Twenty-Eight Fungal Secondary Metabolites Detected in Pig Feed Samples: Their Occurrence, Relevance and Cytotoxic Effects In Vitro

 , , and
, , and
Abstract
:1. Introduction
2. Results
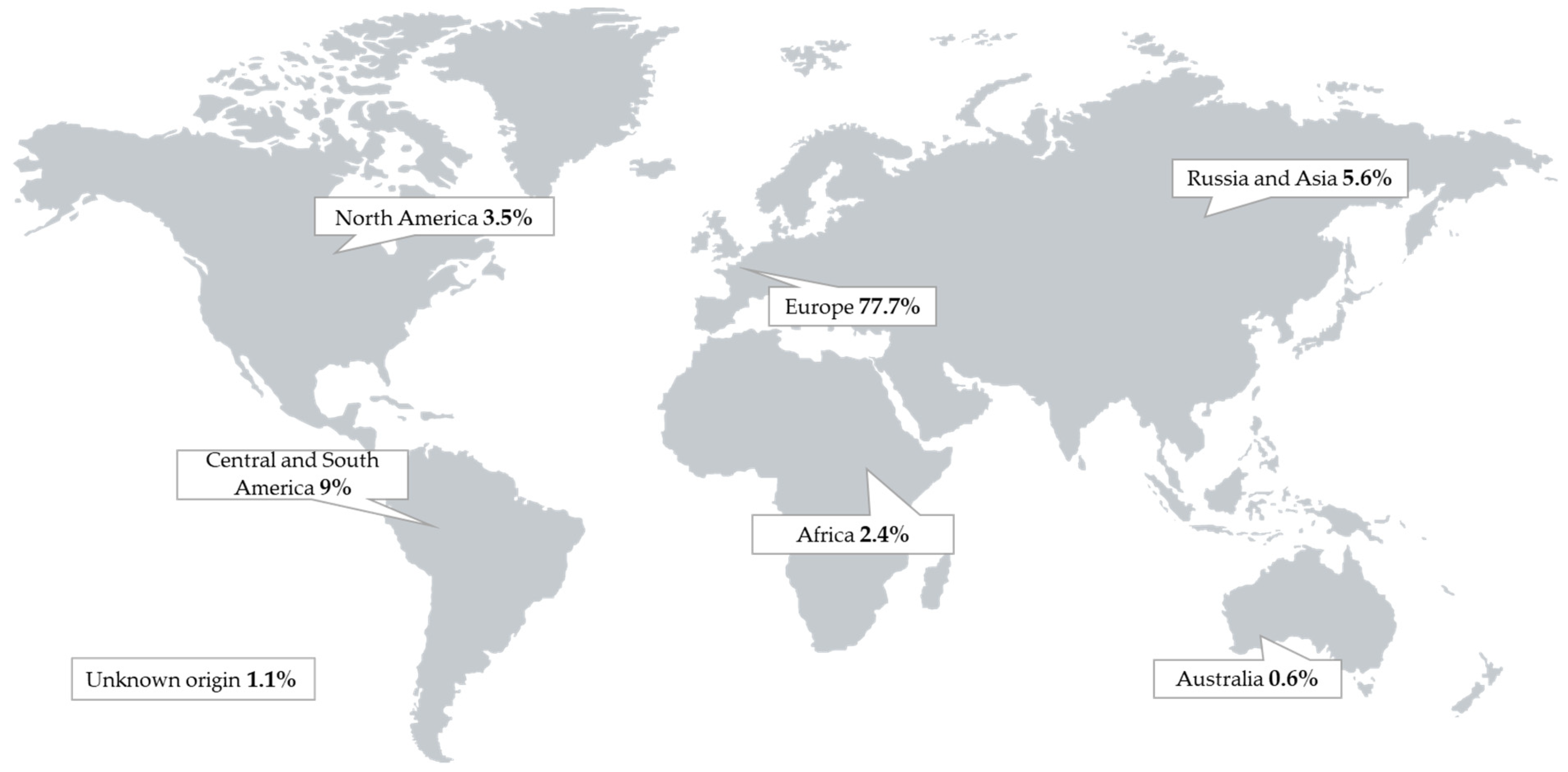
2.1. Occurrence Data
Secondary Fungal Metabolites in Finished Pig Feed Samples
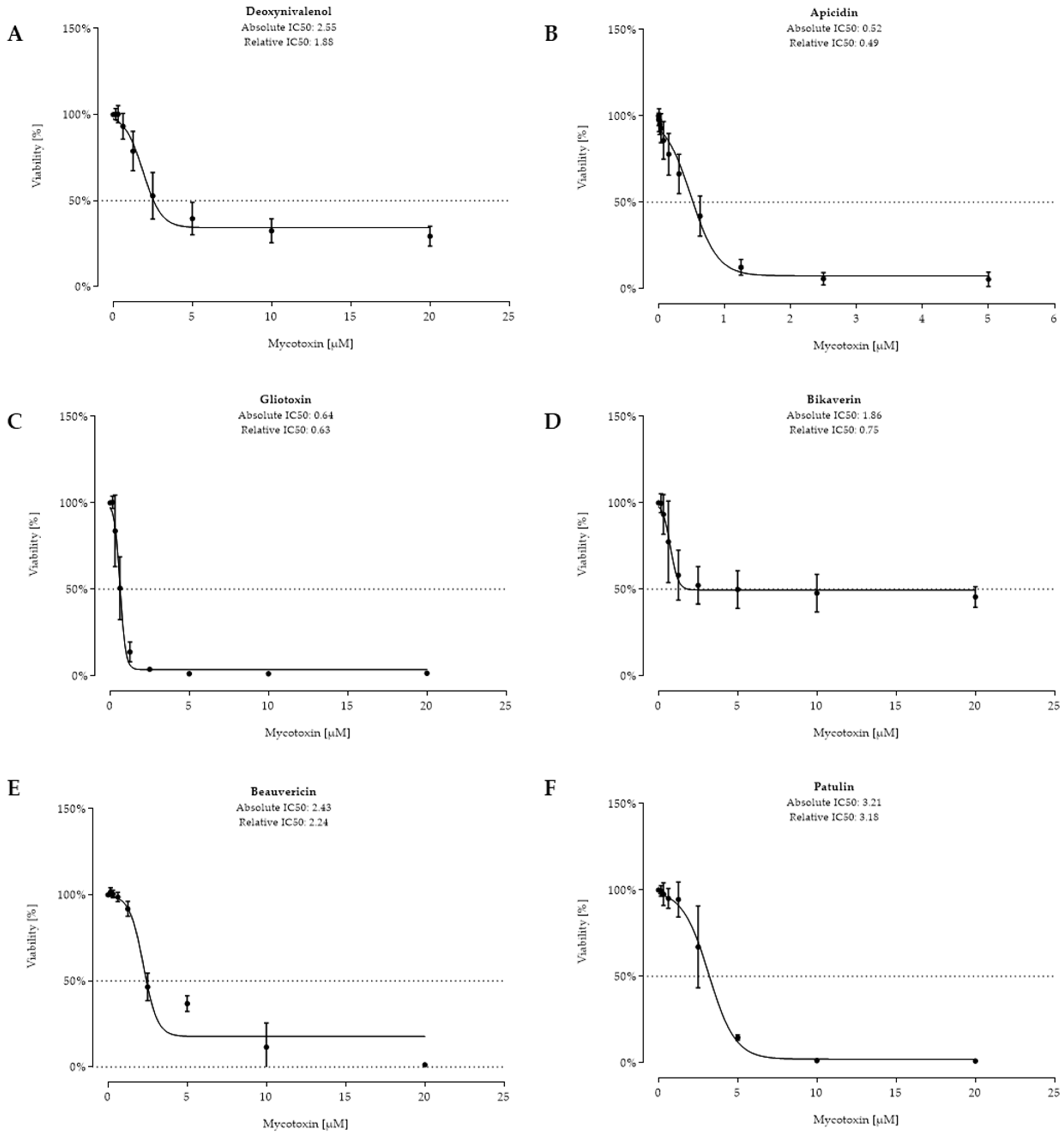
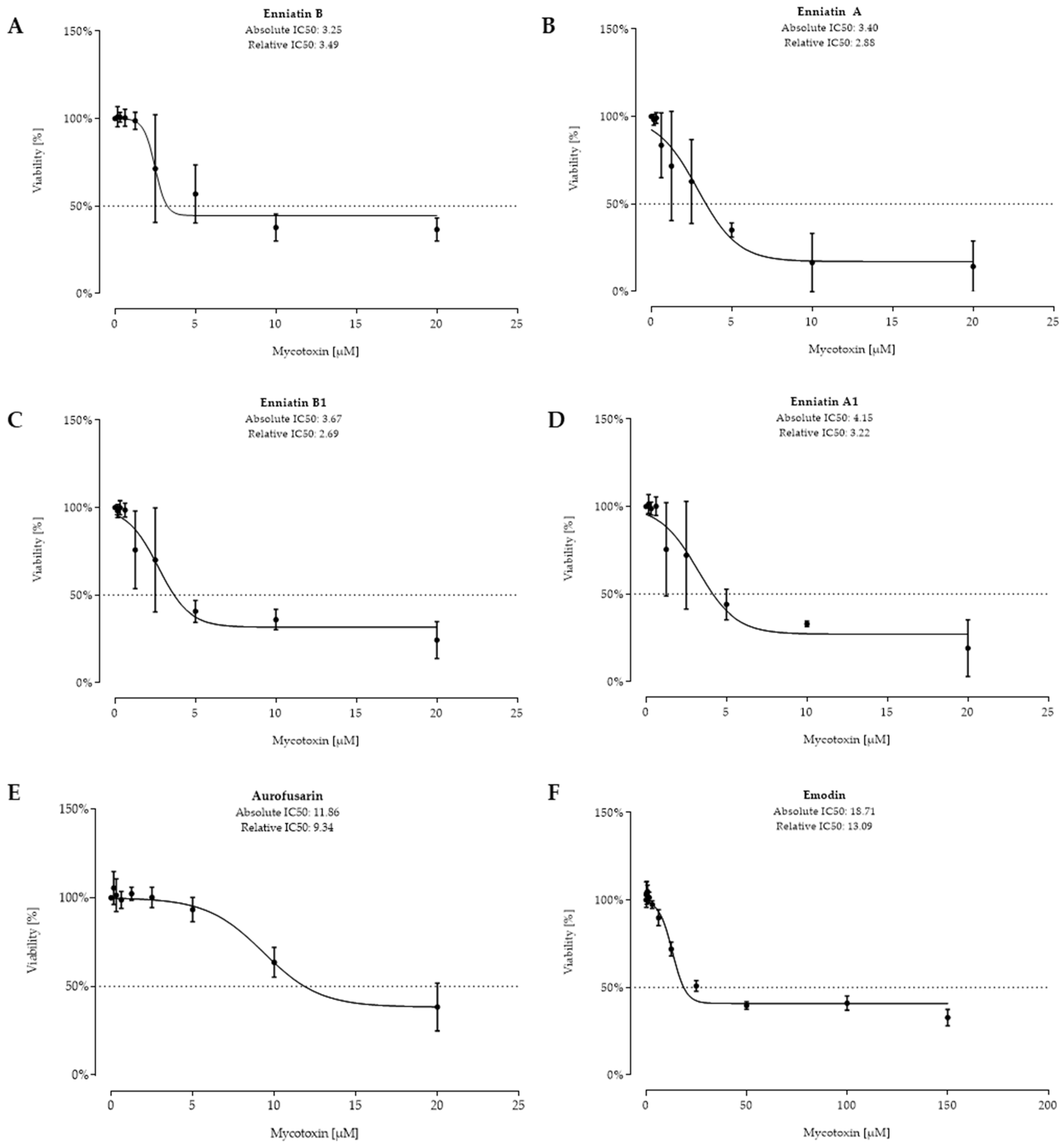
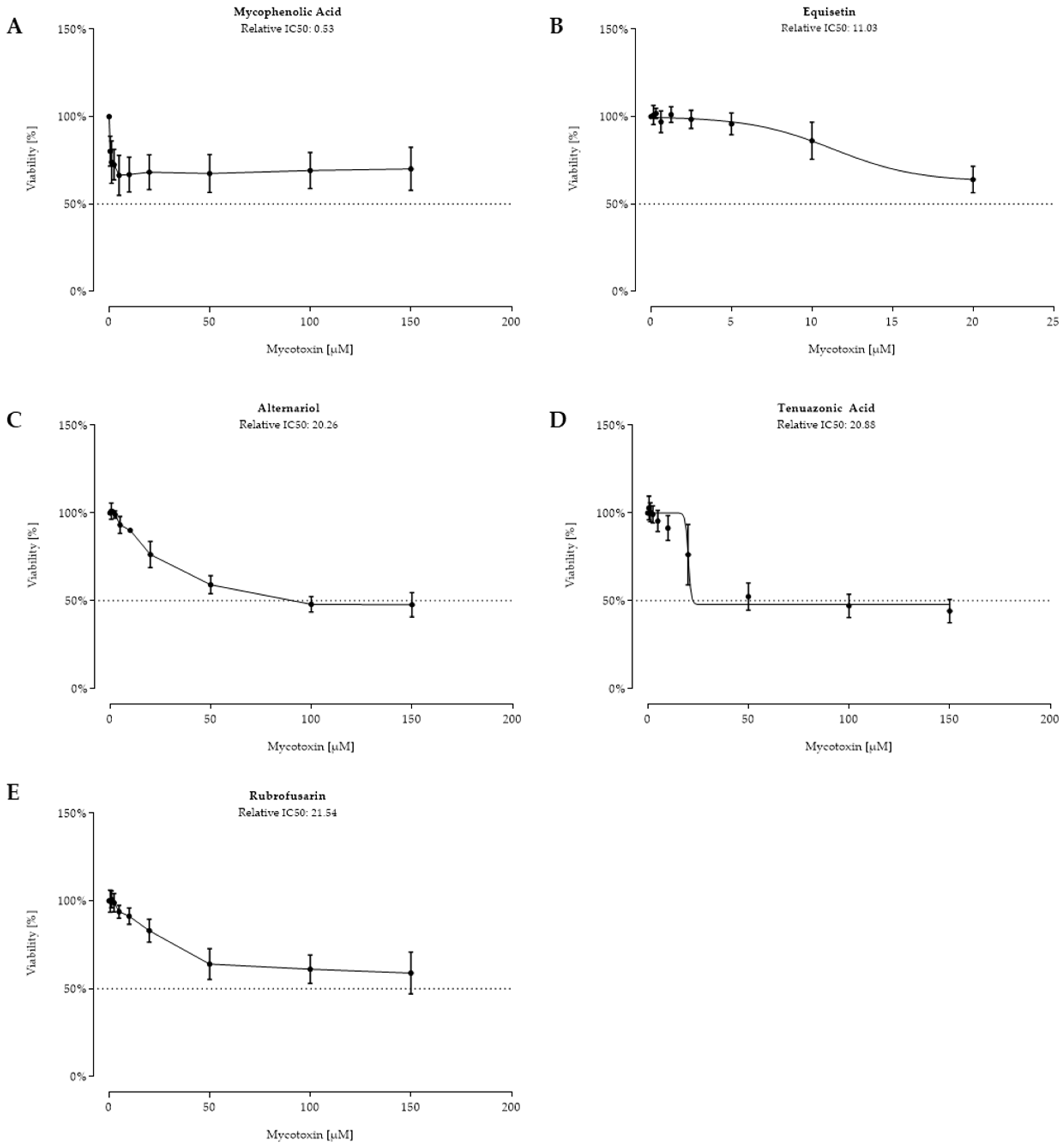
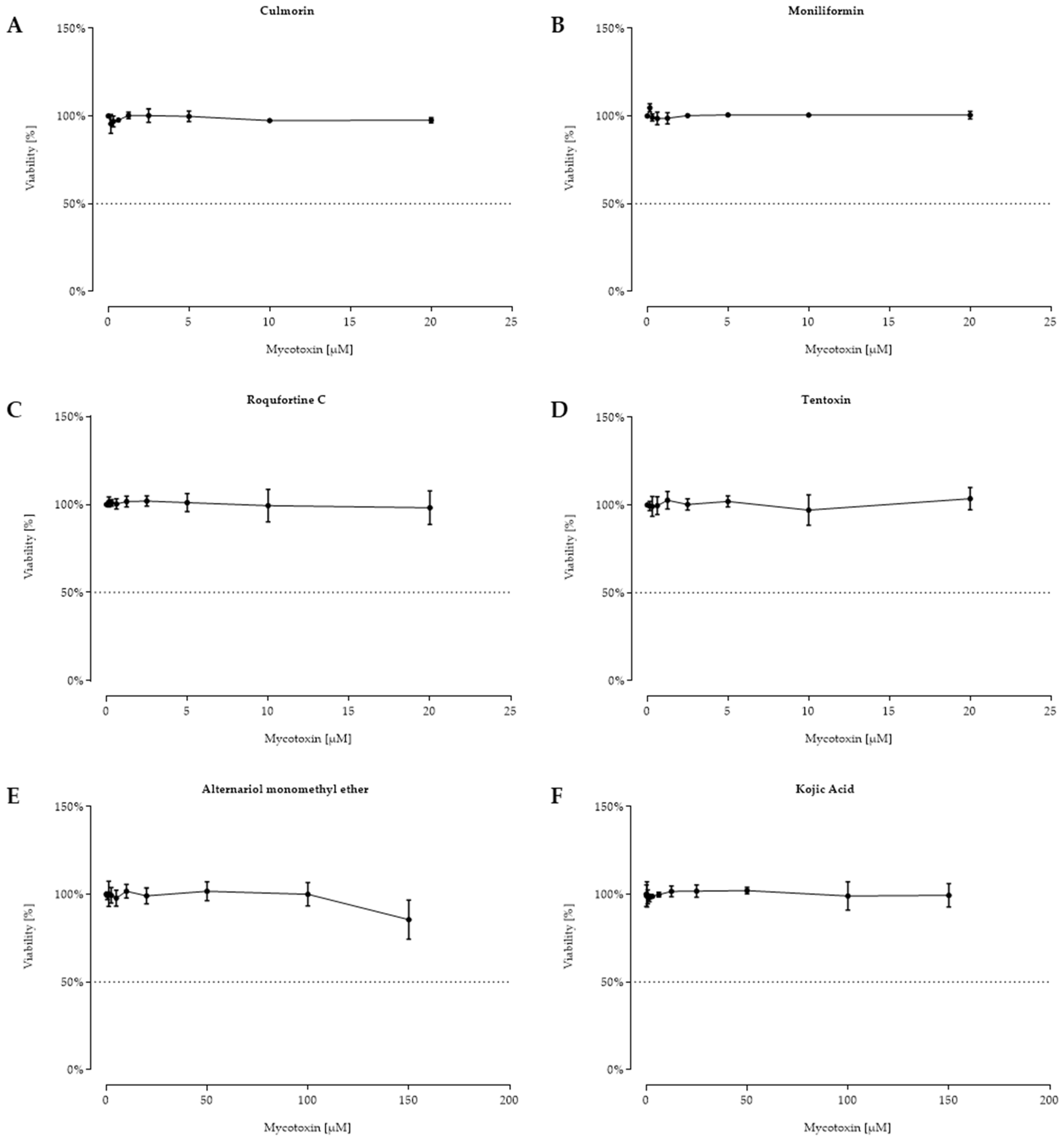
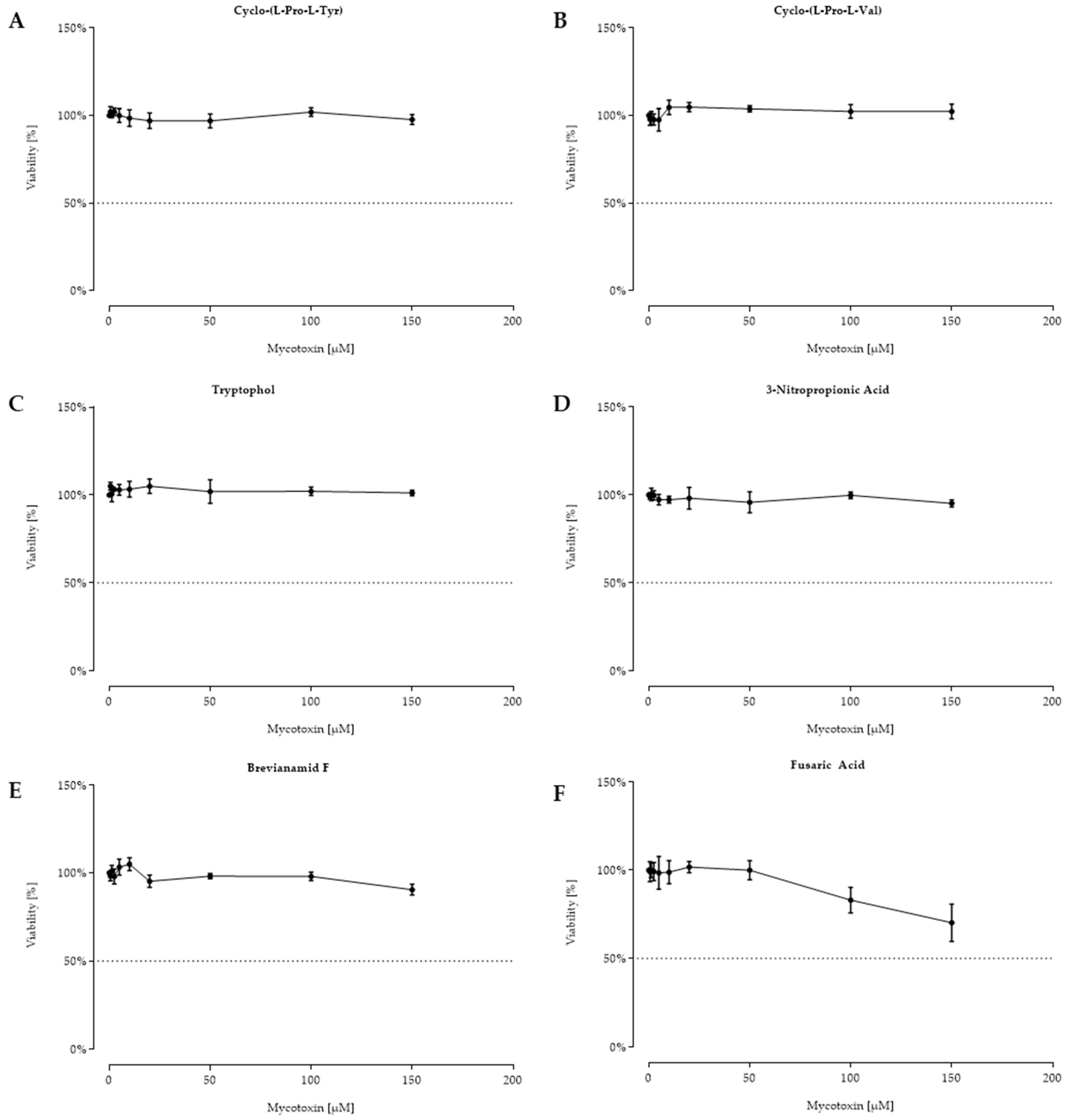
2.2. Cell Viability after 48 h Toxin Treatment
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Cell Culture
5.2. Material
5.3. Method
5.3.1. Cell Viability Assay
5.3.2. LC-MS/MS Multi-Analyte Method
5.3.3. Statistics and Evaluation
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bérdy, J. Bioactive microbial metabolites. J. Antibiot. 2005, 58, C1. [Google Scholar] [CrossRef] [PubMed]
- Commission Regulation (EC) No 1881/2006: Setting maximum levels for certain contaminantns in foodstuffs. Off. J. Eur. Union 2006, 364, 5–24.
- EFSA. Scientific opinion on the risk for public and animal health related to the presence of sterigmatocystin in food and feed. EFSA J. 2016, 11, 3254. [Google Scholar]
- EFSA. Scientific opinion on Ergot alkaloids in food and feed. EFSA J. 2012, 10, 158. [Google Scholar]
- EFSA. Scientific opinion on the risks to human and animal health related to the presence of beauvericin and enniatins in food and feed. EFSA J. 2014, 12, 3802. [Google Scholar] [CrossRef]
- Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Grasl-Kraupp, B.; Hogstrand, C.; et al. Risks to human and animal health related to the presence of moniliformin in food and feed. EFSA J. 2018, 16, 5082. [Google Scholar]
- Gruber-Dorninger, C.; Novak, B.; Nagl, V.; Berthiller, F. Emerging mycotoxins: Beyond traditionally determined food contaminants. J. Agric. Food Chem. 2017, 65, 7052–7070. [Google Scholar] [CrossRef] [PubMed]
- Jestoi, M. Emerging fusarium-mycotoxins fusaproliferin, beauvericin, enniatins, and moniliformin—A review. Crit. Rev. Food Sci. Nutr. 2008, 48, 21–49. [Google Scholar] [CrossRef]
- Fraeyman, S.; Croubels, S.; Devreese, M.; Antonissen, G. Emerging fusarium and alternaria mycotoxins: Occurrence, toxicity and toxicokinetics. Toxins 2017, 9, 228. [Google Scholar] [CrossRef]
- Sulyok, M.; Krska, R.; Schuhmacher, R. A liquid chromatography/tandem mass spectrometric multi-mycotoxin method for the quantification of 87 analytes and its application to semi-quantitative screening of moldy food samples. Anal. Bioanal. Chem. 2007, 389, 1505–1523. [Google Scholar] [CrossRef]
- Van der Fels-Klerx, H.J.; Klemsdal, S.; Hietaniemi, V.; Lindblad, M.; Ioannou-Kakouri, E.; van Asselt, E.D. Mycotoxin contamination of cereal grain commodities in relation to climate in North West Europe. Food Addit. Contam. Part A 2012, 29, 1581–1592. [Google Scholar] [CrossRef] [PubMed]
