Risk Assessment of Pectenotoxins in New Zealand Bivalve Molluscan Shellfish, 2009–2019

 ,
,
Abstract
:1. Introduction
2. Results
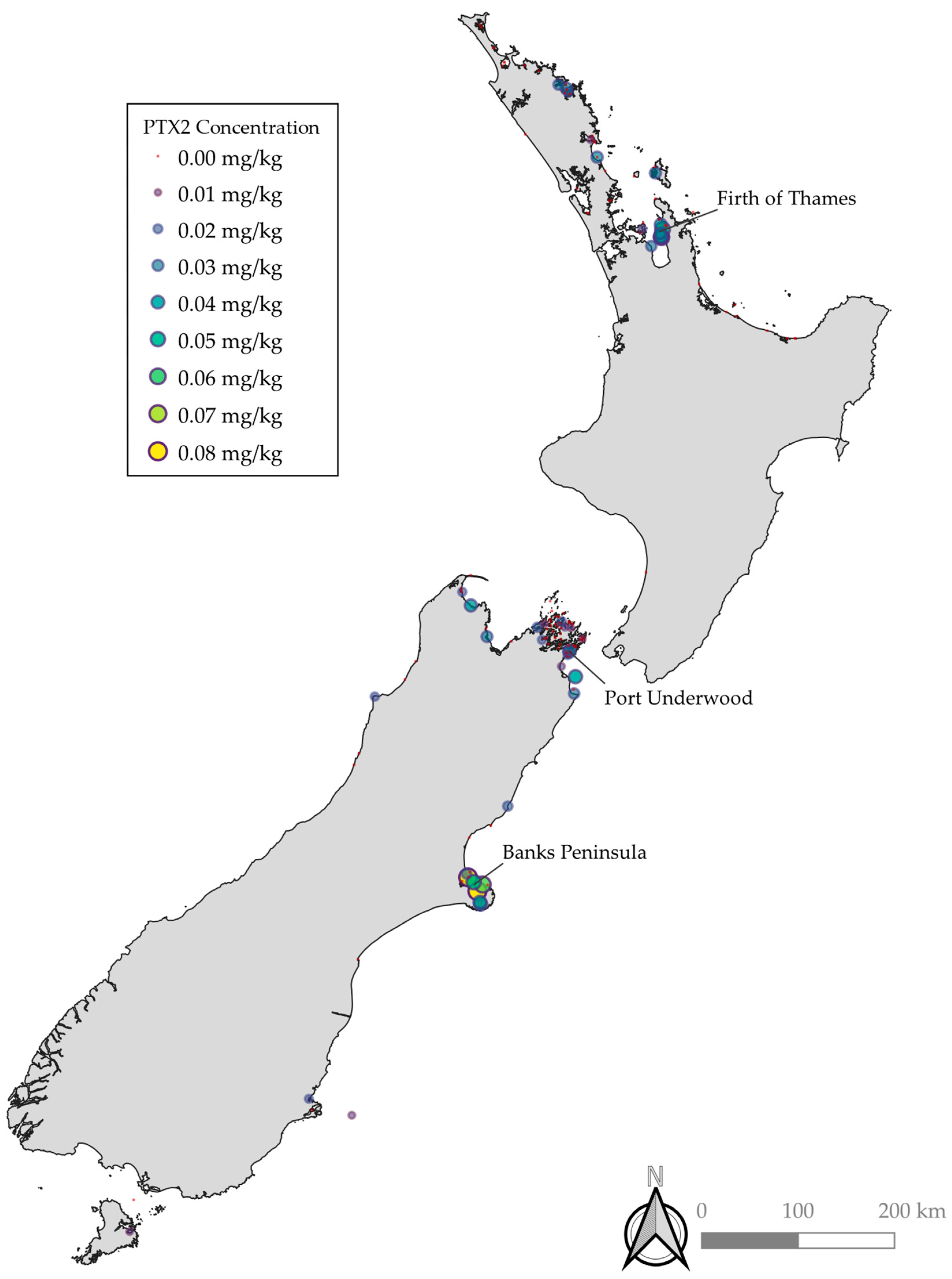
2.1. Distribution of PTXs in New Zealand
2.1.1. Spatial Distribution of PTXs
