipopolysaccharide
 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Materials
2.1.1. Probiotic Strain
2.1.2. Pulp Characterization
2.2. Fermentation
2.3. Optimization of Cultivation Conditions
2.3.1. Quantification of Lactic Acid Content
2.3.2. Chemical Characterization of Ethyl Phases from Unfermented and Fermented Juices
2.4. Animal Experimentation
2.4.1. Animals
2.4.2. Short-Term Administration of L. rhamnosus-Fermented and Unfermented Cupuaçu Juice
2.4.3. Induction and Evaluation of LPS-Mediated Endotoxemia
2.4.4. Determination of Cell Population in the Blood and Peritoneal Cavity
2.5. Statistical Analysis
3. Results
3.1. Nutritional Composition of Cupuaçu Pulp
3.2. Growth and Production of Lactic Acid by Lacticaseibacillus rhamnosus in Cupuaçu Juice
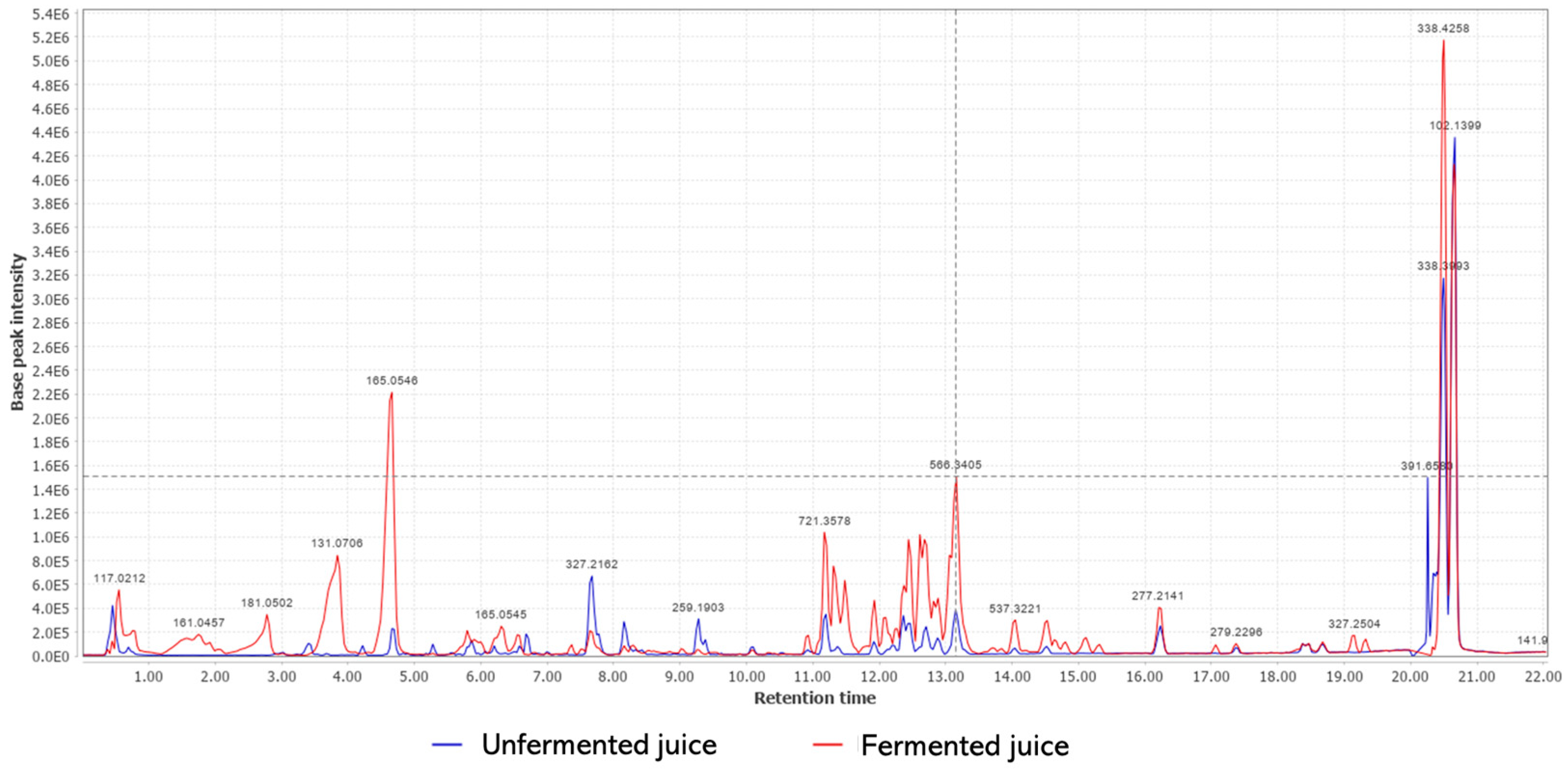
3.3. Chemical Composition
3.4. The Administration of L. rhamnosus-Fermented Juice Reduced the Severity of LPS-Mediated Endotoxemia
3.5. The Administration of L. rhamnosus-Fermented Juice Reduced the Weight Decrease Associated with Endotoxemia Induced by LPS
3.6. The Administration of L. rhamnosus-Fermented Juice Amended the Weight Reduction of Some Organs Associated with Endotoxemia Induced by LPS
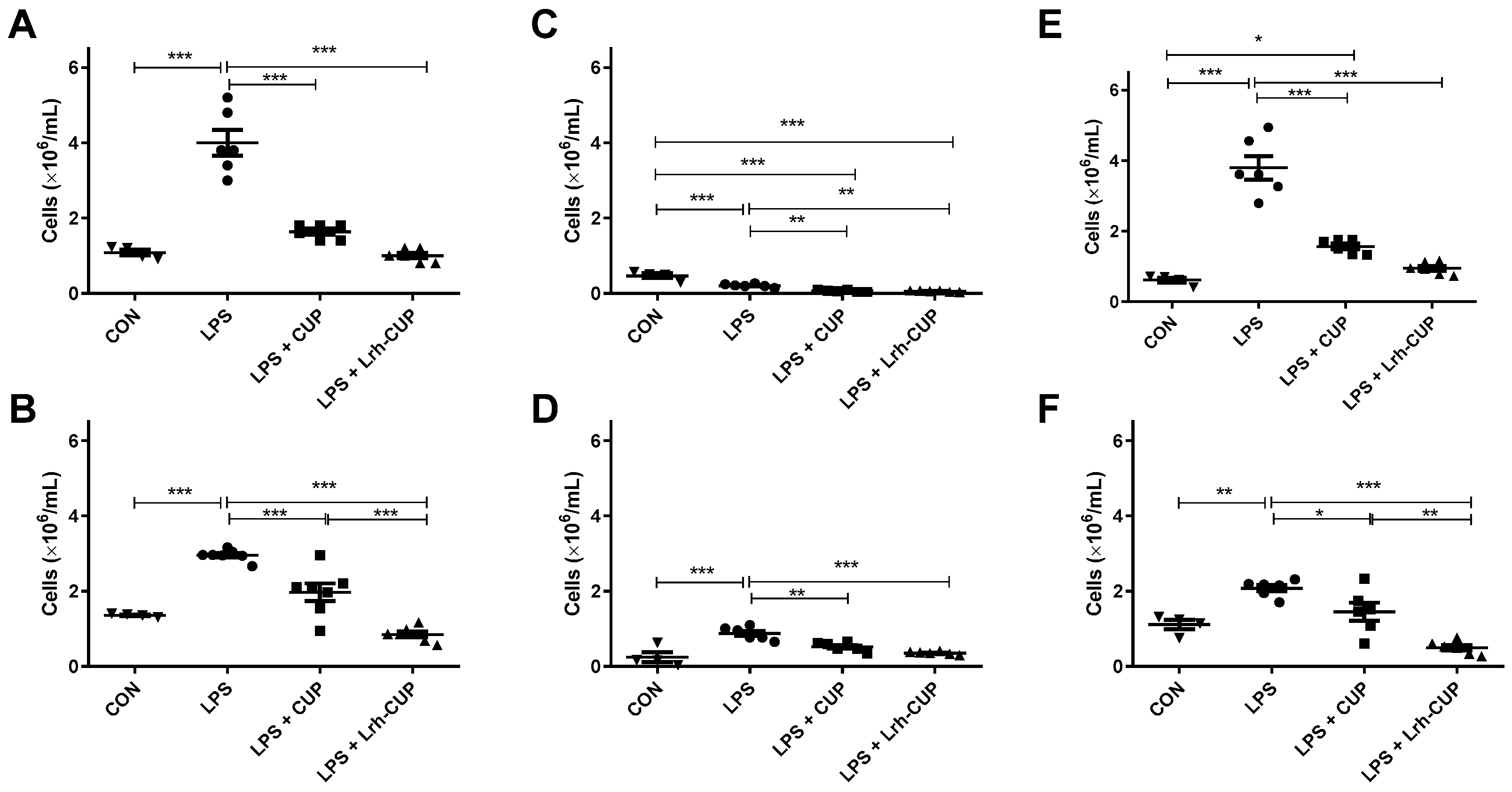
3.7. The Administration of L. rhamnosus-Fermented Juice Amended the Migration of Cells to the Peritoneal Cavity in Mice Submitted to Endotoxemia
3.8. The Administration of L. rhamnosus-Fermented Juice Reduced the Increase of Cells in the Blood Associated with LPS-Mediated Endotoxemia
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kamisoglu, K.; Haimovich, B.; Calvano, S.E.; Coyle, S.M.; Corbett, S.A.; Langley, R.J.; Kingsmore, S.F.; Androulakis, I.P. Human Metabolic Response to Systemic Inflammation: Assessment of the Concordance between Experimental Endotoxemia and Clinical Cases of Sepsis/SIRS. Crit. Care 2015, 19, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cochet, F.; Peri, F. The Role of Carbohydrates in the Lipopolysaccharide (LPS)/Toll-Like Receptor 4 (TLR4) Signalling. Int. J. Mol. Sci. 2017, 18, 2318. [Google Scholar] [CrossRef] [Green Version]