- Van der Fels-Klerx, H.; Kandhai, M.; Brynestad, S.; Dreyer, M.; Börjesson, T.; Martins, H.; Uiterwijk, M.; Morrison, E.; Booij, C. Development of a European system for identification of emerging mycotoxins in wheat supply chains. World Mycotoxin J. 2009, 2, 119–127. [Google Scholar] [CrossRef]
- Streit, E.; Schwab, C.; Sulyok, M.; Naehrer, K.; Krska, R.; Schatzmayr, G. Multi-mycotoxin screening reveals the occurrence of 139 different secondary metabolites in feed and feed ingredients. Toxins 2013, 5, 504–523. [Google Scholar] [CrossRef] [PubMed]
- Hietaniemi, V.; Rämö, S.; Yli-Mattila, T.; Jestoi, M.; Peltonen, S.; Kartio, M.; Sieviläinen, E.; Koivisto, T.; Parikka, P. Updated survey of Fusarium species and toxins in Finnish cereal grains. Food Addit. Contam. Part A 2016, 33, 831–848. [Google Scholar] [CrossRef] [PubMed]
- Lindblad, M.; Gidlund, A.; Sulyok, M.; Börjesson, T.; Krska, R.; Olsen, M.; Fredlund, E. Deoxynivalenol and other selected fusarium toxins in swedish wheat—Occurrence and correlation to specific fusarium species. Int. J. Food Microbiol. 2013, 167, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Juan, C.; Covarelli, L.; Beccari, G.; Colasante, V.; Mañes, J. Simultaneous analysis of twenty-six mycotoxins in durum wheat grain from Italy. Food Control 2016, 62, 322–329. [Google Scholar] [CrossRef]
- De Souza, M.D.L.M.; Sulyok, M.; Freitas-Silva, O.; Costa, S.S.; Brabet, C.; Machinski, M., Jr.; Sekiyama, B.L.; Vargas, E.A.; Krska, R.; Schuhmacher, R. Cooccurrence of mycotoxins in maize and poultry feeds from Brazil by liquid chromatography/tandem mass spectrometry. Sci. World J. 2013, 2013, 427369. [Google Scholar] [CrossRef] [PubMed]
- Kovalsky, P.; Kos, G.; Nährer, K.; Schwab, C.; Jenkins, T.; Schatzmayr, G.; Sulyok, M.; Krska, R. Co-occurrence of regulated, masked and emerging mycotoxins and secondary metabolites in finished feed and maize–An extensive survey. Toxins 2016, 8, 363. [Google Scholar] [CrossRef]
- Goossens, J.; Pasmans, F.; Verbrugghe, E.; Vandenbroucke, V.; De Baere, S.; Meyer, E.; Haesebrouck, F.; De Backer, P.; Croubels, S. Porcine intestinal epithelial barrier disruption by the Fusarium mycotoxins deoxynivalenol and T-2 toxin promotes transepithelial passage of doxycycline and paromomycin. BMC Vet. Res. 2012, 8, 245. [Google Scholar] [CrossRef]
- Ficheux, A.S.; Sibiril, Y.; Parent-Massin, D. Effects of beauvericin, enniatin b and moniliformin on human dendritic cells and macrophages: An invitro study. Toxicon 2013, 71, 1–10. [Google Scholar] [CrossRef]
- Zouaoui, N.; Mallebrera, B.; Berrada, H.; Abid-Essefi, S.; Bacha, H.; Ruiz, M.J. Cytotoxic effects induced by patulin, sterigmatocystin and beauvericin on CHO-K1 cells. Food Chem. Toxicol. 2016, 89, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Bensassi, F.; Gallerne, C.; Sharaf El Dein, O.; Hajlaoui, M.R.; Bacha, H.; Lemaire, C. Cell death induced by the Alternaria mycotoxin Alternariol. Toxicol. Vitr. 2012, 26, 915–923. [Google Scholar] [CrossRef] [PubMed]