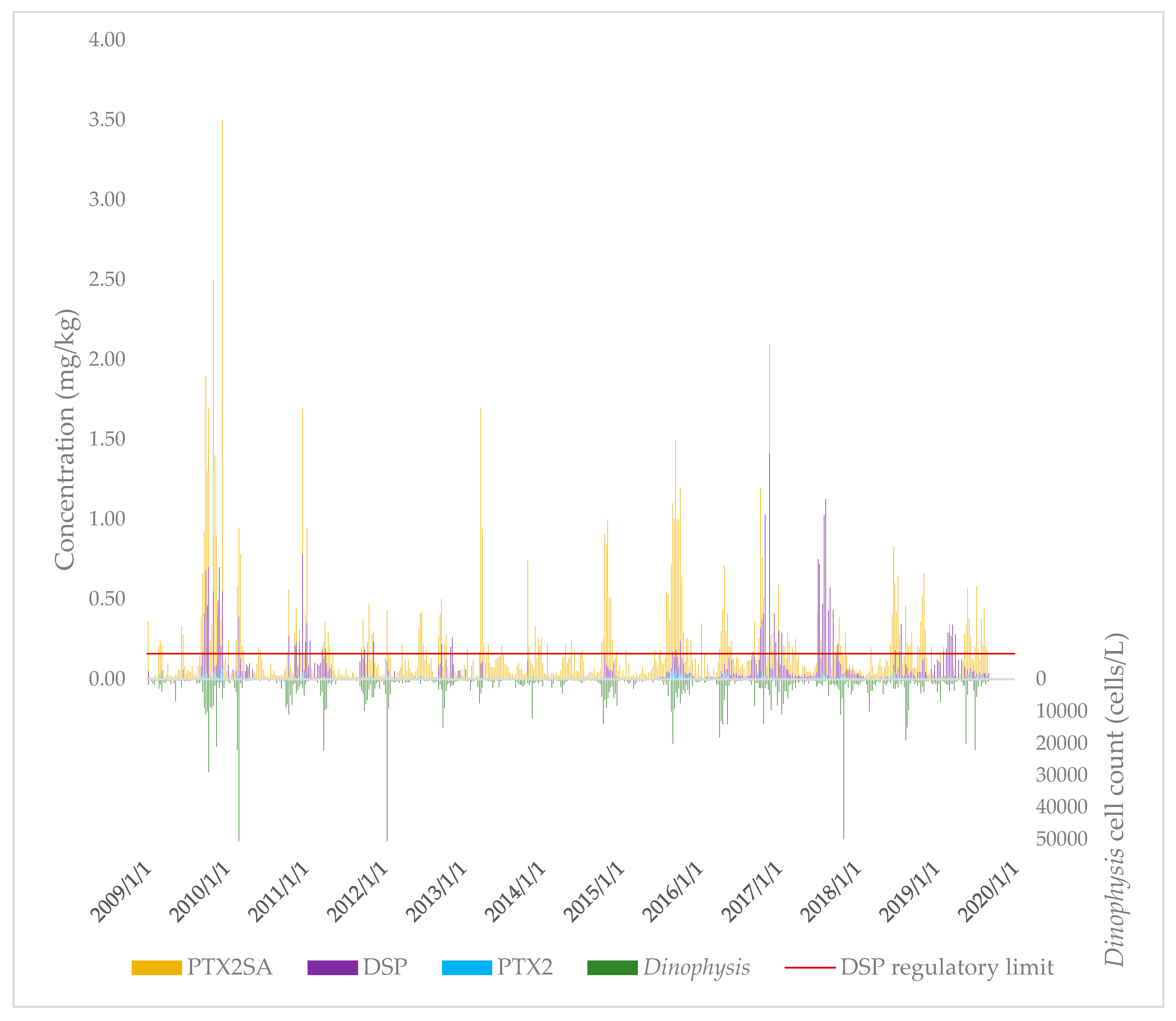
2.1.2. Temporal Distribution of PTXs
2.1.3. Species Distribution of PTX2
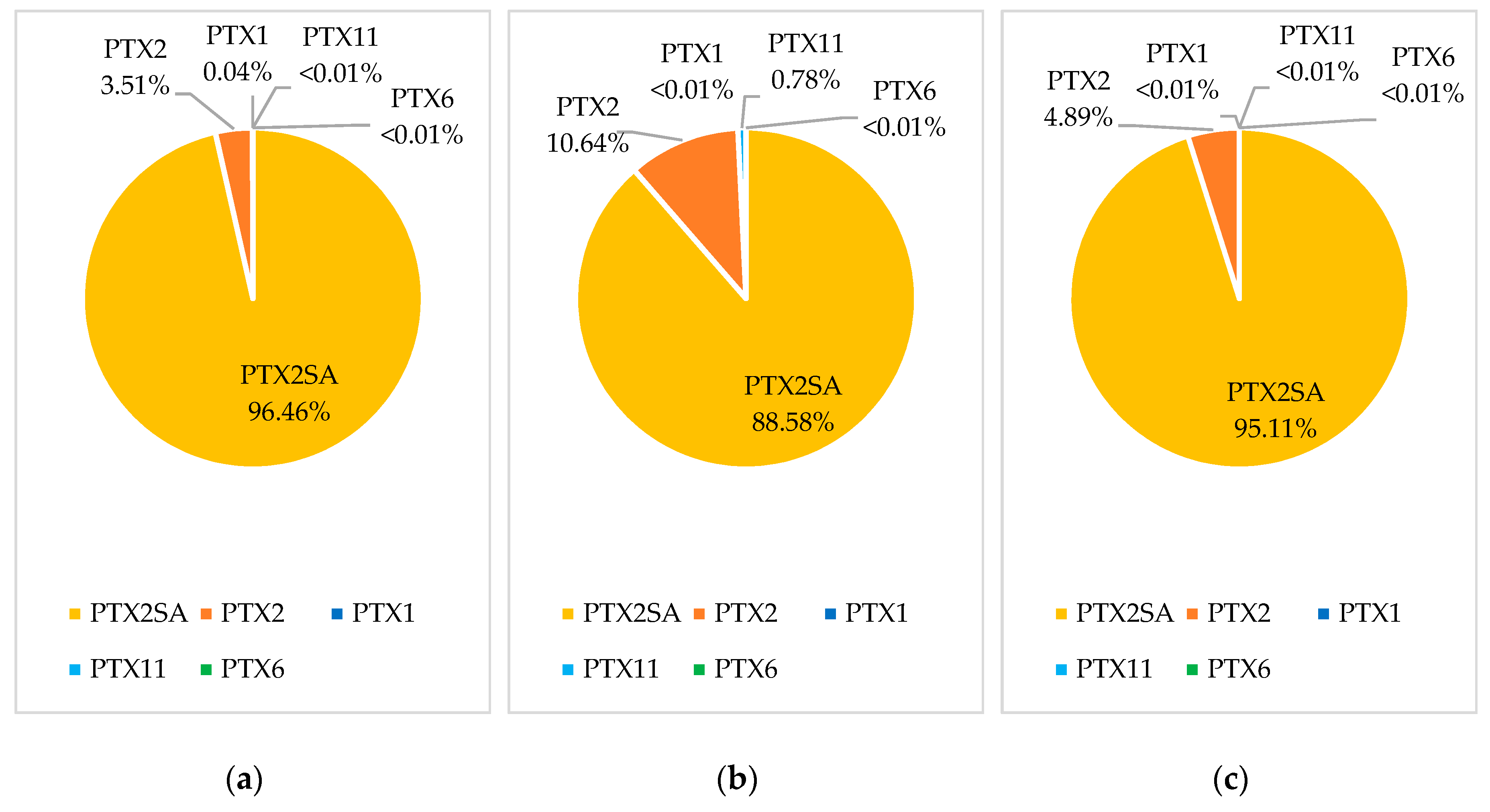
2.1.4. Pectenotoxin Profiles
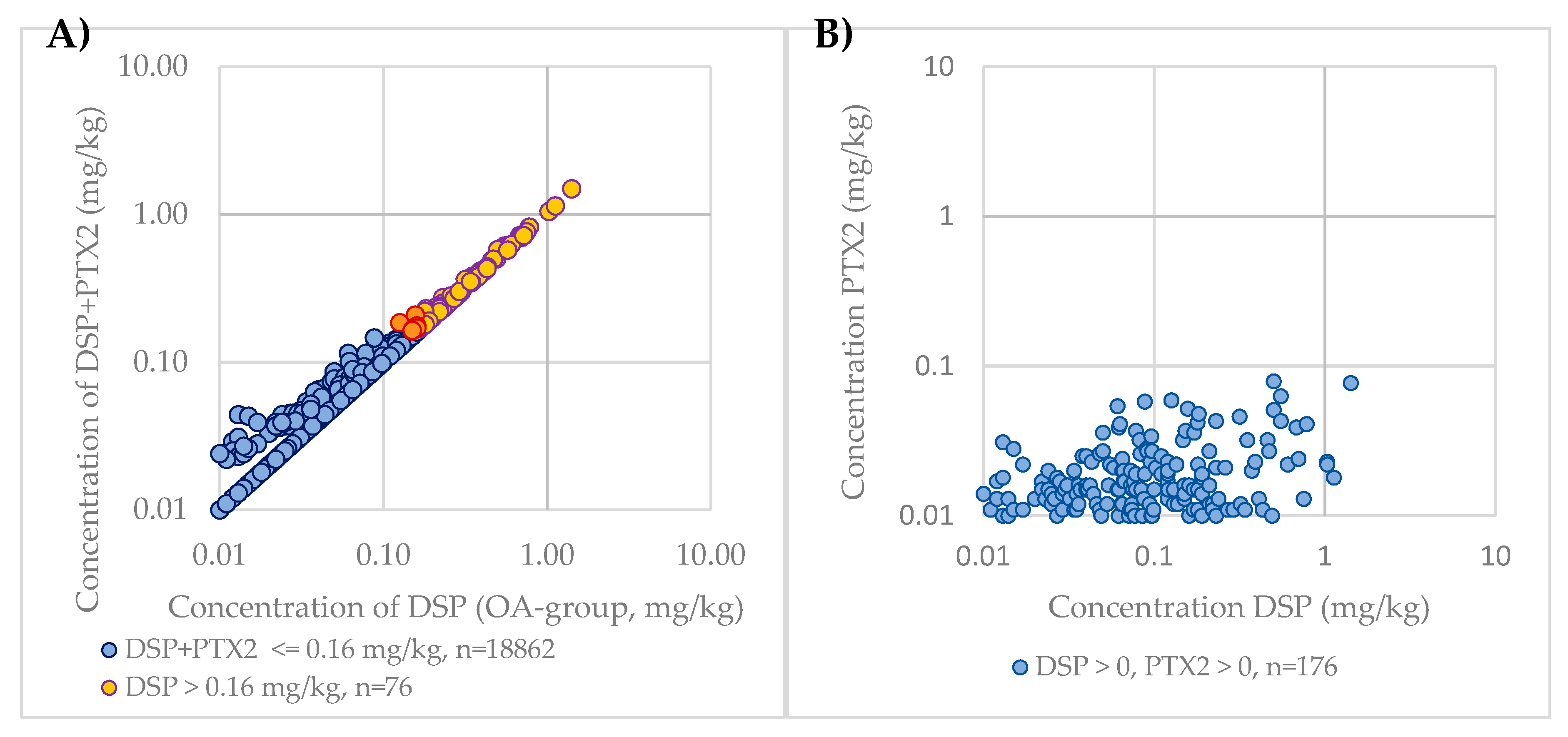
2.1.5. Impact of PTXs Contribution to DSP Levels
2.2. Risk Assessment
2.2.1. Deterministic Risk Assessment
2.2.2. Probabilistic Risk Assessment
3. Discussion
4. Materials and Methods
4.1. Exposure Data
- (a)
- PTX2 was at or above reportable levels (0.01 mg/kg).
- (b)
- DSP toxins were at or above reportable levels (0.05 mg/kg until June 2015, 0.01 mg/kg after June 2015).
- (c)
- PTX2SAs (sum of PTX2SA and 7-epi-PTX2SA) was at or above 0.1 mg/kg (10-fold higher than the reporting limit of 0.01 mg/kg).