- Bertani, B.; Ruiz, N. Function and Biogenesis of Lipopolysaccharides. EcoSal Plus 2018, 8, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Fuke, N.; Nagata, N.; Suganuma, H.; Ota, T. Regulation of Gut Microbiota and Metabolic Endotoxemia with Dietary Factors. Nutrients 2019, 11, 2277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moludi, J.; Maleki, V.; Jafari-Vayghyan, H.; Vaghef-Mehrabany, E.; Alizadeh, M. Metabolic Endotoxemia and Cardiovascular Disease: A Systematic Review about Potential Roles of Prebiotics and Probiotics. Clin. Exp. Pharmacol. Physiol. 2020, 47, 927–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satokari, R. High Intake of Sugar and the Balance between Pro- and Anti-Inflammatory Gut Bacteria. Nutrients 2020, 12, 1348. [Google Scholar] [CrossRef]
- Koutroulis, I.; Batabyal, R.; McNamara, B.; Ledda, M.; Hoptay, C.; Freishtat, R.J. Sepsis Immunometabolism: From Defining Sepsis to Understanding How Energy Production Affects Immune Response. Crit Care Explor. 2019, 1, e0061. [Google Scholar] [CrossRef]
- Cavaillon, J.; Singer, M.; Skirecki, T. Sepsis Therapies: Learning from 30 Years of Failure of Translational Research to Propose New Leads. EMBO Mol. Med. 2020, 12, e10128. [Google Scholar] [CrossRef]
- Abdo, Z.; LeCureux, J.; LaVoy, A.; Eklund, B.; Ryan, E.P.; Dean, G.A. Impact of Oral Probiotic Lactobacillus Acidophilus Vaccine Strains on the Immune Response and Gut Microbiome of Mice. PloS ONE 2019, 14, e0225842. [Google Scholar] [CrossRef] [Green Version]
- Sireswar, S.; Biswas, S.; Dey, G. Adhesion and Anti-Inflammatory Potential of Lactobacillus Rhamnosus GG in a Sea Buckthorn Based Beverage Matrix. Food Funct. 2020, 11, 2555–2572. [Google Scholar] [CrossRef]
- Ávila, P.R.M.; Michels, M.; Vuolo, F.; Bilésimo, R.; Burger, H.; Milioli, M.V.M.; Sonai, B.; Borges, H.; Carneiro, C.; Abatti, M.; et al. Protective Effects of Fecal Microbiota Transplantation in Sepsis Are Independent of the Modulation of the Intestinal Flora. Nutrition 2020, 73, 110727. [Google Scholar] [CrossRef]
- Haro, C.; Mónaco, M.E.; Medina, M. Lactobacillus Casei Beneficially Modulates Immuno-Coagulative Response in an Endotoxemia Model. Blood Coagul. Fibrinolysis 2018, 29, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Nation, M.L.; Dunne, E.M.; Joseph, S.J.; Mensah, F.K.; Sung, V.; Satzke, C.; Tang, M.L.K. Impact of Lactobacillus Reuteri Colonization on Gut Microbiota, Inflammation, and Crying Time in Infant Colic. Sci. Rep. 2017, 7, 15047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, J.; Qi, S.; Cui, Y.; Dou, X.; Luo, X.M.; Liu, J.; Zhu, T.; Ma, Y.; Wang, H. Lactobacillus Rhamnosus GG Attenuates Lipopolysaccharide-Induced Inflammation and Barrier Dysfunction by Regulating MAPK/NF-ΚB Signaling and Modulating Metabolome in the Piglet Intestine. J. Nutr. 2020, 150, 1313–1323. [Google Scholar] [CrossRef]