- Jarolim, K.; Wolters, K.; Woelflingseder, L.; Pahlke, G.; Beisl, J.; Puntscher, H.; Braun, D.; Sulyok, M.; Warth, B.; Marko, D. The secondary Fusarium metabolite aurofusarin induces oxidative stress, cytotoxicity and genotoxicity in human colon cells. Toxicol. Lett. 2018, 284, 170–183. [Google Scholar] [CrossRef] [PubMed]
- Malachová, A.; Sulyok, M.; Beltrán, E.; Berthiller, F.; Krska, R. Optimization and validation of a quantitative liquid chromatography-tandem mass spectrometric method covering 295 bacterial and fungal metabolites including all regulated mycotoxins in four model food matrices. J. Chromatogr. A 2014, 1362, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Vaclavikova, M.; Malachova, A.; Veprikova, Z.; Dzuman, Z.; Zachariasova, M.; Hajslova, J. “Emerging” mycotoxins in cereals processing chains: Changes of enniatins during beer and bread making. Food Chem. 2013, 136, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Vidal, A.; Ouhibi, S.; Ghali, R.; Hedhili, A.; De Saeger, S.; De Boevre, M. The mycotoxin patulin: An updated short review on occurrence, toxicity and analytical challenges. Food Chem. Toxicol. 2019, 129, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Kosalec, I.; Pepeljnjak, S. Chemistry and biological effects of gliotoxin. Arh. Hig. Rada Toksikol. 2004, 55, 313–320. [Google Scholar] [PubMed]
- Kanora, A.; Maes, D. The role of mycotoxins in pig reproduction: A review. Vet. Med. 2009, 54, 565–576. [Google Scholar] [CrossRef]
- Schierack, P.; Nordhoff, M.; Pollmann, M.; Weyrauch, K.D.; Amasheh, S.; Lodemann, U.; Jores, J.; Tachu, B.; Kleta, S.; Blikslager, A.; et al. Characterization of a porcine intestinal epithelial cell line for in vitro studies of microbial pathogenesis in swine. Histochem. Cell Biol. 2006, 125, 293–305. [Google Scholar] [CrossRef]
- Nossol, C.; Barta-Böszörményi, A.; Kahlert, S.; Zuschratter, W.; Faber-Zuschratter, H.; Reinhardt, N.; Ponsuksili, S.; Wimmers, K.; Diesing, A.K.; Rothkötter, H.J. Comparing two intestinal porcine epithelial cell lines (IPECs): Morphological differentiation, function and metabolism. PLoS ONE 2015, 10, e0132323. [Google Scholar] [CrossRef]
- Sebaugh, J.L. Guidelines for accurate EC50/IC50 estimation. Pharm. Stat. 2011, 10, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Pestka, J.J. Deoxynivalenol: Mechanisms of action, human exposure, and toxicological relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Paulick, M.; Winkler, J.; Kersten, S.; Schatzmayr, D.; Schwartz-Zimmermann, H.E.; Dänicke, S. Studies on the bioavailability of deoxynivalenol (DON) and DON sulfonate (DONS) 1, 2, and 3 in pigs fed with sodium sulfite-treated DON-contaminated maize. Toxins 2015, 7, 4622–4644. [Google Scholar] [CrossRef] [PubMed]
- Han, J.W.; Ahn, S.H.; Park, S.H.; Wang, S.Y.; Bae, G.U.; Seo, D.W.; Kwon, H.K.; Hong, S.; Hoi, Y.; Lee, Y.W.; et al. Apicidin, a histone deacetylase inhibitor, inhibits proliferation of tumor cells via induction of p21(WAF1/Cip1) and gelsolin. Cancer Res. 2000, 60, 6068–6074. [Google Scholar] [PubMed]