4.2. Risk Assessment
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yasumoto, T.; Murata, M.; Oshima, Y.; Sano, M.; Matsumoto, G.; Clardy, J. Diarrhetic shellfish toxins. Tetrahedron 1985, 41, 1019–1025. [Google Scholar] [CrossRef]
- Yasumoto, T.; Oshima, Y.; Yamaguchi, M. Occurence of a new type of shellfish poisoning in the Tohoku district. Bull. Jpn. Soc. Sci. Fish. 1978, 44, 1249–1255. [Google Scholar] [CrossRef]
- Suzuki, T.; Beuzenberg, V.; MacKenzie, L.; Quilliam, M.A. Liquid chromatography–mass spectrometry of spiroketal stereoisomers of pectenotoxins and the analysis of novel pectenotoxin isomers in the toxic dinoflagellate Dinophysis acuta from New Zealand. J. Chromatogr. A 2003, 992, 141–150. [Google Scholar] [CrossRef]
- McNabb, P.; Selwood, A.I.; Holland, P.T.; Aasen, J.; Aune, T.; Eaglesham, G.; Hess, P.; Igarishi, M.; Quilliam, M.; Slattery, D.; et al. Multiresidue Method for Determination of Algal Toxins in Shellfish: Single-Laboratory Validation and Interlaboratory Study. J. Aoac Int. 2005, 88, 761–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- University of Otago and Ministry of Health. A Focus on Nutrition: Key Findings of the 2008/09 New Zealand Adult Nutrition Survey; Ministry of Health: Wellington, New Zealand, 2011; ISBN 978-0-478-37348-6. [Google Scholar]
- EFSA (The European Food Safety Authority). Scientific Opinion of the Panel on Contaminants in the Food Chain on a Request from the European Commission on Marine Biotoxins in Shellfish—Pectenotoxin Group. Efsa J. 2009, 1109, 1–47. [Google Scholar]
- Yasumoto, T. Polyether toxins produced by dinoflagellates. In Mycotoxins and Phycotoxins ’88; Natori, S., Hashimoto, K., Ueno, Y., Eds.; Elsevier: Amsterdam, The Netherlands, 1989; pp. 375–382. [Google Scholar]
- Yoon, Y.M.; Kim, Y.C. [Acute toxicity of pectenotoxin-2 and its effects on hepatic metabolising enzyme system in mice]. Korean J. Toxicol. 1997, 13, 183–186. (In Korean) [Google Scholar]
- Sasaki, K.; Wright, J.L.C.; Yasumoto, T. Identification and Characterization of Pectenotoxin (PTX) 4 and PTX7 as Spiroketal Stereoisomers of Two Previously Reported Pectenotoxins. J. Org. Chem. 1998, 63, 2475–2480. [Google Scholar] [CrossRef]
- Miles, C.O.; Wilkins, A.L.; Munday, R.; Dines, M.H.; Hawkes, A.D.; Briggs, L.R.; Sandvik, M.; Jensen, D.J.; Cooney, J.M.; Holland, P.T.; et al. Isolation of pectenotoxin-2 from Dinophysis acuta and its conversion to pectenotoxin-2 seco acid, and preliminary assessment of their acute toxicities. Toxicon 2004, 43, 1–9. [Google Scholar] [CrossRef]
- Miles, C.O.; Wilkins, A.L.; Munday, J.S.; Munday, R.; Hawkes, A.D.; Jensen, D.J.; Cooney, J.M.; Beuzenberg, V. Production of 7-epi-Pectenotoxin-2 Seco Acid and Assessment of Its Acute Toxicity to Mice. J. Agric. Food Chem. 2006, 54, 1530–1534. [Google Scholar] [CrossRef]
- Suzuki, T.; Walter, J.A.; LeBlanc, P.; MacKinnon, S.; Miles, C.O.; Wilkins, A.L.; Munday, R.; Beuzenberg, V.; MacKenzie, A.L.; Jensen, D.J.; et al. Identification of pectenotoxin-11 as 34S-hydroxypectenotoxin-2, a new pectenotoxin analogue in the toxic dinoflagellate Dinophysis acuta from New Zealand. Chem. Res. Toxicol.. 2006, 19, 310–318. [Google Scholar] [CrossRef]
- Ishige, M.; Satoh, N.; Yasumoto, T. Pathological studies on mice administered with the causative agent of diarrhetic shellfish poisoning (okadaic acid and pectenotoxin-2). Hokkaidoritsu Eisei Kenkyushoho 1988, 38, 15–18. [Google Scholar]
- Ogino, H.; Kumagai, M.; Yasumoto, T. Toxicologic evaluation of yessotoxin. Nat. Toxins. 1997, 5, 255–259. [Google Scholar] [CrossRef]