- Mendes, Y.C.; Mesquita, G.P.; Costa, G.D.E.; Barbosa da Silva, A.C.; Gouveia, E.; Silva, M.R.C.; Monteiro-Neto, V.; de Miranda, R.d.C.M.; Nascimento da Silva, L.C.; Zagmignan, A. Evaluation of Growth, Viability, Lactic Acid Production and Anti-Infective Effects of Lacticaseibacillus Rhamnosus ATCC 9595 in Bacuri Juice (Platonia Insignis). Foods 2021, 10, 603. [Google Scholar] [CrossRef] [PubMed]
- de Sousa, M.A.; Rama, G.R.; Volken de Souza, C.F.; Granada, C.E. Acid Lactic Lactobacilli as a Biotechnological Toll to Improve Food Quality and Human Health. Biotechnol. Prog. 2020, 36, e2937. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Choi, K.B.; Park, J.H.; Kim, K.H. Metabolite Profile Changes and Increased Antioxidative and Antiinflammatory Activities of Mixed Vegetables after Fermentation by Lactobacillus Plantarum. PLoS ONE 2019, 14, e0217180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Tao, Y.; Li, D.; Wen, G.; Zhou, J.; Manickam, S.; Han, Y.; Chai, W.S. Fermentation of Blueberry Juices Using Autochthonous Lactic Acid Bacteria Isolated from Fruit Environment: Fermentation Characteristics and Evolution of Phenolic Profiles. Chemosphere 2021, 276, 130090. [Google Scholar] [CrossRef]
- Li, T.; Jiang, T.; Liu, N.; Wu, C.; Xu, H.; Lei, H. Biotransformation of Phenolic Profiles and Improvement of Antioxidant Capacities in Jujube Juice by Select Lactic Acid Bacteria. Food Chem. 2021, 339, 127859. [Google Scholar] [CrossRef]
- Lu, Y.; Tan, C.W.; Chen, D.; Liu, S.Q. Potential of Three Probiotic Lactobacilli in Transforming Star Fruit Juice into Functional Beverages. Food Sci. Nutr. 2018, 6, 2141–2150. [Google Scholar] [CrossRef]
- Kwaw, E.; Ma, Y.; Tchabo, W.; Apaliya, M.T.; Wu, M.; Sackey, A.S.; Xiao, L.; Tahir, H.E. Effect of Lactobacillus Strains on Phenolic Profile, Color Attributes and Antioxidant Activities of Lactic-Acid-Fermented Mulberry Juice. Food Chem. 2018, 250, 148–154. [Google Scholar] [CrossRef]
- Pugliese, A.G.; Tomas-Barberan, F.A.; Truchado, P.; Genovese, M.I. Flavonoids, Proanthocyanidins, Vitamin C, and Antioxidant Activity of Theobroma Grandiflorum (Cupuassu) Pulp and Seeds. J. Agric. Food Chem. 2013, 61, 2720–2728. [Google Scholar] [CrossRef]