- Darkin-Rattray, S.J.; Gurnett, A.M.; Myers, R.W.; Dulski, P.M.; Crumley, T.M.; Allocco, J.J.; Cannova, C.; Meinke, P.T.; Colletti, S.L.; Bednarek, M.A.; et al. Apicidin: A novel antiprotozoal agent that inhibits parasite histone deacetylase. Proc. Natl. Acad. Sci. USA 1996, 93, 13143–13147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.S.; Lee, K.R.; Kim, J.C.; Lim, S.H.; Seo, J.A.; Lee, Y.W. A hemorrhagic factor (apicidin) produced by toxic Fusarium isolates from soybean seeds. Appl. Environ. Microbiol. 1999, 65, 126–130. [Google Scholar]
- Pena, G.A.; Pereyra, C.M.; Armando, M.R.; Chiacchiera, S.M.; Magnoli, C.E.; Orlando, J.L.; Dalcero, A.M.; Rosa, C.A.R.; Cavaglieri, L.R. Aspergillus fumigatus toxicity and gliotoxin levels in feedstuff for domestic animals and pets in Argentina. Lett. Appl. Microbiol. 2010, 50, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Nieminen, S.M.; Mäki-Paakkanen, J.; Hirvonen, M.R.; Roponen, M.; Von Wright, A. Genotoxicity of gliotoxin, a secondary metabolite of Aspergillus fumigatus, in a battery of short-term test systems. Mutat. Res. 2002, 520, 161–170. [Google Scholar] [CrossRef]
- Upperman, J.S.; Potoka, D.A.; Zhang, X.R.; Wong, K.; Zamora, R.; Ford, H.R. Mechanism of intestinal-derived fungal sepsis by gliotoxin, a fungal metabolite. J. Pediatr. Surg. 2003, 38, 966–970. [Google Scholar] [CrossRef]
- Limón, M.C.; Rodríguez-Ortiz, R.; Avalos, J. Bikaverin production and applications. Appl. Microbiol. Biotechnol. 2010, 87, 21–29. [Google Scholar] [CrossRef]
- Deshmukh, R.; Mathew, A.; Purohit, H.J. Characterization of antibacterial activity of bikaverin from Fusarium sp. HKF 15. J. Biosci. Bioeng. 2014, 117, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Fuska, J.; Proksa, B.; Fuskova, A. New potential cytotoxic and antitumor substances. I. In vitro effect of bikaverin and its derivatives on cells of certain tumors. Neoplasma 1975, 22, 335–338. [Google Scholar] [PubMed]
- Font, G.; Prosperini, A.; Ruiz, M.J. Cytotoxicity, bioaccessibility and transport by Caco-2 cells of enniatins and beauvericin. Toxicol. Lett. 2011, 205, S159. [Google Scholar] [CrossRef]
- Prosperini, A.; Juan-García, A.; Font, G.; Ruiz, M.J. Beauvericin-induced cytotoxicity via ROS production and mitochondrial damage in Caco-2 cells. Toxicol. Lett. 2013, 222, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Meca, G.; Font, G.; Ruiz, M.J. Comparative cytotoxicity study of enniatins A, A1, A2, B, B1, B4 and J3 on Caco-2 cells, Hep-G2 and HT-29. Food Chem. Toxicol. 2011, 49, 2464–2469. [Google Scholar] [CrossRef] [PubMed]
- Fraeyman, S.; Meyer, E.; Devreese, M.; Antonissen, G.; Demeyere, K.; Haesebrouck, F.; Croubels, S. Comparative in vitro cytotoxicity of the emerging Fusarium mycotoxins beauvericin and enniatins to porcine intestinal epithelial cells. Food Chem. Toxicol. 2018, 121, 566–572. [Google Scholar] [CrossRef]