- Suzuki, T.; Mitsuya, T.; Matsubara, H.; Yamasaki, M. Determination of pectenotoxin-2 after solid-phase extraction from seawater and from the dinoflagellate Dinophysis fortii by liquid chromatography with electrospray mass spectrometry and ultraviolet detection. Evidence of oxidation of pectenotoxin-2 to pectenotoxin-6 in scallops. J. Chromatogr. A 1998, 815, 155–160. [Google Scholar] [PubMed]
- FAO/IOC/WHO. Report of the Joint FAO/IOC/WHO ad hoc Expert Consultation on Biotoxins in Bivalve Molluscs; Food and Agriculture Organization of the United Nations: Oslo, Norway, 2004; p. 31. [Google Scholar]
- Botana, L.M.; Hess, P.; Munday, R.; Nathalie, A.; DeGrasse, S.L.; Feeley, M.; Suzuki, T.; Berg, M.V.D.; Fattori, V.; Gamarro, E.G.; et al. Derivation of toxicity equivalency factors for marine biotoxins associated with Bivalve Molluscs. Trends Food Sci. Technol. 2017, 59, 15–24. [Google Scholar] [CrossRef] [Green Version]
- FAO; WHO. Technical paper on Toxicity Equivalency Factors for Marine Biotoxins Associated with Bivalve Molluscs; FAO: Rome, Italy, 2016; 108p, ISBN 978-92-5-109345-0. [Google Scholar]
- Fladmark, K.E.; Serres, M.H.; Larsen, N.L.; Yasumoto, T.; Aune, T.; Døskeland, S.O. Sensitive detection of apoptogenic toxins in suspension cultures of rat and salmon hepatocytes. Toxicon 1998, 36, 1101–1114. [Google Scholar] [CrossRef]
- FAO; WHO; IOC. Report of the Working Group Meeting to Assess the Advice from the Joint FAO/WHO/IOC Ad Hoc Expert Consultation on Biotoxins in Bivalve Molluscs; CX/FFP 06/28/6-Add.1; FAO: Beijing, China, 2006. [Google Scholar]
- EFSA (The European Food Safety Authority). Scientific Opinion of the Panel on Contaminants in the Food Chain on a request from the European Commission on Marine Biotoxins in Shellfish—Summary on regulated marine biotoxins. EFSA J. 2009, 130, 1–23. [Google Scholar]
- FAO; WHO. Report of the Thirty-Second Session of the Codex Committee on Fish and Fishery Products; FAO: Bali, Indonesia, 2013. [Google Scholar]
- Terao, K.; Ito, E.; Ohkusu, M.; Yasumoto, T. A comparative Study of the Effects of DSP-toxins on mice and rats. In Toxic Phytoplankton Blooms in the Sea: Proceedings of the Fifth International Conference on Toxic Marine Phytoplankton, Newport, RI, USA, 28 October–1 November 1991; Theodore, Y.S., Smayda, J., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 581–587. [Google Scholar]
- Hamano, Y.; Kinoshita, Y.; Yasumoto, T. Enteropathogenicity of Diarrhetic Shellfish Toxins in Intestinal Models. J. Food Hyg. Soc. Jpn. 1986, 27, 375–379. [Google Scholar] [CrossRef]
- Rao, G.N.; Peace, T.A.; Hoskins, D.E. Training could prevent deaths due to rodent gavage procedure. Contemp. Top. Lab. Anim. Sci. 2001, 40, 7–8. [Google Scholar]
- Damsch, S.; Eichenbaum, G.; Tonelli, A.; Lammens, L.; van den Bulck, K.; Feyen, B.; Vandenberghe, J.; Megens, A.; Knight, E.; Kelley, M. Gavage-related reflux in rats: Identification, pathogenesis, and toxicological implications. Toxicol. Pathol. 2011, 39, 348–360. [Google Scholar] [CrossRef] [Green Version]
- Munday, R. Toxicology of Seafood Toxins: A Critical Review. In Seafood and Freshwater Toxins: Pharmacology, Physiology, and Detection; Botana, L.M., Ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 197–290. [Google Scholar]
- Burgess, V.A. Toxicology Investigations with the Pectenotoxin-2 Seco Acids; Griffith University: Queensland, Australia, 2003. [Google Scholar]
- Munday, R. Toxicology of Seafood Toxins: Animal Studies and Mechanisms of Action. Recent Adv. Anal. Mar. Toxins. 2017, 78, 211. [Google Scholar]
- FAO; WHO. Report of the Thirty-Third Session of the Codex Committee on Fish and Fishery Products; FAO: Bergen, Norway, 2014. [Google Scholar]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2019; Available online: https://www.R-project.org (accessed on 31 October 2019).