- Barros, H.R.d.M.; García-Villalba, R.; Tomás-Barberán, F.A.; Genovese, M.I. Evaluation of the Distribution and Metabolism of Polyphenols Derived from Cupuassu (Theobroma Grandiflorum) in Mice Gastrointestinal Tract by UPLC-ESI-QTOF. J. Funct. Foods 2016, 22, 477–489. [Google Scholar] [CrossRef]
- Helrich, K. Official Methods of Analysis of the Association of Official Analytical Chemists; Association of Official Analytical Chemists: Arlington, VA, USA, 1990. [Google Scholar]
- Zenebon, O.; Pascuet, N.S.; Tiglea, P. Métodos Físico-Químicos Para Análise de Alimentos, 4th ed.; Instituto Adolfo Lutz: São Paulo, Brazil, 2008; 1020p. [Google Scholar]
- Farias, N.; Soares, M.; Gouveia, E. Enhancement of the Viability of Lactobacillus Rhamnosus ATCC 7469 in Passion Fruit Juice: Application of a Central Composite Rotatable Design. LWT 2016, 71, 149–154. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, F.; Wang, Z.; Li, Z.; Li, J. PNU-282987 Attenuates Intestinal Epithelial Barrier Dysfunction in LPS-Induced Endotoxemia. Inflammation 2020, 43, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Mendes, S.J.F.; Sousa, F.I.A.B.; Pereira, D.M.S.; Ferro, T.A.F.; Pereira, I.C.P.; Silva, B.L.R.; Pinheiro, A.J.M.C.R.; Mouchrek, A.Q.S.; Monteiro-Neto, V.; Costa, S.K.P.; et al. Cinnamaldehyde Modulates LPS-Induced Systemic Inflammatory Response Syndrome through TRPA1-Dependent and Independent Mechanisms. Int. Immunopharmacol. 2016, 34, 60–70. [Google Scholar] [CrossRef]
- Mukaka, M. A Guide to Appropriate Use of Correlation Coefficient in Medical Research. Malawi Med. J. 2012, 24, 69–71. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, R.N.; Grimbs, S.; Behrends, B.; Bernaert, H.; Ullrich, M.S.; Kuhnert, N. Origin-Based Polyphenolic Fingerprinting of Theobroma Cacao in Unfermented and Fermented Beans. Food Res. Int. 2017, 99, 550–559. [Google Scholar] [CrossRef]
- Duarte, W.F.; Dias, D.R.; Oliveira, J.M.; Teixeira, J.A.; de Almeida e Silva, J.B.; Schwan, R.F. Characterization of Different Fruit Wines Made from Cacao, Cupuassu, Gabiroba, Jaboticaba and Umbu. LWT-Food Sci. Technol. 2010, 43, 1564–1572. [Google Scholar] [CrossRef]
- Pereira-Caro, G.; Borges, G.; Nagai, C.; Jackson, M.C.; Yokota, T.; Crozier, A.; Ashihara, H. Profiles of Phenolic Compounds and Purine Alkaloids during the Development of Seeds of Theobroma Cacao Cv. Trinitario. J. Agric. Food Chem. 2013, 61, 427–434. [Google Scholar] [CrossRef]