- Springler, A.; Vrubel, G.J.; Mayer, E.; Schatzmayr, G.; Novak, B. Effect of Fusarium-derived metabolites on the barrier integrity of differentiated intestinal porcine epithelial cells (IPEC-J2). Toxins 2016, 8, 345. [Google Scholar] [CrossRef] [PubMed]
- Broekaert, N.; Devreese, M.; Demeyere, K.; Berthiller, F.; Michlmayr, H.; Varga, E.; Adam, G.; Meyer, E.; Croubels, S. Comparative in vitro cytotoxicity of modified deoxynivalenol on porcine intestinal epithelial cells. Food Chem. Toxicol. 2016, 95, 103–109. [Google Scholar] [CrossRef]
- Mahnine, N.; Meca, G.; Elabidi, A.; Fekhaoui, M.; Saoiabi, A.; Font, G.; Mañes, J.; Zinedine, A. Further data on the levels of emerging Fusarium mycotoxins enniatins (A, A1, B, B1), beauvericin and fusaproliferin in breakfast and infant cereals from Morocco. Food Chem. 2011, 63, 161–165. [Google Scholar] [CrossRef]
- Devreese, M.; Broekaert, N.; De Mil, T.; Fraeyman, S.; De Backer, P.; Croubels, S. Pilot toxicokinetic study and absolute oral bioavailability of the Fusarium mycotoxin enniatin B1 in pigs. Food Chem. Toxicol. 2014, 63, 161–165. [Google Scholar] [CrossRef] [Green Version]
- Meca, G.; Mañes, J.; Font, G.; Ruiz, M.J. Study of the potential toxicity of enniatins A, A 1, B, B 1 by evaluation of duodenal and colonic bioavailability applying an invitro method by Caco-2 cells. Toxicon 2012, 59, 1–11. [Google Scholar] [CrossRef]
- Saleh, I.; Goktepe, I. The characteristics, occurrence, and toxicological effects of patulin. Food Chem. Toxicol. 2019, 129, 301–311. [Google Scholar] [CrossRef]
- Puel, O.; Galtier, P.; Oswald, I.P. Biosynthesis and toxicological effects of patulin. Toxins 2010, 2, 613–631. [Google Scholar] [CrossRef]
- Dvorska, J.E.; Surai, P.F.; Speake, B.K.; Sparks, N.H.C. Effect of the mycotoxin aurofusarin on the antioxidant composition and fatty acid profile of quail eggs. Br. Poult. Sci. 2001, 42, 643–649. [Google Scholar] [CrossRef]
- Dvorska, J.E.; Surai, P.F.; Speake, B.K.; Sparks, N.H.C. Antioxidant systems of the developing quail embryo are compromised by mycotoxin aurofusarin. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2002, 131, 197–205. [Google Scholar] [CrossRef]
- Tola, S.; Bureau, D.P.; Hooft, J.M.; Beamish, F.W.H.; Sulyok, M.; Krska, R.; Encarnação, P.; Petkam, R. Effects of wheat naturally contaminated with Fusarium mycotoxins on growth performance and selected health indices of red tilapia (Oreochromis niloticus × O. mossambicus). Toxins 2015, 7, 1929–1944. [Google Scholar] [CrossRef]
- Uhlig, S.; Jestoi, M.; Kristin Knutsen, A.; Heier, B.T. Multiple regression analysis as a tool for the identification of relations between semi-quantitative LC-MS data and cytotoxicity of extracts of the fungus Fusarium avenaceum (syn. F. arthrosporioides). Toxicon 2006, 48, 567–579. [Google Scholar] [CrossRef]
- Vejdovszky, K.; Warth, B.; Sulyok, M.; Marko, D. Non-synergistic cytotoxic effects of Fusarium and Alternaria toxin combinations in Caco-2 cells. Toxicol. Lett. 2016, 241, 1–8. [Google Scholar] [CrossRef]