- Pouillot, R.; Delignette-Muller, M.-L. Evaluating variability and uncertainty separately in microbial quantitative risk assessment using two R packages. Int. J. Food Microbiol. 2010, 142, 330–340. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | No. Samples | Detections | % Detected | Min | Max | Mean | Median | 97.5 PCTL |
|---|---|---|---|---|---|---|---|---|
| 2009 | 1688 | 56 | 3.3% | 0.010 | 0.063 | 0.019 | 0.015 | 0.048 |
| 2010 | 1618 | 14 | 0.9% | 0.010 | 0.041 | 0.014 | 0.011 | 0.035 |
| 2011 | 1684 | 21 | 1.2% | 0.010 | 0.043 | 0.016 | 0.014 | 0.038 |
| 2012 | 1647 | 13 | 0.8% | 0.011 | 0.025 | 0.015 | 0.013 | 0.024 |
| 2013 | 1723 | 5 | 0.3% | 0.010 | 0.021 | 0.017 | 0.019 | 0.021 |
| 2014 | 1776 | 10 | 0.6% | 0.010 | 0.016 | 0.013 | 0.014 | 0.016 |
| 2015 | 1871 | 66 | 3.5% | 0.010 | 0.059 | 0.021 | 0.017 | 0.053 |
| 2016 | 1836 | 21 | 1.1% | 0.010 | 0.079 | 0.026 | 0.021 | 0.078 |
| 2017 | 1924 | 14 | 0.7% | 0.010 | 0.027 | 0.017 | 0.017 | 0.026 |
| 2018 | 1857 | 12 | 0.6% | 0.011 | 0.058 | 0.023 | 0.017 | 0.054 |
| 2019 | 1323 | 19 | 1.4% | 0.010 | 0.024 | 0.014 | 0.012 | 0.023 |
| Total | 18947 | 251 | 1.3% | 0.010 | 0.079 | 0.019 | 0.015 | 0.052 |
| Month | No. Samples | Detections | % Detected | Min | Max | Mean | Median | 97.5 PCTL |
|---|---|---|---|---|---|---|---|---|
| January | 1615 | 10 | 0.6% | 0.011 | 0.043 | 0.020 | 0.016 | 0.041 |
| February | 1617 | 30 | 1.9% | 0.010 | 0.027 | 0.014 | 0.012 | 0.026 |
| March | 1679 | 10 | 0.6% | 0.010 | 0.023 | 0.013 | 0.011 | 0.021 |
| April | 1594 | 11 | 0.7% | 0.010 | 0.039 | 0.016 | 0.014 | 0.035 |
| May | 1634 | 10 | 0.6% | 0.011 | 0.022 | 0.015 | 0.015 | 0.022 |
| June | 1574 | 15 | 1.0% | 0.010 | 0.058 | 0.020 | 0.016 | 0.052 |
| July | 1594 | 9 | 0.6% | 0.011 | 0.027 | 0.016 | 0.015 | 0.026 |
| August | 1563 | 21 | 1.3% | 0.010 | 0.052 | 0.022 | 0.018 | 0.047 |
| September | 1514 | 47 | 3.1% | 0.010 | 0.059 | 0.021 | 0.018 | 0.054 |
| October | 1593 | 50 | 3.1% | 0.010 | 0.046 | 0.017 | 0.015 | 0.034 |
| November | 1542 | 28 | 1.8% | 0.010 | 0.079 | 0.024 | 0.017 | 0.078 |
| December | 1428 | 10 | 0.7% | 0.010 | 0.063 | 0.021 | 0.013 | 0.058 |
| Total | 18947 | 251 | 1.3% | 0.010 | 0.079 | 0.019 | 0.015 | 0.052 |