- Porter, L.J.; Ma, Z.; Chan, B.G. Cacao Procyanidins: Major Flavanoids and Identification of Some Minor Metabolites. Phytochemistry 1991, 30, 1657–1663. [Google Scholar] [CrossRef]
- Neiverth, A.; Prim, L.R.; Franck, C.L.; Nisihara, R. Sepsis in Burned Adult Patients: Study of Serie of Cases in Brazil. J. Burn Care Res. 2020, 41, 900–904. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, N.R.C.; Pontes, G.F.; Jacob, F.L.; Deprá, J.V.S.; Porto, J.P.P.; de Lima, F.R.; de Albuquerque, M.R.T.C. Analysis of Trends in Sepsis Mortality in Brazil and by Regions from 2010 to 2019. Rev. Saude Publica 2022, 56, 25. [Google Scholar] [CrossRef]
- van der Poll, T.; van de Veerdonk, F.L.; Scicluna, B.P.; Netea, M.G. The Immunopathology of Sepsis and Potential Therapeutic Targets. Nat. Rev. Immunol. 2017, 17, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, H.; Chen, W.; Zhong, Q.; Zhang, G.; Chen, W. Beneficial Effects of Tomato Juice Fermented by Lactobacillus Plantarum and Lactobacillus Casei: Antioxidation, Antimicrobial Effect, and Volatile Profiles. Molecules 2018, 23, 2366. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, B.T.; Bujna, E.; Fekete, N.; Tran, A.T.M.; Rezessy-Szabo, J.M.; Prasad, R.; Nguyen, Q.D. Probiotic Beverage From Pineapple Juice Fermented With Lactobacillus and Bifidobacterium Strains. Front. Nutr. 2019, 6, 54. [Google Scholar] [CrossRef] [Green Version]
- Bancalari, E.; Castellone, V.; Bottari, B.; Gatti, M. Wild Lactobacillus Casei Group Strains: Potentiality to Ferment Plant Derived Juices. Foods 2020, 9, 314. [Google Scholar] [CrossRef] [Green Version]
- Stortz, J.A.; Raymond, S.L.; Mira, J.C.; Moldawer, L.L.; Mohr, A.M.; Efron, P.A. Murine Models of Sepsis and Trauma: Can We Bridge the Gap? ILAR J. 2017, 58, 90–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pohlmann, S.; Scheu, S.; Ziegler, V.; Schupp, N.; Henninger, C.; Fritz, G. Hepatic Rac1 GTPase Contributes to Liver-Mediated Basal Immune Homeostasis and LPS-Induced Endotoxemia. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 1277–1292. [Google Scholar] [CrossRef]
- Fitzpatrick, S.F.; Lambden, S.; Macias, D.; Puthucheary, Z.; Pietsch, S.; Mendil, L.; McPhail, M.J.W.; Johnson, R.S. 2-Hydroxyglutarate Metabolism Is Altered in an in Vivo Model of LPS Induced Endotoxemia. Front. Physiol. 2020, 11, 147. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Pérez, C.; García-Villanova, B.; Guerra-Hernández, E.; Verardo, V. Grape Seeds Proanthocyanidins: An Overview of In Vivo Bioactivity in Animal Models. Nutrients 2019, 11, 2435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cadoná, F.C.; Dantas, R.F.; de Mello, G.H.; Silva, F.P., Jr. Natural Products Targeting into Cancer Hallmarks: An Update on Caffeine, Theobromine, and (+)-Catechin. Crit. Rev. Food Sci. Nutr. 2022, 62, 7222–7241. [Google Scholar] [CrossRef]
- Gu, R.; Shi, Y.; Huang, W.; Lao, C.; Zou, Z.; Pan, S.; Huang, Z. Theobromine Mitigates IL-1β-Induced Oxidative Stress, Inflammatory Response, and Degradation of Type II Collagen in Human Chondrocytes. Int. Immunopharmacol. 2020, 82, 106226. [Google Scholar] [CrossRef]
- Costabile, A.; Bergillos-Meca, T.; Rasinkangas, P.; Korpela, K.; de Vs, W.M.; Gibson, G.R. Effects of Soluble Corn Fiber Alone or in Synbiotic Combination with Lactobacillus Rhamnosus GG and the Pilus-Deficient Derivative GG-PB12 on Fecal Microbiota, Metabolism, and Markers of Immune Function: A Randomized, Double-Blind, Placebo-Controlled, Crossover Study in Healthy Elderly (Saimes Study). Front. Immunol. 2017, 8, 1443. [Google Scholar] [CrossRef] [PubMed]
- Saliganti, V.; Kapila, R.; Kapila, S. Consumption of Probiotic Lactobacillus Rhamnosus (MTCC: 5897) Containing Fermented Milk Plays a Key Role in Development of the Immune System in Newborn Mice during the Suckling-Weaning Transition. Microbiol. Immunol. 2016, 60, 261–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spacova, I.; Lievens, E.; Verhoeven, T.; Steenackers, H.; Vanderleyden, J.; Lebeer, S.; Petrova, M.I. Expression of Fluorescent Proteins in Lactobacillus Rhamnosus to Study Host-Microbe and Microbe-Microbe Interactions. Microb. Biotechnol. 2018, 11, 317–331. [Google Scholar] [CrossRef] [Green Version]
- Petrova, M.I.; Imholz, N.C.E.; Verhoeven, T.L.A.; Balzarini, J.; van Damme, E.J.M.; Schols, D.; Vanderleyden, J.; Lebeer, S. Lectin-Like Molecules of Lactobacillus Rhamnosus GG Inhibit Pathogenic Escherichia Coli and Salmonella Biofilm Formation. PLoS ONE 2016, 11, e0161337. [Google Scholar] [CrossRef] [Green Version]
- Saliganti, V.; Kapila, R.; Sharma, R.; Kapila, S. Feeding Probiotic Lactobacillus Rhamnosus (MTCC 5897) Fermented Milk to Suckling Mothers Alleviates Ovalbumin-Induced Allergic Sensitisation in Mice Offspring. Br. J. Nutr. 2015, 114, 1168–1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, F.; Liu, L.; Cao, H.; Moore, D.J.; Washington, M.K.; Wang, B.; Peek, R.M.; Acra, S.A.; Polk, D.B. Neonatal Colonization of Mice with LGG Promotes Intestinal Development and Decreases Susceptibility to Colitis in Adulthood. Mucosal. Immunol. 2017, 10, 117–127. [Google Scholar] [CrossRef] [Green Version]