- Izhaki, I. Emodin—A secondary metabolite with multiple ecological functions in higher plants. N. Phytol. 2002, 155, 205–217. [Google Scholar] [CrossRef]
- Springler, A.; Hessenberger, S.; Reisinger, N.; Kern, C.; Nagl, V.; Schatzmayr, G.; Mayer, E. Deoxynivalenol and its metabolite deepoxy-deoxynivalenol: Multi-parameter analysis for the evaluation of cytotoxicity and cellular effects. Mycotoxin Res. 2017, 33, 25–37. [Google Scholar] [CrossRef]
- Fontaine, K.; Mounier, J.; Coton, E.; Hymery, N. Individual and combined effects of roquefortine C and mycophenolic acid on human monocytic and intestinal cells. World Mycotoxin J. 2015, 9, 51–62. [Google Scholar] [CrossRef]
- Juan-Garcí, A.; Fernández-Blanco, C.; Font, G.; Ruiz, M.J. Toxic effects of alternariol by in vitro assays: A review. Rev. Toxicol. 2015, 31, 196–203. [Google Scholar]
- Juan-García, A.; Juan, C.; König, S.; Ruiz, M.J. Cytotoxic effects and degradation products of three mycotoxins: Alternariol, 3-acetyl-deoxynivalenol and 15-acetyl-deoxynivalenol in liver hepatocellular carcinoma cells. Toxicol. Lett. 2015, 235, 8–16. [Google Scholar] [CrossRef]
- Woelflingseder, L.; Warth, B.; Vierheilig, I.; Schwartz-Zimmermann, H.; Hametner, C.; Nagl, V.; Novak, B.; Šarkanj, B.; Berthiller, F.; Adam, G.; et al. The Fusarium metabolite culmorin suppresses the in vitro glucuronidation of deoxynivalenol. Arch. Toxicol. 2019, 93, 1729–1743. [Google Scholar] [CrossRef] [Green Version]
- Weber, J.; Vaclavikova, M.; Wiesenberger, G.; Haider, M.; Hametner, C.; Fröhlich, J.; Berthiller, F.; Adam, G.; Mikula, H.; Fruhmann, P. Chemical synthesis of culmorin metabolites and their biologic role in culmorin and acetyl-culmorin treated wheat cells. Org. Biomol. Chem. 2018, 16, 2043–2048. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.C.; Ledoux, D.R.; Bermudez, A.J.; Fritsche, K.L.; Rottinghaus, G.E. Effects of moniliformin on performance and immune function of broiler chicks. Poult. Sci. 2000, 79, 26–32. [Google Scholar] [CrossRef]
- Wattana-Amorn, P.; Charoenwongsa, W.; Williams, C.; Crump, M.P.; Apichaisataienchote, B. Antibacterial activity of cyclo(L-Pro-L-Tyr) and cyclo(D-Pro-L-Tyr) from Streptomyces sp. strain 22–4 against phytopathogenic bacteria. Nat. Prod. Res. 2016, 30, 1980–1983. [Google Scholar] [CrossRef]
- Kosalec, I.; Ramić, S.; Jelić, D.; Antolović, R.; Pepeljnjak, S.; Kopjar, N. Assessment of tryptophol genotoxicity in four cell lines in vitro: A pilot study with alkaline comet assay. Arh. Hig. Rada Toksikol. 2011, 62, 41–49. [Google Scholar] [CrossRef]
- Kosalec, I.; Šafranić, A.; Pepeljnjak, S.; Bačun-Družina, V.; Ramić, S.; Kopjar, N. Genotoxicity of tryptophol in a battery of short-term assays on human white blood cells in vitro. Basic Clin. Pharmacol. Toxicol. 2008, 102, 443–452. [Google Scholar] [CrossRef]