| Organism 1 | Sites | No. Samples | Detections | % Detected | Min | Max | Mean | Median | 97.5 PCTL |
|---|---|---|---|---|---|---|---|---|---|
| Green-lipped mussel | 83 | 15947 | 186 | 1.2% | 0.010 | 0.079 | 0.019 | 0.015 | 0.056 |
| Pacific oyster | 22 | 1141 | 40 | 3.5% | 0.010 | 0.027 | 0.015 | 0.015 | 0.026 |
| Clam | 11 | 1042 | 6 | 0.6% | 0.013 | 0.027 | 0.018 | 0.016 | 0.026 |
| Scallop | 20 | 298 | 4 | 1.3% | 0.012 | 0.032 | 0.020 | 0.017 | 0.031 |
| Dredge oyster | 8 | 228 | 1 | 0.4% | 0.043 | 0.043 | 0.043 | 0.043 | 0.043 |
| Surf clam | 6 | 97 | 5 | 5.2% | 0.010 | 0.024 | 0.015 | 0.012 | 0.023 |
| Blue mussel | 12 | 56 | 7 | 12.5% | 0.011 | 0.042 | 0.021 | 0.020 | 0.039 |
| Queen scallop | 2 | 52 | 2 | 3.8% | 0.010 | 0.011 | 0.011 | 0.011 | 0.011 |
| Tuatua | 5 | 28 | 0 | ||||||
| Pipi | 2 | 19 | 0 | ||||||
| Cockle | 3 | 17 | 0 | ||||||
| Oyster | 5 | 9 | 0 | ||||||
| Abalone | 3 | 8 | 0 | ||||||
| Geoduck | 3 | 5 | 0 | ||||||
| Total | 144 | 18947 | 251 | 1.3% | 0.010 | 0.079 | 0.019 | 0.015 | 0.052 |
| Parameter | Units | 97.5 Percentile | Maximum |
|---|---|---|---|
| Concentration PTX2 | mg PTX2/kg | 0.01 | 0.079 |
| Exposure by eating 100 g | µg PTX2/person | 1.0 | 7.9 |
| µg PTX2/kg b.w. | 0.02 | 0.13 | |
| Exposure by eating 268 g | µg PTX2/person | 2.7 | 21.2 |
| µg PTX2/kg b.w. | 0.04 | 0.35 | |
| Exposure by eating 400 g | µg PTX2/person | 4.0 | 31.6 |
| µg PTX2/kg b.w. | 0.07 | 0.53 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boundy, M.J.; Harwood, D.T.; Kiermeier, A.; McLeod, C.; Nicolas, J.; Finch, S. Risk Assessment of Pectenotoxins in New Zealand Bivalve Molluscan Shellfish, 2009–2019. Toxins 2020, 12, 776. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12120776
Boundy MJ, Harwood DT, Kiermeier A, McLeod C, Nicolas J, Finch S. Risk Assessment of Pectenotoxins in New Zealand Bivalve Molluscan Shellfish, 2009–2019. Toxins. 2020; 12(12):776. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12120776
Chicago/Turabian StyleBoundy, Michael J., D Tim Harwood, Andreas Kiermeier, Cath McLeod, Jeane Nicolas, and Sarah Finch. 2020. "Risk Assessment of Pectenotoxins in New Zealand Bivalve Molluscan Shellfish, 2009–2019" Toxins 12, no. 12: 776. https://0-doi-org.brum.beds.ac.uk/10.3390/toxins12120776