- Makrgeorgou, A.; Leonardi-Bee, J.; Bath-Hextall, F.J.; Murrell, D.F.; Tang, M.L.K.; Roberts, A.; Boyle, R.J. Probiotics for Treating Eczema. Cochrane Database Syst. Rev. 2018, 11, CD006135. [Google Scholar] [CrossRef]
- Panpetch, W.; Chancharoenthana, W.; Bootdee, K.; Nilgate, S.; Finkelman, M.; Tumwasorn, S.; Leelahavanichkul, A. Lactobacillus Rhamnosus L34 Attenuates Gut Translocation-Induced Bacterial Sepsis in Murine Models of Leaky Gut. Infect. Immun. 2017, 86, e00700-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vajro, P.; Mandato, C.; Licenziati, M.R.; Franzese, A.; Vitale, D.F.; Lenta, S.; Caropreso, M.; Vallone, G.; Meli, R. Effects of Lactobacillus Rhamnosus Strain GG in Pediatric Obesity-Related Liver Disease. J. Pediatr. Gastroenterol. Nutr. 2011, 52, 740–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, F.C.; de Barros, P.P.; Rossoni, R.D.; Junqueira, J.C.; Jorge, A.O.C. Lactobacillus Rhamnosus Inhibits Candida Albicans Virulence Factors in Vitro and Modulates Immune System in Galleria Mellonella. J. Appl. Microbiol. 2017, 122, 201–211. [Google Scholar] [CrossRef]
- Bleau, C.; Monges, A.; Rashidan, K.; Laverdure, J.P.; Lacroix, M.; van Calsteren, M.R.; Millette, M.; Savard, R.; Lamontagne, L. Intermediate Chains of Exopolysaccharides from Lactobacillus Rhamnosus RW-9595M Increase IL-10 Production by Macrophages. J. Appl. Microbiol. 2010, 108, 666–675. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | −1.41 | −1 | 0 | +1 | +1.41 |
|---|---|---|---|---|---|
| Inoculum density (OD600) | 0.77 | 1.00 | 1.55 | 2.10 | 2.33 |
| Pulp concentration (mg/mL) | 135.25 | 163.00 | 230.00 | 297.00 | 324.75 |
| Attribute | Moisture | Ashes | Proteins | Lipids | Carbohydrates * |
|---|---|---|---|---|---|
| Content (%) | 84.77 ± 0.2 | 3.3 ± 0.9 | 2.53 ± 0.05 | 0.35 ± 0.0 | 9.45 ± 0.0 |
| % Of Dry Mass | - | 21.77 | 16.62 | 2.30 | 62.05 |
| Assay | OD600nm | Pulp (mg/mL) | Growth (CFU/mL) | Final pH | Lactic Acid (g/L) | G/pH Ratio | G/[La] Ratio |
|---|---|---|---|---|---|---|---|
| 1 | 1.00 | 163.00 | 6.13 | 4.5 | 1.32 | 1.36 | 4.65 |
| 2 | 1.00 | 297.00 | 7.32 | 4.4 | 3.96 | 1.66 | 1.85 |
| 3 | 2.10 | 163.00 | 8.41 | 4.9 | 3.25 | 1.72 | 2.59 |