- Pihlström, T. Method Validation and Quality Control Procedures for Pesticide Residues Analysis in Food and Feed; DG SANCO/12495/2011; European Commission Directorate-General for Health and Food Safety: Brussels, Belgium, 2011; pp. 1–41. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rank | Metabolite | Mean Concentration | Median Concentration | Maximum Concentration | Prevalence |
|---|---|---|---|---|---|
| 1 | Cyclo-(L-Pro-L-Tyr) | 321 | 105 | 34,910 | 87.6% |
| 2 | Moniliformin, t > 2.0 ppb | 66 | 17 | 2053 | 82.6% |
| 3 | Enniatin B | 73 | 30 | 1514 | 82.2% |
| 4 | Enniatin B1 | 78 | 34 | 1846 | 82.1% |
| 5 | Aurofusarin | 932 | 210 | 85,360 | 80.7% |
| 6 | Culmorin | 905 | 118 | 157,114 | 79.7% |
| 7 | Enniatin A1 | 30 | 14 | 549 | 77.4% |
| 8 | Deoxynivalenol, t > 1.5 ppb | 634 | 193 | 34,862 | 77.0% |
| 9 | Tryptophol | 291 | 138 | 10,270 | 75.4% |
| 10 | Zearalenone* | 126 | 18 | 9905 | 73.3% |
| 11 | 15-Hydroxy-culmorin* | 468 | 152 | 19,320 | 73.2% |
| 12 | Beauvericin, t > 2.0 ppb | 17 | 6 | 413 | 68.7% |
| 13 | Emodin | 17 | 4 | 591 | 69.3% |
| 14 | Infectopyron* | 983 | 294 | 66,094 | 65.9% |
| 15 | Brevianamid F | 44 | 25 | 1170 | 65.2% |
| 16 | Equisetin | 50 | 11 | 6120 | 64.2% |
| 17 | Cyclo-(L-Pro-L-Val) | 187 | 71 | 5042 | 62.1% |
| 18 | DON-3-glucoside*, t > 15.0 ppb | 74 | 22 | 2741 | 62.8% |
| 19 | Asperglaucide* | 113 | 31 | 6232 | 61.8% |
| 20 | Nivalenol*, t > 15.0 ppb | 65 | 31 | 1143 | 56.7% |
| 21 | 3-Nitro-propionic acid | 16 | 6 | 509 | 56.5% |
| 22 | Tenuazonic acid | 255 | 82 | 9910 | 55.0% |
| 23 | Apicidin | 22 | 8 | 1568 | 52.2% |
| 24 | Alternariol | 17 | 4 | 2508 | 50.7% |
| 25 | Enniatin A | 7 | 3 | 307 | 49.5% |
| 26 | Alternariol monomethyl ether | 6 | 3 | 208 | 40.3% |
| 27 | Tentoxin | 8 | 3 | 157 | 37.3% |
| 28 | Kojic acid | 192 | 78 | 3030 | 33.7% |
| 29 | Bikaverin | 58 | 19 | 1564 | 29.8% |
| 30 | Fusaric acid | 333 | 81 | 5566 | 13.0% |
| 31 | Mycophenolic acid | 39 | 8 | 1178 | 13.1% |
| 32 | Rubrofusarin | 199 | 38 | 1696 | 2.3% |
| 33 | Gliotoxin | 5 | 5 | 6 | 0.2% |
| 34 | Patulin | <LOD | <LOD | <LOD | n.a. |
| Rank | Fungal Metabolite | IC50 Value (µM) | IC50 Value (µg/kg) | Occurrence | |||
|---|---|---|---|---|---|---|---|
| Absolute | Relative | Absolute | Relative | Median (µg/kg) | Maximum (µg/kg) | ||
| 1 | API | 0.52 | 0.49 | 324 | 306 | 8 | 1568 |
| 2 | GLIO | 0.64 | 0.63 | 209 | 206 | 5 | 6 |
| 3 | BIK | 1.86 | 0.75 | 711 | 287 | 19 | 1564 |
| 4 | BEA | 2.43 | 2.24 | 1905 | 1756 | 6 | 413 |
| Control | DON | 2.55 | 1.88 | 756 | 557 | 193 | 34,862 |
| 5 | PAT | 3.21 | 3.18 | 495 | 490 | <LOD | <LOD |
| 6 | EnnB | 3.25 | 3.49 | 2079 | 2233 | 30 | 1514 |
| 7 | EnnA | 3.40 | 2.88 | 2319 | 1964 | 3 | 307 |
| 8 | EnnB1 | 3.67 | 2.69 | 2400 | 1759 | 34 | 1846 |
| 9 | EnnA1 | 4.15 | 3.22 | 2772 | 2151 | 14 | 549 |
| 10 | AUR | 11.86 | 9.34 | 6766 | 5329 | 210 | 85,360 |
| 11 | EMO | 18.71 | 13.09 | 5056 | 3537 | 4 | 591 |
| 12 | MPA | nc | 0.53 | nc | 170 | 8 | 1178 |
| 13 | EQUI | nc | 11.03 | nc | 4120 | 11 | 6120 |