| 4 | 2.10 | 297.00 | 9.07 | 3.7 | 4.98 | 2.45 | 1.82 |
| 5 | 0.77 | 230.00 | 7.29 | 4.7 | 0.65 | 1.55 | 11.25 |
| 6 | 2.33 | 230.00 | 8.46 | 3.9 | 5.72 | 2.17 | 1.48 |
| 7 | 1.55 | 135.25 | 7.44 | 4 | 1.44 | 1.86 | 5.17 |
| 8 | 1.55 | 324.75 | 9.43 | 4.6 | 1.13 | 2.05 | 8.34 |
| 9 | 1.55 | 230.00 | 8.67 | 4.2 | 1.25 | 2.06 | 6.92 |
| 10 | 1.55 | 230.00 | 9.19 | 4.5 | 1.14 | 2.04 | 8.05 |
| Sample | RT (min) | [M-H] | Proposed Substance | Reference |
|---|---|---|---|---|
| Unfermented | 0.46 | 341.1159 | Saccharose | [30] |
| 0.70 | 191.0227 | Quinic acid | [30] | |
| 3.03 | 137.0244 | Tyrosol | [31] | |
| 5.17 | 353.1801 | Chlorogenic acid | [30] | |
| 5.86 | 173.1176 | Diethyl succinate | [31] | |
| Fermented | 2.78 | 181.0502 | Theobromine | [32] |
| 8.16 | 329.2318 | Vanillic acid glucoside | [30] | |
| 13.84 | 633.3425 | Epicatechin-(2beta- > 5,4beta- > 6)-ent-epicatechin Epicatechin-(2beta- > 7,4beta- > 6)-ent-epicatechin | [33] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zagmignan, A.; Mendes, Y.C.; Mesquita, G.P.; Santos, G.D.C.d.; Silva, L.d.S.; de Souza Sales, A.C.; Castelo Branco, S.J.d.S.; Junior, A.R.C.; Bazán, J.M.N.; Alves, E.R.; et al. Short-Term Intake of Theobroma grandiflorum Juice Fermented with Lacticaseibacillus rhamnosus ATCC 9595 Amended the Outcome of Endotoxemia Induced by Lipopolysaccharide. Nutrients 2023, 15, 1059. https://0-doi-org.brum.beds.ac.uk/10.3390/nu15041059
Zagmignan A, Mendes YC, Mesquita GP, Santos GDCd, Silva LdS, de Souza Sales AC, Castelo Branco SJdS, Junior ARC, Bazán JMN, Alves ER, et al. Short-Term Intake of Theobroma grandiflorum Juice Fermented with Lacticaseibacillus rhamnosus ATCC 9595 Amended the Outcome of Endotoxemia Induced by Lipopolysaccharide. Nutrients. 2023; 15(4):1059. https://0-doi-org.brum.beds.ac.uk/10.3390/nu15041059
Chicago/Turabian StyleZagmignan, Adrielle, Yasmim Costa Mendes, Gabrielle Pereira Mesquita, Gabrielle Damasceno Costa dos Santos, Lucas dos Santos Silva, Amanda Caroline de Souza Sales, Simeone Júlio dos Santos Castelo Branco, Alexsander Rodrigues Carvalho Junior, José Manuel Noguera Bazán, Edinalva Rodrigues Alves, and et al. 2023. "Short-Term Intake of Theobroma grandiflorum Juice Fermented with Lacticaseibacillus rhamnosus ATCC 9595 Amended the Outcome of Endotoxemia Induced by Lipopolysaccharide" Nutrients 15, no. 4: 1059. https://0-doi-org.brum.beds.ac.uk/10.3390/nu15041059