| 14 | ALT | nc | 20.26 | nc | 5232 | 4 | 2508 |
| 15 | TeA | nc | 20.88 | nc | 4761 | 82 | 9910 |
| 16 | RUB | nc | 21.54 | nc | 5865 | 38 | 1696 |
| Chemical | Purity | Solvent | Highest Tested Concentration (µM) | Company |
|---|---|---|---|---|
| Alternariol (Alternaria sp.) | ≈96% | DMSO | 150 | Sigma-Aldrich |
| Alternariol monoethyl ether (Alternaria alternata) | ≥98% | DMSO | 150 | Sigma-Aldrich |
| Apicidin (Fusarium sp.) | ≥95% | DMSO | 5 | Santa Cruz |
| Aurofusarin (Fusarium graminearum) | ≥97% | DMSO | 20 | AdipoGen Life Sciences |
| Beauvericin (Beauveria sp.) | ≥95% | DMSO | 20 | AdipoGen Life Sciences |
| Bikaverin (Fusarium sp.) | 95% | DMSO | 20 | Santa Cruz |
| Brevianamid F | >98% | DMSO | 150 | MedChem Express |
| Cyclo(L-Pro-L-Tyr) | >98% | DMSO | 150 | BioAustralis |
| Cyclo(L-Pro-L-Val) | >98% | DMSO | 150 | BioAustralis |
| Culmorin | 100% | ACN | 20 | Generous gift from Dr. Fruhmann |
| Deoxynivalenol (Fusarium sp.) | ≥95% | Distilled water | 20 | Biopure |
| Emodin | ≥97% | DMSO | 150 | Sigma-Aldrich |
| Enniatin A, A1, B, B1 (Gnomonia errabunda) | ≥95% | DMSO | 20 | Sigma-Aldrich |
| Equisetin (Fusarium equiseti) | >99% | DMSO | 20 | Santa Cruz |
| Fusaric acid (Gibberella fujikuroi) | ≥98% | 96% EtOH | 150 | Sigma-Aldrich |
| Gliotoxin (Gladiocladium fimbriatum) | ≥97% | DMSO | 20 | Santa Cruz |
| Kojic acid | ≥99% | Distilled water | 150 | Sigma-Aldrich |
| Moniliformin (Fusarium moniliforme) | ≥99% | Distilled water | 20 | BioAustralis |
| Mycophenolic acid (Penicillium brevicompactum) | ≥98% | DMSO | 150 | Sigma-Aldrich |
| Patulin | 98% | DMSO | 20 | Santa Cruz |
| Roquefortine C (Penicillium sp.) | ≥98% | DMSO | 20 | AdipoGen Life Sciences |
| Tentoxin (Alternaria tenuis) | ≥95% | 70% EtOH | 20 | Sigma-Aldrich |
| Tenuazonic acid (Alternaria alternata) | ≥98% | DMSO | 150 | AdipoGen Life Sciences |
| Tryptophol | >98% | DMSO | 150 | AdipoGen Life Sciences |
| 3-Nitropropionic acid | ≥97% | 70% EtOH | 150 | Sigma-Aldrich |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Novak, B.; Rainer, V.; Sulyok, M.; Haltrich, D.; Schatzmayr, G.; Mayer, E. Twenty-Eight Fungal Secondary Metabolites Detected in Pig Feed Samples: Their Occurrence, Relevance and Cytotoxic Effects In Vitro. Toxins 2019, 11, 537. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11090537
Novak B, Rainer V, Sulyok M, Haltrich D, Schatzmayr G, Mayer E. Twenty-Eight Fungal Secondary Metabolites Detected in Pig Feed Samples: Their Occurrence, Relevance and Cytotoxic Effects In Vitro. Toxins. 2019; 11(9):537. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11090537
Chicago/Turabian StyleNovak, Barbara, Valentina Rainer, Michael Sulyok, Dietmar Haltrich, Gerd Schatzmayr, and Elisabeth Mayer. 2019. "Twenty-Eight Fungal Secondary Metabolites Detected in Pig Feed Samples: Their Occurrence, Relevance and Cytotoxic Effects In Vitro" Toxins 11, no. 9: 537. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins11090537